univer?!ty of bath
TRANSCRIPT

University of Bath
PHD
Sulphite preservation of British fresh sausage.
Banks, Jeffrey Gordon
Award date:1983
Awarding institution:University of Bath
Link to publication
Alternative formatsIf you require this document in an alternative format, please contact:[email protected]
Copyright of this thesis rests with the author. Access is subject to the above licence, if given. If no licence is specified above,original content in this thesis is licensed under the terms of the Creative Commons Attribution-NonCommercial 4.0International (CC BY-NC-ND 4.0) Licence (https://creativecommons.org/licenses/by-nc-nd/4.0/). Any third-party copyrightmaterial present remains the property of its respective owner(s) and is licensed under its existing terms.
Take down policyIf you consider content within Bath's Research Portal to be in breach of UK law, please contact: [email protected] with the details.Your claim will be investigated and, where appropriate, the item will be removed from public view as soon as possible.
Download date: 19. Feb. 2022

UNIVER?!TY OF BATH


SULPHITE PRESERVATION OF BRITISH FRESH
SAUSAGE
Submitted by Jeffrey Gordon Banks
for the degree of Ph.D.
of the University of Bath
1983
COPYRIGHT
Attention is drawn to the fact that copyright of this thesis rests
with its author. This copy of the thesis has been supplied on
condition that anyone who consults it is understood to recognise
that its copyright rests with its author and that no quotation
from the thesis and no information derived from it may be published
without the prior written consent of the author.
This thesis may be made available for consultation within the
University Library and may be photocopied or lent to other
libraries for the purposes of consultation.

ProQuest Number: U344211
All rights reserved
INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest U344211
Published by ProQuest LLC(2015). Copyright of the Dissertation is held by the Author.
All rights reserved.This work is protected against unauthorized copying under Title 17, United States Code.
Microform Edition © ProQuest LLC.
ProQuest LLC 789 East Eisenhower Parkway
P.O. Box 1346 Ann Arbor, Ml 48106-1346

11
Contents
PageTitle page
Acknowledgements
Summary
Introduction 1
Literature Review; 4
Sausage manufacture 4
Microbiology 6
Sulphite preservation 39
(+ Figure 1)
Materials and Methods; 60
Sausage manufacture - storage and sampling 60
Sulphite determination 62
Sulphite-binding agents determination 67
Sulphate determination 68
pH determination 69
Isolation, enumeration, identification and
maintenance of micro-organisms 70
Pure culture studies 83
(+ Figures 2-9)
Results ; 95
Chemical results 95
Microbiology of sausage 125
Pure culture studies 147
Salmonella-seeding of sausage 157
(+ Figures 10-70)

Ill
Page
Salmonella Contamination of Sausagre 222
Introduction 222
Literature review 235
Materials and Methods 238
Results 250
Discussion 258
(+ Figures 71-72)
Pseudomonas
Introduction and historical perspective 258
Early work with meat isolates 259
Sources of Pseudomonas 252
Classification of Pseudomonas frcm sausage 264
(+ Figures 73-77)
Discussion 294
(+ Figures 85-90) 323
Commercial Implications (+ Figures 78-84) 308— '— — J f — IT—t— »->—.-r--Appendix 329
References 335

IV,
ACKNOWLEDGEMENTS
I would like to thank the following:
The Science and Engineering Research Council for the research
grant.
The St. Ivel Technical Centre, Bradford-on-Avon, for the C.A.S.E.
award, financial assistance and access to their factories,
laboratories and library.
The School of Biological Sciences at the University of Bath and
Professor A.H. Rose for provision of various facilities.
Ron Board for introducing me to the fascinating world of
"bangers" and for his encouragement, advice and criticism
throughout this project.
My fajmily for their support, and my wife, Karen, for her patience,
Mrs. Judy Harbutt for skilfully typing this thesis.

V.
SUMMARY
The microbial associations that developed in commercially
produced sulphited and unsulphited sausages during storage for
up to 8 days at a variety of températures were identified. Such
associations were comprised of dominant, e.g. Brochothrix
thermosphacta, yeasts and lactic acid bacteria, or minor, e.g.
Pseudomonadaceae and Enterobacteriaiceae, groups. A detailed
analysis of numerous isolates of thie previously-ignored Gram-
negative microflora using computer-assisted numerical techniques
revealed that sulphite concentration influenced the composition
of the Enterobacteriaceae but not the pseudomonads.
A method was developed for the assay of free and bound
sulphur IV oxospecies in culture mecdia or meat-based samples.
It was superior to existing techniques and demonstrated limited,
irretrievable - by oxidation - and extensive reversible - by
binding - loss of sulphite preservative frcm freshly manufactured
and stored sausage.
Selected micro-organisms isolated from sausage were tested
for their tolerance of free and bound sulphur IV oxospecies
in batch and "turbidometer" culture under various conditions.
In general this technique reflected the elective/selective
role of sulphite in sausage and allowed the microbial associ
ation to be defined more clearly in terms of tolerance of the
active preservative.

VI.
The incidence and level of contamination of sausage and
its ingredients with Salmonella was established
using a 'most-probable number' method. Despite a high
incidence of contamination of most of the meat ingredients and
finished product with these organisms, the level of infection ■
with the exception of mechanically recovered meat - was low.
Thus the role of sulphur IV oxospecies in determining the
fate of these food poisoning organisms in sausage or culture
media was assessed by deliberate infection with rifampicin-
resistant Salmonella mutants.
With a view to extending the shelf life of fresh sausage,
a limited survey of adjuncts or alternative "preservatives"
to sulphite using pilot-scale batches of sausage meat was
done.

INTRODUCTION
As British fresh sausages are made from diced or minced meat,
fat and rind from various sources, there are ample opportunities
for contamination with both food spoilage and food poisoning
micro-organisms. Extensive comminution of the ingredients in
a bowl chopper ensures not only that the contaminants are distributed
randomly, but that they are suspended in an environment notable
for its chemical activity as well as its potential to support
extensive microbial growth. Thus three amylases - one present
in rusk, another in porcine muscle and a third in the salivary
gland - and a maltase of meat origin ensure that substrate
levels of glucose, maltose, maltptriose and maltotetrose are
available to contaminants of pork sausages stored at 4° or 22°C
(Abbiss, 1978). The accumulation of valerate and changes in the
lipid pattern, as indexed by g.l.c. analysis of stored sausages,
have led also to the tentative conclusio n that enzyme-mediated
changes of fats and proteins occur in sausages (Leads, 1979).
As yet, however, the relative importance of enzymes of microbial
and meat origin have not been established. There is evidence
also that a small amount of the legally-permitted preservative
of sausages, sulphite or metabisulphite, is irretrievably lost
in stored sausages and a large amount is reversibly bound such
that its antimicrobial properties are negated (Brown, 1977).
Determinations of pH changes provide yet another index of
chemical changes, there being an acid drift of ca. 1 pH in
sulphited and ca. 2.8 units in unsulphited sausage (Brown, 1977).
Although sulphite is regarded as a preservative, the physiological
basis of its mode of action is unknown and the studies of the

microbiology of sausages permit only a general conclusion, namely
that it selects a Gram-positive flora (Dyett and Shelley, 1962,
1966; Dowdell and Board, 1967, 1968, 1971). Brown (1977) was
probably the first to note that Pseudomonas spp. grew in sulphited
sausages and he demonstrated, also, that enterobacteria flourished
in unsulphited ones stored at 22°c. Thus he supported indirectly
the notion (Dyett and Shelley, 1962) that sulphite plays an
important role in preventing the growth of Salmonella in sausages.
As these studies (Dowdell and Board, 1968, 1971) which defined
the microbial association - lactic acid bacteria, yeasts, micro
cocci and Microbacterium thermosphactum - of stored sausages
did not analyse the concentration of free sulphite in the products
examined, the general definition of this association may well
mask an important feature, namely that its members differ
markedly in their tolerance of free sulphite. A chemical method
was developed in the present study so that the concentration of
free and bound sulphite in sausages examined microbiologically
could be established. Many of the isolates from these experi
ments were identified by computer analysis and their tolerance
of free sulphite at different incubation temperatures established.
A novel test system was developed to establish the sulphite
tolerance of pure cultures at different incubation temperatures.
These studies, which are discussed on pp.147 -157 , have resulted
in a better definition of the microbial association found in
British fresh sausage.
As noted above, Dyett and Shelley (1962, 1966) surmised that
sulphite was responsible for the good public health record of

British fresh sausage in spite of the high incidence of contamination
with Salmonella (Roberts et al., 1975). Although large numbers
of samples were included in the surveys of Roberts et al. (1975)
and those of Turnbull and Rose (1982) and Barrell (1982), no
attempts were made to establish the level of contamination or to
identify the ingredients responsible for such infection. In the
present study, quantitative methods were used to establish the
level of Salmonella contamination of British fresh sausages and
ingredients taken in the course of routine production from a
factory. This phase of the study was extended to define the
behaviour of Salmonella in stored sulphited or unsulphited
sausage. Previous studies of the microbiology of sausages have
shown that batches made in experimental kitchens rarely contain
a microflora comparable to that produced in a factory. It
needs to be stressed therefore that the major work discussed
in this thesis was done with material taken from a factory.

LITERATURE REVIEW
Manufacture of pork and pork and beef sausage
Recipes for the pork sausages and the pork and beef sausages
included in this study are given in the Appendix (p.329 ).
As the method of manufacture differs slightly between firms, a
brief account of that used in this study will be given.
Large (ca. 50 x 20 x 20 cm) blocks of lean pork, and/or beef
and pork backfat, minced semi-lean pork (head and belly meat) and
cooked rinds were added to a bowl chopper (Laska, Austria) -
capacity, 500 I. The mincing of meat is associated with a
rise in the glucose concentration due to the action of intrinsic
a-amylases, maltases and amylo-l-6-glycosidases (Newbold and
Scopes, 1971) and accounts in part for the increased chemical
activity of the sausage meat. Seasonings, ice, water and poly
phosphates were added to the meat and a slurry produced by rapid
chopping. The seasonings contained: (a) spices, some of which
may act as bacteriostatic/bacteriocidal agents (Chipault et al.,
1952) or antioxidants (Chipault et al., 1952, 1956); (b) a
pigment ("Red 20"); (c) preservative, (sodium sulphite or
metabisulphite) to give a final concentration just above the
legally-permitted maximum of ca, 600 yg SO^/g freshly-made
sausage, and (d) sodium chloride. Rusk was the last ingredient
to be chopped into the meat slurry, its principal functions
being to retain water (Haq et al., 1973) thereby ensuring
successful extrusion of the slurry into casings and to contribute
to flavour (Leads, 1979) . Of the two types of casing in
common use, reconstituted collagen or synthetic cellulose.

5 .
the former was used.
Probable changes during preparation
Little work has been done on physical changes occurring
during the manufacture of British fresh sausage. This section
relies therefore on observations made in studies of related products,
frankfurters, Thuringer and Bologna-type products.
"Bowl chopping" destroys the structure of the fat and meat.
The cell contents dissolve in the iced water and it is surmised
that very rapid comminution coats fat particles with a protein
film (Borchert et al., 1967), The presence of ice during
processing probably favours the formation of a pseudo-emulsion
simply because the solubility of protein in dilute brine is in
creased by chill temperatures (Karmas, 1977). Moreover, the
addition of sodium chloride at an early stage of the preparation
of a slurry has been shown to optimize the extraction of the
salt-soluble proteins, actomysin and myosin, and provide an
ionic environment (Karmas, 1977) which increases the rate of
formation of the "pseudo-emulsion" (Theno et al., 1978).
The addition of polyphosphate salts - "Calgon" (sodium hexa-
metaphosphate) or fibrisol VIO - to the mix in the early
stages of bowl chopping, aid the retention of water by the
meat fibres and protein. This is due in part to the stabilization
of the pH at or near neutrality; as meat approaches its iso
electric point (pH 5.5) water and water-soluble materials are
released from the meat matrix (Lawrie, 1978). The number of
fat pockets are reduced also, thus favouring even "émulsification"

(Gerrard, 1976). The protein film is considered to be a major
barrier to the coalescence of small fat globules formed by
vigorous chopping (Acton and Saffle, 1972). It could be in
ferred from this discussion that British fresh sausage was basically
a fat-in-water emulsion. Indeed this term is commonly used in the
trade, even though the commodity rarely if ever has either the
structure or the properties of a conventional oil-in-water
emulsion.
Microbiology*
Sources of micro-organisms
As the initial size and composition of the microflora present
in freshly made sausage can influence the development of the
microbial association during storage (Dowdell and Board, 1971;
Brown, 1977), it is pertinent to consider factors that contribute
to the contamination of the meat ingredients, the principal
depots of infection (Dowdell and Board, 1968, 1971). There are
two major sources of the micro-organisms on meat, namely those of
gut or mucous-membrane origin that infect the meat during
slaughter and those acquired through contact of meat with equipment
etc. Ingram (1949) was perhaps the first to distinguish between
these two sources of infection when he alluded to organisms of
* The names of micro-organisms used in this thesis are those
given in the cited reports.

"intrinsic" and "extrinsic" origin respectively. He stated that
the intrinsic micro-organisms in the tissues of healthy animals
need not be pathogenic; the latter would be expected to occur
in the carcasses of animals which had been suffering from overt
or covert infections. The high temperature storage of carcass
meat has been considered (Ingram and Dainty, 1971) to be the main
reason for growth of intrinsic bacteria, such as Clostridium spp. ,
which cause "bone taint" for example. The occurrence of organisms
in meat suggests that some may have penetrated the wall of the
gut or mucous membranes and been disseminated via the lymphatic
and vascular systems (Arnold, 1928; Nickel and Gisske, 1941;
Bogadi and Sewell, 1974; Gill et al., 1976). Although many
workers have reported the isolation of micro-organisms from within
meat (e.g. Ayres, 1955; Lechowich, 1971; Lawrie, 1974)
a critical review of the literature
(Gill, 1979) drew attention to technical difficulties, particularly
with respect to asepsis, that need to be controlled before valid
conclusions can be made. Gill (1979) cites the observations
of Haines and Scott (1940) and Sharp (1963) who showed that
flaming of dissection instruments was at best only 70% successful
in achieving sterilisation. It is evident from the literature
(Table 1), that a greater incidence and level of contamination
of deep muscle tissue was found by workers before 1970, whereas
the consensus of opinion after this date favours the view that
such tissues in healthy animals are usually sterile (e.g. Hasegawa
et ai., 1970; Buckley et ai., 1976 and Gill et ai., 1978).
Indeed, Roberts (1980) concluded that the presence of small

Icn3PQ)uSx:c3u-i0Q)3050)•H-P
Ü0)E»oTJM-i0053ffl4J05U•PS'rH0•HO4JA0)x:&H
r-405
I
iwO4-5•H4Jg
05
P 05 0505 y (0O 4J c0 y 0 0p ço 5 U-l
A 0 yy 'O c> 3 >14- 0 y pP 05 0U x; Ai •P 04 y05 y Û4 euE 0 05 eu05 •P P 05 y 05 •p•P U U 3 P uC y s y y itj y e uto 0 3 0 y •p •u 3 0tr y •p y 0 x: y •P yp 0 0 y y y 00 1-4 •P rp 0 •p X) •P 1-41 >1 P >1 4J P 0 P >10 X 4J X: 0. y P -P X:p A 05 & a y y W Ouy (0 0 Oi (0 P y ■u 0 to•p 4J rp 05 4J •p 05 3 M 4-5E W V w V5 w kl O CO
051•Ha(0yI0V•rH2
O'
055(tJ -P'P c/fi 05 O ÛU
sgg0-1s
= T) 05 0) P -P 05 P
8
&P4-1§
05Cr>•H
CNOrsi M O
OM r~ TT
1Ip§
VD
mr-4 1/50505 rprp05 Mw yw 'q 05y e r-4 44 uoK (d y vp05 05CM V 4-5 c >1 rp05 C C 0 Adr4 y td w WN r4 44 Cc P y y tdX: y y Ad > >144 05 T3 y Q fd•p c c •p 04 Py y fd z y y« *3 > XI z
r-CM
05«
4-5-P(0u
CMo o
S'I
05•S'a,
&I-p'CJ•HP4J05O
1/5 O
M o00 y r~ r»1/5 05 0505 44 1—I 00r4 y 05 m r -
VD f '- 05w V 05 M 05 rp
o rp y rp MM y yy 44
X 44 C y •P 444J E-t c 0 •P y yy 1 y (fi y Ad0 N r-4 3 y >1 44p U 05 p y y > y yy 3 U5 y Ad O' y r-4
'O Q 05 y y y Ad rpc 'O r-4 c •p y O' y rpy y y z y y 3 •PN c« > m N ü
05rp■P4J5»Q)5
g>■H4Jêg

numbers of viable micro-organisms in carcasses of healthy animals
was of very limited commercial importance, and he suggested that
the process of slaughter introduced micro-organisms into tissues.
Thus in an investigation of bone marrow of hogs, Jensen and Hess
(1941) isolated pigmented bacteria which had been used to
contaminate the "sticking knife". Similar results were obtained
when genetically-marked micro-organisms were used in this type
of experiment (Mackey and Derrick, 1979). Moreover, the surmise
(Wilson and Miles, 1964) that bacteria may reach the tissues by
penetration of the intestinal epithelium was supported by the
demonstration (Mackey and Derrick, 1979) that organisms admini
stered orally to pigs were isolated from the lungs and spleen
post-mortem. Although there is evidence of translocation (e.g.
Arnold, 1928; Nickel and Qisske, 1941), Roberts (1980) was of
the opinion that it makes a negligible contribution to the
contamination of market meat.
Extrinsic micro-organigns
Micro-organisms of the general environment are the major
contaminants of carcass meat. They infect initially cut surfaces
but may penetrate into meat during storage (Gill and Penney, 1977).
The following review deals with the two important aspects of
contamination in factories in which slaughter, butchery and
manufacture are done (Fig. 1), firstly the sources of contamination,
and secondly the factors influencing the level of contamination.

10.
Sources of contamination
The skin of the animal, dust, soil, water and air in lairages
and in abattoirs can all harbour substantial numbers of micro
organisms (Empey and Scott, 1939; Stolle, 1981) and climatic
conditions, particularly rainfall and temperature, may affect
the size and composition of the populations (Newton et al.,1978).
Scalding, singeing and "black scraping" are processes that
could be expected to have an important influence on the level of
contamination of a carcass. Scalding may cause a reduction of
0.9 - 2.5 log cycles in the total viable (Dockerty et ai., 1970;
Snijders, 1976; Snijders and Gerats, 1976) and of 3 log cycles in
the Enterobacteriaceae count (Gerats et ai., 1981). High temper
atures and alkaline water favour the reduction in bacterial
numbers (Dockerty et ai., 1970). The attachment of micro
organisms to porcine skin appears to offer protection (Butler
et ai., 1979). Thus thç numbers of Lactobacillus and Pseudomonas
putrefaciens inoculated intentionally ontp pigs' skin were reduced
by upwards of 3 - 4 log cycles during scalding (Butler et ai.,
1980). Singeing kills only those micro-organisms on porcine
skin with which the flame makes direct contact. Indeed some
workers, (e.g. Dockerty et ai., 1970 ; Butler et ai., 1980)
report small reductions only in the number of micro-organisms
during this process whereas others (e.g. Snijders, 1976;
Snijders and Gerats, 1976; Rasch et ai., 1978) infer that this
process is much more effective. Thus the former group of
workers noted a 0.4 - 1 and the latter a 2.5 - 3 log cycle
reduction in the total viable count. Scraping by hand does not

11
reduce the number of micro-organisms on the carcass (Dockerty et
al., 1970); indeed, mechanical scraping may even contribute
to contamination of the carcass (Gerats et al., 1981).
Throughout all these processes, the stick wound is the
principal avenue for microbial penetration of deep tissue;
subsequently the cut along the mid-line of the belly - the
initial step in evisceration - results in contamination of
internal tissues and surfaces. Micro-organisms derived from
workers, equipment or the animals' skin together with excessive
handling of the carcass results in an increase in the number of
contaminants (Dockerty et al., 1970). Gardner (1980) and
Roberts et al. (1980) demonstrated that the standards of
hygiene at the evisceration, stage determined the ultimate level
of contamination of the carcass. Rupture of the intestines
influences the size of the microbial load and the probability
of contamination of the carcass with food poisoning organisms
of gut origin (Gerats et al., 1981). The low temperature and
high humidity of the chill rooms favours the colonization of
the carcass with Gram-negative, aerobic psychrotrophic micro
organisms (Gill, 1980) and subsequent hand butchering, cleaning
of bones by machine to produce mechanically recovered meat
(MRM), and mincing are processes which have the potential to
increase further the level of contamination of meat. Thus there
is a discord in the literature dealing with the microbiological
status of deep tissues of freshly slaughtered animals, reports
that micro-organisms can be isolated readily (e.g. Narayan, 1966;
Narayan and Takacs, 1966; Pusztai, 1970), contrasting with

12
those which state that large amounts of sterile muscle can be
removed from carcasses (e.g. Radouco-Thomas et al., 1959;
Gardner and Carson, 1967; and Ockerman et ai., 1969). The
observations of the latter list of workers is supported by
Jensen and Hess (1941) who failed to isolate micro-organisms
in a biopsy of porcine muscle and yet noted an incidence of
contamination of 66% in samples taken immediately post-mortem.
Microbial invasion of the tissues
Micro-organisms lacking the determinants of virulence will
presumably fail to combat the antimicrobial systems should they
invade an unstressed host. This presumably accounts for the
germ-free state of the tissue fluids of healthy animals (Gill,
1979). With debilitated animals, however, opportunist micro
organisms may be the cause of short-lived, persistent or
recurrent infections (von Graevenitz, 1977). From his critical
review of the literature Gill (1979) concluded that ante-mortem
invasion leads to "intrinsic contamination".
The microbial associations of fresh sausage
Many of the early reports on sausage were concerned with
the contamination of the product with micro-organisms of public
health significance, the so-called "sanitary quality" of sausage
(Cary, 1916). Indeed Sulzbacher and McLean (1951) , who studied
American pork sausages, stated that little was known about
the types of micro-organisms present in fresh sausage despite

13
several reports dealing with their numbers (e.g. van der Slooten,
1907; Savage, 1908; Cary, 1916). An examination of 316 isolates
of pork sausage by Sulzbacher and McLean (1951), who based identi
fication of isolates on descriptions in the 6th edition of
Bergey's Manual (Breed et al., 1948), revealed that ca. 74%
could be assigned to six genera (Table 2). In the light of the
present study, several features of this report need comment.
The large proportion of Proteus spp. is misleading because
these organisms were isolated from Salmonella-Shigella agar
after enrichment of sausage in tetrathionate broth. The majority
(70%) of pseudomonads were active producers of lipases and
proteases and, although they were isolated frequently from
freshly-made sausage, they could not be recovered in large
numbers from the stored product (Table 3). Microbacterium sp.,
which formed large populations in the chilled product, were
implicated in the production of acidic flavours. All 47
isolates of Microbacterium comprised one species that differed
from any of those given in Bergeys Manual (Breed et al., 1948).
These non-mptile. Gram-positive asporogenous bacilli were unable
to reduce nitrates to nitrites, were catalase-positive, gelatin-
negative and produced lactate and carbon dioxide from carbo
hydrates. A later report (McLean and Sulzbacher, 1953)
assigned these organisms to a new species, Microbacterium
thermosphactum. The original report (Sulzbacher and McLean,
1951) did not state whether or not sulphite was included in
the sausages.

14
Table 2. The microorganisms isolated frcm American pork
sausage*
Genus Number of isolates % of total
Bacterium 65 20.6
Microbacterium 47 14.9
Achromobacter 40 12.7
Pseudomonas 34 10.8
Bacillus 28 8.9
Proteus 21 6.7
* Adapted from Sulzbacher and McLean (1951)
Table 3. Types and distribution of micro-organisms in sausageand on equipment^
Genus Freshsausage
storedsausage Spices Equipment
Pseudomonas - - +
Microbacterium ++++ - -
Alcaligenes + ++ - -
Achr omobacter +-(- ++ - 4-
Bacterium -F-l-f ++ - 4-
Bacillus 4- + 4-4- 4-
*Now identified with Brochothrix thermosphacta (Sneath and
Jones, 1976)
■(-Adapted fran Sulzbacher and McLean (1951) .

15.
> 1
3

15.
British fresh sausage
The meat ingredients infect fresh sausage with those organisms
(Table 4) which occur in large numbers both in the stored product
and in the abattoir environment (Ayres, 1955). Dyett and
Shelley (1962, 1966) were the first to examine the effect of
sulphite on the keeping qualities of British fresh pork and beef
sausages. They identified the majority of their isolates from
sulphited sausage with Bacillus, Micrpcoccus or Streptococcus;
the absence of Microbacterium thermosphactum was probably due
to the incubation of the isolation medium (Plate Count agar)
at 30°C. The first evidence that a particular microbial associ
ation was a common feature of British fresh sausages was provided
by Dowdell and Board (1967) who established also that an "active
fermentation" began in sausages which had been stored for
> 24 h at 4^ or 22°C. During the first 24 h of storage at
4°C, the numbers of the Gram negative, aerobic bacilli-the
numerically-dominant group at time of manufacture - diminished,
and rapid multiplication of Microbacterium thermosphactum, yeasts
and lactic acid bacteria ensured that the characteristic Gram
positive/yeast association was formed within 48 h. Further
surveys showed that (1) pork sausage was similar to beef sausage
in terms of the level and type of contamination, (2), there was
an extensive range of contamination (3), the counts in
particular brands of sausage fell within a part of the overall
range and (4), the scatter of counts of a particular brand
increased with an increased total viable count (Dowdell and Board,
1968). The initial microbial load was an important determinant
in the selection of the microbial association. Using this 4th

17.
0)E
W<uu
oc0
•H4->fO•HUsin(T3fd•H120uu
•HE(U
CnC•Hin
•H
Iu
in4Jc:
•H
C8(wO
§•HVs•l-<iw•Hmw(d
>1u
a•H12<
CÜf—I •§
*0)
cnDSW0)
cwJGin•rHu-r4a%S4o■pin
■8Id
PId
Æ
I§Ü
•H
•HC•S'W
c.2-UuQ)
iwC•rH
U-to
u
CMCM
U
CD 0 > •0) m
Id•H
•iMc
NU
I4JIdu
+ + + +
+ I + +
CM m 22 X X
in ^ 0 0g g g gr-4 r"4 r-4 r—I
I I
i4JÜ
I-c: •4J A
"M "MS: 5:
41Cg•HI
+1 +1
+1 +1
CM
8 8
tn•H eM 3(d •M0 ÜA Q) Q)û, <d iden >44en W (n3 3 3M Ü üM ü ü•H 0 0 enü 0 V Sfd 0 0 fi4 en 4J 4J 00 41 Q. Q. (444J çn <Ü <D •HÜ id k, k, rHid Q 4J 41 0>1 tn u
in CM CM CM iH O22222 VV V V V V
âÀ A Û4 eu04 A eu Ck
•H en en tn en (4-M 04 tu>44 en ü Û4 tn 41% 3 0 en (d Ü0 ü 41 •H G fdN 0 en tn V 12
0 0 3 'H 0*H ü G M (4 0 4113 0 0 M 41 'a41 •H ü "M tn 3 G44 '3 3 ü 0 ty •HP Q) <Ü <d M tn 0
eu KJ pq U eu
ug•Hs
CTi
'Bidg73Gid
IRgU
IH
T33T3<
> 1•Hc0en0)f-Ha.
G•HV.
&
1G•HIOu
•HS0)12G
•H
en•H54->Id
1£

18,
criterion, Dowdell and Board (1968) separated 40 samples of
pork and beef sausages into the following categories: Type
A (20% of samples) had an initial microbial load of 5 - 10 x 10^
micro-organisms/g and was dominated numerically by yeasts; Type
B (15%) contained initially 1 - 10 x 10^ organisms/g and was
dominated by unidentified Gram positive bacilli; Microbacterium
thermosphactum and micrococci comprised a small fraction only
of the microflora; Type C, the most commonly occurring (60%
of samples) microbial association, was formed primarily of
Microbacterium thermosphactum, although small numbers of other
Gram-positive bacilli, yeasts and Gram negative bacilli were
present also. Sausages of the last category, which had an initial
level of contamination of 1 - 100 x 10^ micro-organisms/g,
appeared to be produced consistently by large manufacturers.
Local butchers, on the other hand, produced another category
of sausage (type D), having a different type of microbial
association. The very high initial level of infection (1 - 10 7
X 10 micro-organisms/g) consisted mainly of aerobic Gram
negative bacilli, identified with the 'Pseudomonas - Achromobacter
complex'.
Dowdell and Board (1971) noted also that on storage at
refrigeration or room temperature the majority of pork or beef
sausages favoured the growth of one or other of two microbial
associations. Thus with storage at 4°C, Microbacterium thermo-
sphactum and yeasts became dominant whereas at 22°C Microbacterium
thermosphactum, yeasts, lactic acid bacteria and micrococci
grew. The microbial contaminants were assigned to one of three

19,
categories, dominant, major or minor. These distinctions were
made not only with regard to the size of the initial infection
but to the fate of the organisms in the stored product also
(Table 4). Although it has been demonstrated unequivocally
that microbial associations exist in commercial sausage
(Dowdell and Board, 1967, 1968, 1971), other worlcers haye
failed on occasions to verify this finding (e.g. Ashworth et al.,
1974). Perhaps the main reason for such differences was that
the latter studied sausages produced on a very small scale in
a test kitchen. It has been established that the latter product
is markedly different, particularly from a microbiological
standpoint, from the typical commercial sausage (Brown, 1977).
Further support for the existence of microbial associations
was provided by Brown (1977), Abbiss (1978) and Leads (1979).
They confirmed that Microbacterium thermosphactum dominated
the microflora of the majority of commercially-made pork
sausages, and that growth of lactic acid bacteria was stimulated
by a high initial microbial load. Furthermore, they demonstrated
that the characteristic ’yeast-dominated' miçroflora, which
was associated with a low initial bacterial count, was a
factory-specific phenomenon. Although the last mentioned
group of workers agreed with the contentions of Dowdell and Board
(1967, 1968, 1971) that microbial growth was faster and greater
at the surface than at the centre of sausages, they did not
give evidence in support of the conclusion that lactic acid
bacteria grew more extensively at the last mentioned site
(Dowdell and Board, 1971).

20,
Thus it is evident from the literature that a particular group
of micro-organisms tends to dominate the microbial flora of
stored British fresh sausage and, although Brown (1977)
provided evidence that the situation was caused by strong
selective pressures, these still await definition. The role
of sulphite in the selection of the microbial association was
therefore studied in detail (pp.125 - 151).
Enterobacteriaceae
The family Enterobacteriaceae may be defined as a group
of Gram-negative bacilli that are either motile - peritrichous
flagella - or non-motile, which grow aerobically or anaerobically
on simple media and MacÇonkey's bile-salt-lactose medium. They
are oxidase-negative and, with one exception, catalase positive,
capable of reducing nitrates to nitrites. They ferment glucose
in peptone water with the production of either acid or acid
and gas, and degrade glucose and other carbohydrates both
fermentatively and oxidatively.
In 1937 Rahn proposed the family name Enterobacteriaceae
to include organisms which had been assigned to the genera
Escherichia, Salmonella, Aerohacter, Klebsiella, Proteus,
Erwinia, Eberthella and Shigella. The new family included
also strains, of Serratia, Pseudomonas, Flavobacterium and
Achromobacter which fermented glucose with the production of
gas. All these "taxa" were placed in a single genus, Entero-
bacter. With the adoption of the first Bacteriological Code

21.
(Buchanan et al., 1948) both Enterohacter Rahn, 1937 and
Enterobacteriaceae Rahn 1937 became illegitimate because they
did not conform to the rules which were made retroactive to
bacterial names (Anon, 1951). Although the Judicial Commission
(Anon, 1958) chose to conserve the name Enterobacteriaceae Rahn
1937 as Enterobacteriaceae Rahn 1937, nom. fam. cpns. (Opin.
15, Jud. Comm., 1958) it was not included, however, in the
Approved List of Bacterial Names (Skerman et al., 1980) and
was considered (Lapage, 1979) to have been formed in contravention
of Rules 9 and 21a of the Code (Anon, 1951) . The proposal
(Lapage, 1979) of Enterobacteraceae as an alternative to
Enterobacteriaceae has been opposed (Farmer et ai., 1980)
because the former has not been validly published and thus has
no standing. Furthermore, proposals to replace Enterobacteriaceae
Rahn, 1937, nom. fam. cons. (Opin. 15, Jud. Comm., 1958) with
Enterobacteriaceae fam. nov. nom. rev. (Ewing et ai., 1980)
or with Escherichiaceae (Goodfellow and TrUper, 1982) have been
made. The genera currently assigned to the family Entero
bacteriaceae are given in Table 5. In view of the confusion
surrounding the choice of family name noted above, the term
Enterobacteriaceae will be used in this thesis, because of its
common use in food microbiology.
Sources of Enterobacteriaceae
From the standpoint of contamination of pork with Entero
bacteriaceae (Table 6), five main depots of infection can be
considered :

22
Table 5. Members of the family Enterobacteriaceae’
( Buchanan and Gibbons, 1974) Genera not included in Bergeys Manual (Buchanan and Gibbons,1974 but proposed recently
Ci trohacter Cedecea
Enterohacter Rahnella
Escherichia Buttiauxella
Hafnia Levinea
Klebsiella Obesumbacterium
Proteus Tatumella
Salmonella Kluyvera
Serratia Xenorhabdus
Shigella Providencia
Edwardsella Morganella
Erwinia Branhamella
Yersinia Ewingel la
* General discussions of the taxonomy of this family have been
presented by Johnson et ai. (1975); Sakazaki et ai. (1976)
and Sackin and Jones (1976).

23.
(Uuso CD
g
u
<jO 43 ifl
U
m jQ
II
I
V.
aex U1tn U k
k u H) Q)k 9) •H 4J c(U 4J C 'a Ü Q)
U 0 c IQ &>U s 3 4 Ou •Q (U 0 k
•9 P e U 0)p <ts 01 IQ«H 4J0 C< U co
(0 t»(0 c 0 aC 0 4J U) aifl 0 tnk u u ig
Q> ,<g <g <8 Cl1 p u < a <g0 ü (ü ü tn c
5 3 18 •'H Cl Cl Cl&> 0 ty k «g ig3> M C (U 0 "4 ti) Nig u ■g Ü 5 ig •§ 0
Cqw igcq % 3
igü
tnc0 tn
•4 ca U p ig a 0tn .<g ü Cl tn ü
(0 <g 0ig <v p •o ig kl
3 ü Clty V x> 3ig s 3 cu ë kl tn Clk, uCl Clto >1

24
I«jS(flMâ
I
43XUaT3
3a-s4J(Q
3 «»»»
II
0)a"Os2o>
Cl 73O ' Cl<0 M(1) U3 flfl aU) 1444< fl4 Cl 3g e Ua f l
73 > 0)Cl 0>
43 J< fdU in 43 cnfl 44 3<44 a f l fd
k Cl 1 m U) cnCl Cl E 4C
(D O' 43 kl x :E H 3 in 0 en2 3 73 U <44 3 3 o>42 A C fl Cl 0 uE 3 > g •4 44 U-l
73 2 u 0 kl f l0 P 43 0 l4 73 fl Cla O ' C 73 > E3 73 fl C
0 ■4 73 3U1 U c fl l/l 0 fl
0 C 44 7J 44 kla •4 3 kl O'0 »4 a <44 o \ 0 4 ) C
Cl r~ S fdŒi <T> Cl CTi CO r 4 r 4 0)r4 tN 43 r 4 44 r - 00 CD Hi n f l O ' en <7% ü
0 en Cla 0) Q X Ein co 0 TDCl m u tn O' O' Cz (4 m Cl z Z (du 7J 732 73 C en 73 t3 Mc fl 44 C C 0>
Cl iQ Cl 7J w fl <TJ x :•4 üU1 tn C 43 44 c 73 V) «d% Cl Y Cl 3 0 c Cl 0) X3
0 in U) U 44 fl .4c u Cl V4 ï •4 •H #—4Cl •4 Cl CI Cl O ' 44 4J 3M X "3 4) s z z W W W

25
1. Animal feeds and ingredients are frequently contaminated
with Enterobacteriaceae (Patterson, 1969; Edel et al.,
1973; Stott et al., 1975). Levels of contamination of
upwards of 1.3 x 10^ Enterobacteriaceae/g have been
implicated in the spread of Serratia, Klebsiella, Citro-
bacter, Proteus and Escherichia in a piggery although
pelletting of feeds can lead to a 10^ - 10^ reduction in
the numbers of the organisms (Mossel et al., 1967).
2. Most systems of pig husbandry offer little impediment
to transfer of micro-organisms between animals. The
skin of pigs may harbouf a significant number of
Enterobacteriaceae and the faeces upwards of 10^/g
(Willingale and Briggs, 1955). In addition to direct
contact between pigs, infection via faeces, dirt, air and
drinking water has been demonstrated (Willingale and
Briggs, 1955).
3. Workers and
4. Equipment in the slaughterhouse and processing plant will
be involved in the dissemination of Enterobacteriaceae.
Chill rooms may contain appreciable populations of
psychrotrophic Enterobacteriaceae (Newton et al., 1978).
5. As the gut harbours a large number of Enterobacteriaceae,
slaughtering techniques play an important role in the
transfer of organisms from a depot to the butchered
meat. For example, heavy contamination of the latter
can occur when the gut is punctured during evisceration
and excision of the rectum can lead also to heavy
contamination (Gerats et al., 1981).

26
The inferences drawn from analyses which showed heavy
contamination of uncooked meats with Enterobacteriaceae have
tended to be influenced by two assumptions: (1) that heavy
contamination is likely to be associated with the presence of
Salmonella spp., and (2) that heavy contamination is basically
an indicator of poor hygiene during slaughter and butchering.
The well founded precept that lactose-fermenting Gram-negative
rods - coliforms - and particularly Escherichia çoli in water
is indicative of recent faecal contamination and therefore of
the probable occurrence of Salmonella and Shigella has been
accepted uncritically by some food microbiologists (Anon, 1976).
Although the MPN coliform index is widely used by them, certain
features of the technique - the statistics, interpretation of
results and range of organisms isolated - have not been sufficiently
stressed. For example, the occurrence of false positive results
in water examined for coliforms have been noted repeatedly
(Meyer, 1918; Sears and Putnam, 1923; Leitch, 1925; Thompson,
1927; Koser and Shinn, 1927; Greer and Nyhan, 1928; Andrews
and Presnell, 1972; Dutka, 1973; Bissonette et a i . , 1975
and Dutka and Kwan, 1978). Undue emphasis has been given to
the isolation of commensal or saprophytic micro-organisms
which may or may not be members of the Enterobacteriaceae. Thus
Wilson et a i . (1935) isolated genera of Enterobacteriaceae
which are associated principally with vegetation and Hussong
et a i . (1981) identified several aberrant, intermediate groups
("unclassified new species") of Enterobacteriaceae as well as
members of the genera Pseudomonas, Bacillus, Aeromonas and

27.
Staphylococcus. Although the plasmid borne character (Sackin
and Jones, 1976) of lactose fermentation has been a key feature
of the indicator organisms used in food or water microbiology,
only a few food-borne pathogens of faecal origin have this
property e.g. some Salmonella serotypes of sub genus I
(Threlfell et ai., 1983) and III - the "Arizona" group. Thus
some food microbiologists (Mossel, 1957; Mossel and Ratto,
1970; Newton, 1979) recommend an examination for all rather
than merely the lactose fermenting members of the Enterobacteriaceae,
This is achieved by using glucose instead of lactose in isolation
and enumeration media.
Psychrotrophic Enterobacteriaceae (e.g. Enterohacter,
Hafnia and Serratia spp.), occur and probably grow on unpreserved
meat and meat products stored at chill temperatures.
However Newton and Gill (1978) reported that large numbers
of lactobacilli on anaerobically stored meat inhibited the
growth of a species of Enterohacter. Furthermore, Sutherland
et al. (1975 a,b) noted that Gram-negative, fermentative bacilli
comprised a small fraction (ca. 7%) only of the microflora of
vacuum packed meat. The pH of the substrate also influences the
rate of multiplication of psychrotrophic strains of Enterobact
eriaceae (Newton and Gill, 1980) and Grau (1980) was of the
opinion that the type of anion causing a reduction in the pH
was an important determinant of the rate of growth of strains
of Enterohacter cloacae, Serratia liquefaciens and Yersinia
enterocolitica. In comparison with strains of Pseudomonas,

28.
the relatively slow rate of growth of the individual contaminants
at chill temperatures is the principal factor which selects
against the establishment of large populations of Enterobacteria
ceae on aerobically stored meat (Gill and Newton, 1977). Thus
at temperatures below 20°C, pseudomonads - especially Pseudomonas
fragi - tend to dominate the microbial blooms on meat (Shaw and
Latty, 1982) and in meat products (Molin and Ternstrom, 1982).
The observations by Brown (1977) that presumptive Entero
bacteriaceae grew in unsulphited sausages but to a limited extent
only in sulphited ones led to a detailed study of the actual
changes in the numbers as well as the types of these organisms
in sausages or media having known concentrations of sulphite.
In addition, the search for glucose rather
than lactose fermenting organisms in the study concerned with
Salmonella contamination of sausages and ingredients was done
with the objective of assessing the utility of the Entero
bacteriaceae count as an index of contamination of the product
with these food poisoning organisms.
Brochothrix thermosphacta
As little is known about the natural niche of Brochothrix
thermosphacta (Microbacterium thermosphactum) and, until recently,
its taxonomic relationships were confused, this section gives
particular emphasis to the classification of this organism and
the factors which appear to favour its growth in meat and meat
products such as British fresh sausage.

29
Nomenclature and taxonomy
The genus Miczobacterium was proposed for heat resistant.
Gram-positive, catalase positive, asporogenous bacilli which had
been isolated from dairy sources, especially pasteurized milk and
milk products (Orla-Jensen, 1919). Four species were defined:
Microbacterium lacticum, flavum, mesentericum and liquefaciens.
In the 5th edition of Sergey's Manual (Bergey et al., 1939) the
genus was assigned to the family Bacteriaceae but was transferred
to the tribe Lactobacillaae within the Lactobacteriaceae in the
6th edition (Breed et al., 1948). Microbacterium mesentericum
was renamed Nocardia mesenterica and Microbacterium liquefaciens
was removed to the appendix (Breed at al., 1948). The 7fh
edition (Breed at al., 1957) excluded Microbacterium liquefaciens
and established the genus Microbacterium, comprising two species.
Micro bacterium flavum and Microbacterium lacticum, within the
Corynebacteriaceae. The genus was extended to accommodate
Microbacterium thermosphactum (Buchanan and Gibbons, 1974) despite
the fact that neither Microbacteriqm lacticum nor Microbacterium
flavum were related closely to this nev/ species (Table 7).
On the basis of its biological properties (cell morphology,
staining reactions, cellular inclusions, GC content of DNA,
degree of DNA homology with Corynebacterium diphtheriae PW8,
and pattern of enzymes) and chemical structure (peptidoglycan
type, cell wall polysaccharides, phospholipids, glycolipids
and fatty acids), Microbacterium flavum Orla-Jensen (1919)
has been renamed Corynebacterium flavescens (Barksdale et ai.,
1979) and is recognised by the latter epithet in the Approved

30.
*(0•H
5 * •§•rj6(NOmu•Hw•HO4,1ggÆüS'•HPffl•Hg0)8<44(W•HQ
r-0)rHI
CMr~03m 00 1—1in kO 0103 en rH1—1 1—1 uO M >100 kO G eikO S4 03 0)03 01 ,—1 01 4J <r4 x: X 01 U0 41 E-
01 rd en Gu A G (d 0 01G M N id X en Xi0) (Q r—1 > A 41X 0 w V E01 4J w G 0 <4404 01 T3 id x: 001 00 G 64os G (d G 1 en0 0 en 01en G rH en G SG) 01 T3 •H-r4 (U A •H iH N> •H > 1—1 GO 01 0 0 01Q Z Û Û u G•H
0141idE E3 idÜ 41'N 03 04J r* r—1Ü 4- ü kp 03 031 1 + + 1 + r4iQ 00 <44V3 00 M enG •Hro enS: N 41 01ü >*4 01 ÆH 41% 01 G% rM >164 0 enU D 0g en •HCQ mO k4% Idp E mH a mS; 4J 000 m 41 03
2 0 G rH-G kO -H- 10Gi + + 1 1 m 1 41tfl 13 en >1Q 'H 41S rd en G>4 01 •r4O kl Id-G Û4J 41id01 miGrH os00 4103 0 T31—1 G G01 0I—1 EV kl 3 >1>1 01 3 > 1—1ü rH G 10 1—1ü no M q
Xi >i kl <k| zp u (d3 U E0 < 3 Qkl 0 4J 41 y E •Htm •H G en A 0 kl >1
Xi (0 01 kl 01 130 eu 4J 41 en iH iw 41•H 0 W 01 id 0 T3k r: kl •H 01 u G T3 2 0101 A 4J en G P 0 01 -G 41p k4 0 01 •H kl •H 41 0 00 0 kl kl Æ 41 Qi kl Qi0 g Xi 1 •H U cW> G id Ü enkl 0 V +J N 0 kl T> •H(0 01 >4 Id G 41 < 01 < §; QÆ r—1 en 01 01 >1 Z Ûiu eu eu X CQ U Q O * +~ -H-
ü)w01aG•r4
o•H■§
S
G(ütm
fOt)IIXa4JIIG•H

31
List of Bacterial Names (Skerman et al., 1980). Microbacterium
lacticum is considered by some to be misplaced in the genus
Microbacterium and a proposal to rename it as Aureobacter
liquefaciens nom. nov. within the family Corynebacteriaceae
has been made (M.D. Collins, pers. comm.). The napie Micro
bacterium thermosphactum was coined by McLean and Sulzbacher
(1953) for a heat sensitive contaminant of fresh pork sausage.
This organism was not included in the 7th edition of Bergey's
Manual (Breed et al., 1957) but was assigned to the coryneform
group in the 8th edition (Buchanan and Gibbons, 1974). Its
taxonomic position remains uncertain, it being considered as
a species incertae sedis in the current Approved Eist of Bacterial
Names (Skerman et al., 1980). Scxne workers have suggested that
Microbacteriqm thermosphactum is probably related to the lactic
acid bacteria (Diebel and Evans, I960; Barlow and Kitchell,
1966) whereas others (Davidson, 1970) have supported its inclusion
within the Corynebacteriaceae. The absence of unsaturated and
cyclopropane fatty acids, both of which are commonly found in
lactobacilli, from the lipids of Microbacterium thermosphactum
(Shaw and Stead, 1970) and the presence of a functional cytochrome
system (Davidson and Hartree, 1968; Kelly and Dainty, unpublished
observations) would lend support to the views of the latter;
The exacting nutritional requirements (London, 1976) in the
absence of ammonium ions and cell wall type (Schleifer, 1970;to the former. Sneath and Jones ( 1976 )
Schleifer and Kandler, 1972)/recognized the unsatisfactory
classification of this organism by McLean and Sulzbacher (1953)
and assigned it to a new genus, Brochothrix, containing one
species. They proposed also that the new genus be assigned to

32
the Lactobacillaceae. These proposals were supported by Wilkinson
and Jones (1977) who, in a taxonomic study of Listeria and related
organisms, demonstrated that, although Microbacterium thermo
sphactum was related to the genus Lactobacillus, it formed
a distinct phenon worthy of generic rank. The supposition that
Microbacterium thermosphactum was related to Kurthia (Buchanan
and Gibbons, 1974) was not supported by others (Davies at ai.,
1959; Jones, 1975; Wilkinson and Jones, 1977; Shaw and
Keddie, 1983).
Niche
Despite the numerous reports of the occurrence of Brochothrix
thermosphacta in many types of meat product, its precise ecological
niche is not known (Gardner, 1981). Brochothrix thermosphacta
has been isolated from lairage slurry, cattle hair, rumen contents,
slaughter hall soil, hands of workers, carcasses and chill room
air (Patterson and Gibbs, 1978). The common occurrence of this
organism on the hands of workers, equipment and carcasses
during "boning-out" operations (Newton et al., 1978; Patterson
and Gibbs, 1978) indicate that ample opportunity exists for the
transfer of this organism from these sites to the meat or products
made therefrom.
Muscle post rigour is likely to contain sufficient L-lactate
to prevent anaerobic growth of Brochothrix thermosphacta on
chilled carcasses but the ability of the organism to multiply
in the presence of this acid under aerobic conditions is well
known (Grau, 1980). Despite the absence of esterases capable

33
of degrading fats containing fatty acids > (Collins-Thompson
et al., 1972), glycerol is readily metabolised under aerobic
conditions. Lipolytic action of meat or other microbial systems
would thus appear to be a prerequisite, however, for the growth
of Brochothrix thermosphacta on this substrate. Carbohydrates
would appear to be the preferred metabolites; high glucose con
centrations and an acidic pH favouring acetoin and acetic acid
production, low glucose concentrations, causing the accumulation
of isobutyrate and isovalerate from the breakdown of valine
and leucine (Dainty and Hibbard, 1980). Neither the pH nor
the temperature of carcass meat is likely to curtail the growth
of Brochothrix thermosphacta. Although the optimum pH for
growth is 7.0, it will tolerate a range of pH (5.0 - 9.0; Brownlie,
1966) and proliferate on carcass meat of pH 5.4 - 5.5 (Patterson
and Gibbs, 1977). Brochothrix thermosphacta will grow at 0°C
(Brownlie, 1966) and has a high energetic efficiency over a
wide range of temperatures (Rogers et al., 1980).
The advent of wrapping films having a range of gas permeability
(Barlow J<r Kitchell, 1966; Gardner et al., 1967; Shay et ai., 1978)
and their use in modified-atmosphere packaging (Weidemann, 1965;
Davidson, 1970) led to the recognition of Brochothrix thermo
sphacta in the spoilage of meat and meat products. Some early
reports noted the presence of "coryneform bacteria" or Gram
positive bacilli on meat (Haines, 1937). Such organisms have
been found on a variety of meats: beef (Rogers and McCleskey,
1957; Wolin et ai., 1957; Ayres, I960; Weidemann, 1965),
poultry (Thornley, 1957; Barnes and Shrimpton, 1968; Barnes

34,
et al., 1979), lamb (Barlow and Kitchell, 1966; Newton et al.,
1978) and pork (Gardner et al., 1967; Gardner and Patton, 1969).
Comminution of meat, as in the production of sausage, appears to
result in an increased level of contamination with Brochothrix
thermosphacta and large populations have been reported in American
pork sausage (Sulzbacher and McLean, 1951; Miller, 1964) ,
irradiated frankfurter (Drake et ai., 1958) and British fresh
sausage (Gardner, 1966; Leaton, 1968; Dowdell and Board, 1967,
1968, 1971). Cured sausage rarely contains B. thermosphacta,
a situation which is probably attributable to the sensitivity
of the organism to the NO^ ion (Gardner, 1981).
In the present study, Brochothrix thermosphacta was included
in the studies concerned with sulphite tolerance of members of
the association of British fresh sausage with the objective of
assessing the contribution which this organism's tolerance to
the preservative makes to its success as a colonizer of British
fresh sausage.
Yeasts
The literature on the sources, types and behaviour of yeasts
in meat and meat products is scant (Walker and Ayres, 1970).
Their metabolic activity and biomass (Rose, 1976) would be ex
pected to cause perceptible changes in foods and on occasions
to favour growth of and spoilage by other micro-organisms.
In practice, several factors are known to influence the ability
of yeasts to compete with or aid other micro-organisms in food

35.
e.g. numbers and types of yeasts, availability of nutrients, pH,
redox potential, temperature, water activity (a ) and presence
of preservatives (Ingram, 1958).
Yeasts are ubiquitous (Lodder, 1970) but seldom cause
spoilage of fresh red meats because of their slow rates of growth
at refrigeration temperatures (Ayres, I960), and their low level
of initial contamination (Table 8). If bacterial growth is
restricted, by irradiation or by antibiotics, yeasts may attain
large populations on carcass or processed meat (Barnes et ai.,
1979). Low a^ is another well-known elective factor (Scott, 1936, 1957)
which operates in products such as salami, pepperoni, cervelat
or Thuringer(Cesari, 1919; Cesari and Guilliermond, 1920).
Some asporogenous, lipolytic yeasts do grow on meat, especially
the fatty tissue, at low temperatures (Lea, 1931 a,b/ Vickery,
1936) and they can attain populations comparable numerically to
those of bacteria. Indeed Ingram (1962) was of the opinion that
the importance of these lipolytic yeasts in the spoilage of meat
had been generally underestimated by food microbiologists.
Reports of yeast contamination of sausages generally refer
to products which have been heated, dried, smoked or fermented.
Thus, Oglivy and Ayres (1953) reported a yeast-dominated microflora
in packed frankfurter (a cooked, smoked product). Large
populations of yeasts, especially on the outer surface, were
associated with the spoilage of skinless pork sausage - a partly
cooked version of British fresh sausage (Hockley, 1980; Legan, 1981).
Yeast production of slime on sausages is well documented viz.

36.
fl flÜ
w fl flc fl .4fl V 3 0ü fl .5 ü Q)•4 10 0k, c E 3 û .M 3 fl 3 >4fl Ü 0 < S
Ql (073 ex 10 3■4 10 3 U0 10 ÜC w 3 k,fl 3 7? k fl'S Ü b>c 3» G 4 (0 c:•H 0) 0 fl 3N u fl 42

37.
+ +
E î3
W 10 a ;C 0c fl 10
<u k 4) my V3 U 3ifl fl m EE E §1k
4
a,3 ex (00 (0•H E Qi•H 10 3 10N 0) •4 •4% u 73 44 flQi 3l 44 ■443 E kl 0 33 0 Q Ü kl<0 k 5. 10 0<Ü 10 44t» 0 03» ü '93 3 0M % 4=X d S

38,
U)Q* fltn E cfl fl 3 flfl 73 u t; (U
•4 fl 0 <u c 3fl 73 fl k, flE C k 0 •4 44 Mfl fl (Ü 0 , Q> 3 3Ü > fl 43 Ü cx
>1«4 7344 CH f l3 0)Q 73 ÜQ< 03 c
4473 •4 EC 42fl ex 144rH 03
42 3 03 flfl fl 43 k4•4 03>44 44
44 fl f l klf l C C 3
44 0 0 144f l •4 400) 44 44 CB 73 f l f l f l
03 > > k l73 U M l4 <4403 00 c 03 03k l in f l flO' e 43 43 O'rH 0 0 inO O't-' o 73 73O' , c 00 fl 03t—1 c O' £ 03 JZ
f l f l O tn 00E •4 f l •4 m
f l fH i4 f l rH O'01 03 43 3 43 rHk l 73 3 fl 3fl fl CX fl CX<■ 44 c cV3 44 3 73 373 03 03C 73 C 44 Cfl Ç C 0 0ia Ç 44 42 44 03k l fl CX01 fl E fl .4 fl 03
r4 Q 3 Qr-f '—1 fl flfl 0) C k l3 3 Q tc 3 £ o00 O' O
rH
f l03C
•r4rHV>1Vf l
f l k l >144 44 klU 03 443 44 r~i
73 k l 30 0 Qk l ( XC^ 42
U<41 44 <4403 f l 42 03 44
03 44 03 f lA E •H 43 03
3 E42 73 73f l C 73 03 7303 f l 03 03k l 44<44 44 f l
f l 03 4203 k l 13 42
44 m E 44 03 Uf l fN| 0303 O ' C E 43E rH 03 f l m c n
N ' 0 O ' i n73 0 03 r4 O '03 73 kl rH rH
rH k l <44 44rH 0 f l•H <44 03 42 M f lf f l O E (4 f l 03o c CO kl
f l O ' 44 >1s CO O 03 <
o mkO 73 O ' O ' 73O ' C 44 iH 43 Cr 4 f l kl o f l
0 1f l Q i S k l 3 kl
f l 40 C f l 03 0303 Q 03 k4 73 3 Mk l 0 > C3' ■o 0 rH>1 k l fl c 5 fO
< OQ Q M P 2 3
l£3

39.
saucisson (Cesari, 1919; Cesari and Guilliermond, 1920), weiner
(Mrak and Bonar,1938; Mrak and Phaff, 1948) and British fresh
sausage (Dowdell and Board, 1971).
The resistance of yeasts to sulphur dioxide or sulphite
(Rehm and Wittman, 1962; Dennis, 1978; Dennis and Harris, 1979 )
has led to the surmise that their growth in British fresh sausage
may be favoured (Brown, 1977) but Dyett and Shelley (1955) noted
that, in most instances, the climax populations of yeasts in
sulphited sausage were 1 - 2 log cycles lower than those in the
unsulphited product. Their conclusion that sulphite at 450 ppm
inhibited the growth of yeasts was not in accord with the many
observations of others (e.g. Dowdell and Board, 1967, 1968; Ashworth
et a.i., 1974; Brown, 1977). Brown (1977) was of the opinion
that yeasts were mainly responsible for producing sulphite-binding
agents and that the growth of some of the bacterial contaminants
may be dependent upon the consequent reduction in the concentration
of free sulphite. This observation, together with the discordant
views on the influence of the preservative on yeast growth per se
led to the present study of a number of yeasts for sulphite-
tolerance and potential to produce sulphite-binding compounds.
Sulphite preservation
Although the salts of sulphurous acid have been used as
preservatives in a wide variety of foodstuffs (Schroeter, 1966;
Roberts and McWeeny, 1972), they are of particular value in
products having an acidic pH reaction, e.g. cider (Burroughs and

40.
Sparks, 1964), orange juice (Ingram and Vas, 1950 a,i>) fruit
pulp (Robson, 1968), jams (Dennis and Bugahiar, 1980) and wines
(Faparusi, 1969; Okafor, 1975; King et al., 1981). This
situation obtains because SOg, the most antimicrobial moiety
formed by dissociation of sulphurous acid is at a maximum
concentration (Hammond and Carr, 1975). As SO^ is a particularly
reactive molecule (Joslyn and Braverman, 1954), having the potential
to combine with many compounds in aqueous solution (e.g. aldehydes,
ketones, olefins, sugars, organic acids, thiol groups, enzymes,
cofactors, vitamins, nucleic acids, amino acids and lipids,
Baird-Parker, 1980) , it is used also as an antioxidant (e.g.
in potatoes, Lund, 1968) and an inhibitor of enzymic and non-
enzymic (Burton et ai., 1963), browning in pickles and confectionary
(Sullivan, 1971), and vegetables (Moussa, 1973). It needs to be
stressed that the vast literature on the mechanism of SOg-
preservation in acidic foodstuffs has tended to direct attention
away from the situation which obtains in sulphited products poised
at, or near, a neutral pH. In the light of the present study on
sausage preservation only the literature pertaining to the use of
sulphite in "neutral pH" meat products will be considered in
detail.
Sulphite preservation of meat products
Metabisulphite or sulphite is included in fresh sausage primarily
to delay microbial spoilage. The content of preservative in the
product has been determined traditionally by collection of the
sulphur dioxide (SC^) released from a sample suspended in boiling

41
acid (Monier-Williams, 1927). Indeed this analytical procedure has
led to the general assumption that SO^ per se is the only anti
microbial moiety. In practice, the pH of sausage (6.8 - 6.2) would
not be expected to cause significant SO^ formation from meta
bisulphite and sulphite (Table 9, Ingram, 1948; King et ai.,
1981) and it is probable that bisulphite (HSO^ ) and sulphite 2 -(SO^ ) are the active agents (Hammond and Carr, 1976). Vas and
Ingram (1949) were probably the first food microbiologists to
establish that the pH of a food product was an important determinant
of the efficacy of SO^ and they published a set of curves showing
the proportions of the various molecular species based on the value -6of 5 X 10 for K^ (2nd dissociation constant). Although these
curves have been reproduced widely in the literature - e.g. Rehm
and Wittman, (1962) - King et al. (1981) were of the opinion
that they are in error by two full pH units (7.2 vs. 5.3) at the
pK^ point. But even with their calculated percentage (Table 9)
of each molecular species of undissociated sulphurous acid (H^SO^)- 2in the range of pH, 7.5 - 5.0, based on a K^ of 1.7 x 10 and
-8a K^ of 6.31 X 10 (pK^ = pH 1.77 and pK^ = pH 7.20), SO^
forms a negligible proportion. Although it is well known2-that the absolute content of SO^ /HSOy in sapsage diminishes
with time due to oxidation, little is known about the extent of— 2 — — 2 —binding of HSO^ and SO^ . Oxidation of HSO^ and SO^ to
2-SO^ is well documented (Abel, 1913; Anderton and Locke, 1956;
Fridovitch and Handler, 1961; Hibbert, 1970) and may account
for the irretrevable loss of preservative from stored sausage
studied by Brown (1977). It has been established also that2-reversible combination of SO^ with pyruvate (Burroughs and

42
♦
AIM0a■S
1(/)ni
3%in
2II— I3iniw0in
Ü&ki(ÜyÏ
r—4
E<w0§•H-Q•Haw•H'O
(UCnfOVcH)Üu(UOh
o >
<ur-4As
ICM
CO CO CO LO o OO CO sr COW
6 1— i m KÛ CO VOdP 1-4 CO VO
CM
A
r4 in in m 0 OCO 0 p—1 in COcn 00 CO 1—1 CO<r> 00 VO CO
sO
O m Q in o min in VO VO r-
COcr\
M«tJ•4JQ)
H'•H
2<4-1
%+J
&<

43
Sparks, 1954), acetaldehyde and ketones (Joslyn and Braverman,
1954) to form hydroxysulphonates, glucose and maltose (Ingram
and Vas, 1950 a,b) to form addition complexes or amines (Joslyn
and Braverman, 1954) to produce amine-bisulphites can occur.
As such compounds show little antimicrobial activity (Neuberg,
1929; Rehm, 1964), the present study was concerned with the
development of an analytical procedure which would estimate the2 -concentration of unbound (free) HSO^ /SOy referred to as 'free
sulphite' for convenience.
Lafontaine and his co-workers (1955) were perhaps the first
to note that the addition of sulphite to minced meat resulted in
a failure of the resident microflora to grow. This situation
was dependent not only on the concentration of sulphite but also
on the temperature of storage and the initial concentration of
micro-organisms. Thus a concentration of 300 yg/g total sulphite
(as SOg) caused bacteriostasis at 4°Ç but not at 20°C whereas
higher concentrations (1000 and 3000 ug/ml) were effective at
both temperatures. It is notable that these workers observed
that the addition of sulphite favoured the growth of anaerobic
micro-organisms and that the efficacy of preservation was in
versely related to the initial size of the microbial population.
Furthermore broth studies using selected isolates from minced
meat corroborated the above observations. Krol and Moerman
(1959/60) confirmed the findings of Lafontaine et al. (1955)
and reported also that microbial growth, as indicated by a
total viable count, was restricted for up to 6 days at refrigeration
temperatures in sulphited (300 yg/g) minced meat balls and a

44.
concentration of 900 yg/g resulted in the death of micro-organisms.
Even at the former concentration of sulphite, the Enterobacteriaceae
were particularly sensitive with upwards of a 90% kill during
refrigerated storage for 6 days. Fournaud et al. (1971) also
observed that sulphite was a most effective preservative against
the Enterobacteriaceae and microbial contaminants in general in
the minced pork ingredients of saucissons. Dyett and Shelley
(1962) commented on the inhibition of growth of the Entero
bacteriaceae - Salmonella spp. in particular - in sulphited pork
and beef (Dyett and Shelley, 1966) sausages. Three observations
in the latter report are of especial importance: firstly that
with incubation at temperatures below 22^C, sulphite was most
potent against the 'coli-aerogenes' group and other Gram-negative
bacteria in general; secondly, that addition of sulphite
resulted initially in a kill of the numerically dominant micro
organisms, and thirdly that it retarded the growth rate of the
microbial association thereafter. According to Christian (1963)
sulphite, at a concentration of 3.5 grains/lb was able to extend
the shelf life of minced beef 2 - 3 fold with storage at 41°F.
In accord with the above observations of Dyett and Shelley (1962,
1966) and Krol and Moerman (1959/60), Christian noted that
addition of sulphite increased the "lag" period and suppressed
the rate of growth of the dominant micro-organisms at 34° and 41°F.
Furthermore there was some evidence that the preservative exerted
a selective action on the microbial association: with storage of
unsulphited minced beef at low temperatures. Gram-negative rods
predominated whereas in the presence of sulphite short Gram-
positive rods were numerically dominant. In the former case a

45.
"putrid" spoilage was evident when microbial numbers had reached
ca, 100 X 10^ c.f.u./g whereas "souring" - a feature of sulphited
mince - could not be demonstrated until a viable cell concentration
> 500 X 10^ c.f.u./g had been achieved. Gardner (1968) could
not reproduce all of the findings of Christian (1963) with respect
to sulphite preservation of vacuum-packed baconburgers. The former
worker found that the composition of the normal microflora was not
influenced by sulphite concentration. It should be recognised
that before the addition of sulphite the product contained nitrite
and nitrate which were likely to affect the composition and behaviour
of the microbial association as well as negating the antimicrobial
effect of sulphite (Tompkin et al., 1980). With storage at either
5 , 10° or 22°C, however, sulphite exerted initially a lethal
effect on the dominant micro-organisms and retarded the rate of
their growth thereafter (Gardner, 1968). Thus with addition
of sulphite, extensions to the shelf life of baconburgers were
of the order of 2d, 10 d and 28 d at 22°C, 10°C and 5°C respectively.
In the light of the present study it is noteworthy also that,
regardless of the temperature of storage, sulphite prevented an
acid drift in the baconburgers over the period of storage.
An investigation of the influence of sodium sulphite on
Escherichia coli , Salmonella typhimurium and micro-organisms indi
genous to raw minced meat (Moerman et ai., 1966) revealed that,
with incubation at 15°c, a concentration of 0.03% sulphite caused
bacteriostasis of these organisms but it was less effective
against the normal microflora. With incubation at 20° or 25°C
the addition of the preservative had a negligible effect on the rate

46
or extent of growth of any of the micro-organisms sought. Moerman
and his co-workers (1966) were able to demonstrate a significant
decrease in the concentrations of total sulphite over the storage
period. Thus with incubation for 20 h at 37°C, only 110 yg/g of
the original 300 yg/g sulphite could be recovered whereas after
41 h at 25°C, 170 yg/g remained. The utility of sulphite in
the preservation of refrigerated raw mince was confirmed also
by Mglder (1969) who showed that with storage at 1°C, addition
of sulphite prevented microbial growth for upwards of 7 days.
Following an extensive survey (Dowdell and Board,1968) of
British fresh sausages offered for sale at retail outlets,
Dowdell and Board (1971) were able to define the composition of
the "microbial association" (pp. 16 - 19) of the product.
They observed also that the yeasts and Microbacterium thermosphapta
(Brochothrix thermosphacturn) - dominant members of this microbial
association - were capable of profuse growth in sulphited culture
media whereas pseudomonads and coliforms isolated from fresh
sausage were not. The elective nature of sulphite for a particular
type of microbial association in fresh sausage meat was first
proposed by Hurst (1972) and confirmed by Brown (1977). A
comparison of various systems of preservation for British fresh
sausage manufactured on a small scale (Ashworth et al., 1974)
confirmed the observations of Dowdell and Board (1968, 1971) that
Microbacterium thermosphactum (Brochothrix thermosphacta), lacto-
bacilli, micrococci and yeasts were the dominant microbial groups
in the sulphited product. Ashworth and his co-workers (1974)
demonstrated that although sulphite curtailed the proliferation of

47
coliforms^/'ficrobacterium thermosphactum and pseudomonads, the
lactobacilli, micrococci and yeasts were unaffected.
Furthermore they noted that a combination of polyphosphate
and sulphite acted synergistically to reduce the rate of growth
of the sensitive microbial groups. Indeed the addition of
polyphosphate enhanced the antimicrobial action of sulphite
against lactobacilli, pseudomonads and yeasts in the sausage
meat studied by Tyson (1976). She concluded that sulphite
alone delayed the onset of spoilage of fresh sausage principally
by retarding the rate of growth of the lactobacilli. The preservative
enhanced the rate and extent of growth of the yeasts, whilst only
permitting growth of the pseudomonads when the concentration of
the active (free sulphite) moiety had fallen below a critical
level.
The role of sulphite in sausage preservation was studied in
more detail by Brown (1977) who demonstrated that sulphite
inhibited the growth of coliforms at chill temperatures only.
He was of the opinion that sulphite "steered" the microbial
association towards a "fermentation" dominated by Microbacterium
thermosphactum and lactobacilli. He proposed also that free
sulphite was bound by unidentified compounds produced by the
developing microflora as well as by meat and rusk components.
The above review of the limited literature concerned with
the nature of sulphite-preservation of neutral pH meat products
has revealed several features. It has shown firstly, that the

48,
preservative is effective against micro-organisms only when present
in the free (unbound) state. Secondly, that sulphite is most
efficient at low temperatures and its potential to inhibit
microbial growth decreases with time. Thirdly, that sulphite
appears to elect for a Gram-positive/yeast-dominated microflora,
which invariably results in spoilage due to "souring", perhaps
through selectively inhibiting Gram-negative micro-organisms -
particularly the coliforms and Enterobacteriaceae. It is against
this background of meat microbiology that the wider scope of
literature concerned with the modus operand! of the salts of
sulphurous acid against micro-organisms is now set.
Factors influencing the antimicrobial action of SO^ and sulphite
Above all else it is evident that the efficacy of the salts
of sulphurous acid depends on the degree of ionization of the
molecule (Douglas, 1966; Hammond and Carr, 1976), and the
literature tends to support the view that the antimicrobial2 -efficacy decreases in the order: SO^ (H^SO^) > HSO^ > SO^
(Rehm and Wittman, 1962, 1963). Thus before conclusions regarding
the sensitivity of particular micro-organisms to sulphite can be
made, it is important that the pH of the test system is known.
For example, the inhibition by sulphite of photosynthesis and
respiration in unicellular green and nitrogen fixation in blue-
green algae (Babich and Stotzky, 1974) has been shown to be
strongly pH dependent. This phenomenon was also a feature of
sulphite toxicity to fungi and coliphage (Babich and Stotzky,
1978) although broth studies indicated that a concentration of

49
-2 2 - -5 X 10 M SQ^ (as SO^ or HSO^ ) at pH 7.0 would nevertheless
still inhibit Escherichia coli. Pseudomonas aeruginosa. Bacillus
cereus and Serratia marcescens, Vas and Ingram (1949) reported
that the rate of multiplication of yeasts was directly proportional
to the concentration of SO^ (range, O - 560 mg/2,) at pH 3.4.
The rate was retarded at pH 2.88. Rehm and Wittman (1962)
quoted minimum inhibitory concentrations of SO^ over the pH range
2.5 - 5.0 for other micro-organisms, (yg/ml in parentheses):
Saccharomyces cerevisiae (80-160), Saccharomyces ellipsoïdes
(20-80) ,Ilansenula anomola (240) , Mucor spp. (30-60) and
Pénicillium spp., (20-400).
Like most undissociated antiseptic acid molecules, SOg
probably penetrates the microbial cell more rapidly than ionic
species (Ingram et al., 1956; Rahn and Conn, 1944; Oka, 1964).
Some workers (e.g. Rahn and Conn, 1944; Maoris and Markakis, 1974;
King et al., 1981) claim that undissociated HgSOg is the only
antimycotic moiety and others (Faparusi, 1969; Okafor, 1975;
Hammond and Carr, 1976) state that at commercial concentrations
even SO^ is unsatisfactory for the preservation of products (e.g.
palm wine) which have a pH reaction > 4.0.
The efficacy of sulphite is also influenced drastically by
the presence and activity of binding agents (Hammond and Carr,
1976). Binding of sulphite has been demonstrated by many workers
(e.g. Farnsteiner, 1904; Prater et al., 1944; Ponting and Johnson,
1945; Ingram, 1948; Gehman and Osman, 1954; Richardson, 1970) and
a list of compounds in sausage which are likely to bind SO^/HSO^ / 2 -SO^ has been compiled by Brown (1977). Of primary importance

50,
are carbonyls e.g. aldehydes, ketones and sugars (Luck, 1977);
aldehydes forming hydroxysulphonates, amines forming amine-
bisulphites (Joslyn and Braverman, 1954) and sugars forming
addition compounds (Ingram and Vas, 1950 a,b). An acidic pH
(3 - 5) favours the formation of such complexes (Rehm and Wittman,
1962, 1963) although temperature and concentration of reactants
also influence the rate and extent of binding (Burroughs and Sparks,
1964). Binding compounds may be present in food (Ingram and Vas,
1950 a,b), formed during storage (Burroughs and Sparks, 1964)
or produced by moulds and bacteria (Burroughs and Sparks, 1964;
Weeks, 1969; Rankine and Pocock, 1969).
It is generally accepted that only free (unbound) sulphite
is antimicrobial (Neuberg, 1929; Ingram, 1948; Rehm, 1964).
Thus the claim (Fornachon, 1963 ; Lafonlafourcade and Peynaud,
1974), that bound sulphite killed micro-organisms is probably
illfounded and due to dissociation of the complexed sulphite
producing small concentrations of free preservative.
The salts of sulphurous acid are reported to be more effective
against moulds than yeasts (Robson, 1968; Roberts and McWeeny,
1972; Baird-Parker, 1980). Of the yeasts, those with an oxidative
metabolism (e.g. Pichia membranaefaciens, Debaryomyces spp. and
Rhodotorula spp.) are considered (Rehm and Wittman, 1962) to be
less resistant than those capable of fermentation (e.g. Hansenula
spp., Saccharomyces spp.) and Reed and Peppier (1973) noted that
Pichia membranefaciens and Kloekera apiculata were 'SOg-sensitive'
whereas Saccharomycodes lugwidii, Saccharomyces baileii and

51
Brettanomyces spp. would resist up to 500 yg/ml SO^. It has been
suggested that differential uptake of, and affinity for SO^ is
the reason for the apparent resistance of some yeasts, (Hammond
and Carr, 1976) although uptake is now believed to be by passive
diffusion (Stratford, 1983) and not by active transport (Maoris
and Markakis, 1974). Adaptation of yeasts to SO^ tolerance has
been reported also (Schanderl, 1959; Robson, 1968).
Bacteria are among the most sensitive micro-organisms to
sulphite. Rehm and Wittman (1962) noted the following minimum
inhibitory concentrations at pH 6.0 (ug/ml 80^ in parentheses):
Pseudomonas fluorescens (50), Escherichia coli (100-200),
Staphylococcus aureus (80), Bacillus spp. (50) and Lactobacillus
casei (100). It has been suggested also (Roberts and McWeeny,
1972) that Gram-negative are in general, more sensitive than
Gram-positive bacteria.
Mechanism of action of sulphite
Several potential targets of microbial metabolism may be per
turbed by sulphite. Most workers (e.g. Freese et al., 1973 ; Hammond
and Carr, 1976; Luck, 1977)are of the opinion that sulphite
inhibition results from multisite disruption of metabolism.
It has been concluded by some (Maoris, 1972; Maoris and
Markakis, 1974) but not all (Stratford, 1983) workers that SO^
is actively transported into the yeast cell. The former workers

52,
were able to demonstrate saturation kinetics with increasing SO^
concentration whereas Stratford (1983) contends that transport
is by diffusion only. Indeed he associated the low rate of
diffusion of SO^ into the cells of Saccharpmycodes ludwigii
with the organism's tolerance of 10 times the concentration of
SO^ which would inhibit Saccharomyces cerevisiae for example.
Low (ca. 1 yg/g) concentrations of molecular SO^ can inhibit,
and higher concentrations, kill yeasts (Schimz, 1980), and
stationary-phase cells are more sensitive than those in active
growth (Schimz, 1980). Following exposure to sulphite, most
micro-organisms exhibit a lag period before cell viability
decreases rapidly. Sulphite damage in yeasts is usually ir
reversible after ca. 60 min exposure (Schimz and Holzer, 1979)
and A.T.P. is released into the medium. A possible reason for
the efflux of A.T.P. from yeast has been suggested by Warth
(pers. comm.). He claims that sulphite lowers the internal pH2 -of the cell. Thus when SO^ enters the cell as an uncharged
molecule, it becomes ionized due to the higher pH of the
interior of the yeast cell and releases protons which lower the
pH. As this would result in the disappearance of the membrane
proton gradient, the yeast would expel A.T.P. in an attempt to
release protons and re-establish the gradient.
It has long been known that the addition of sulphite to growing
cells of Saccharomyces cerevisiae caused an increase in the con
centration of glycerol in the medium (Neuberg and Reinfurth, 1918,
1919); small doses of sulphite added at intervals prevent
cytotoxicity and merely "steer " the fermentation (Freeman and

53
Donald, 1957). There has been speculation that the glycerol
formed is from glyceraldehyde-3-phosphate as a result of blockage
of glycolysis (Gancedo et al., 1968). Indeed, Stratford (1983)
found that in aerobically-grown cells of Saccharomyces cerevisiae
the rate of decarboxylation by pyruvate decarboxylase was maintained2 -in the presence of sulphite despite binding of SO^ to acetaldehyde.
It was probable therefore, that NAD^ was regenerated using oxygen as
the terminal electron acceptor. Sulphite may then prevent A.T.P.
generation from glycolysis or increase the rate of A.T.P. utilization,
In the first instance, sulphite would act as an uncoupler of
glycolysis thus facilitating intermediate flow to be maintained.
whilst preventing concomitant ATP generation. In anaerobically-2-grown cells of this organism, Stratford (1983) noted that SO^
bound intracellular acetaldehyde thereby preventing the regeneration
of oxidised NAD^ by ethanol dehydrogenase. Shortage of NAD^
would prevent activity of glyceraldehyde-3-phosphate dehydrogenase
and therefore prevent flow of intermediates via glycolysis and
further decarboxylation of pyruvate. Accumulation of glyceralde-
hyde-3-phosphate would promote the formation of glycerol and re
generate some NAD . No net energy (as ATP) however, could be
formed by this means.
Such observations are in accord with those of others (e.g.
Pfleiderer et al., 1956) who showed that sulphite was active against
-SH groups and was a powerful inhibitor of NAD-dependent reactions.
Indeed, SO^ can react directly with nicotinamide adenine dinucleotide
(NAD) thus preventing the occurrence of energy-yielding oxidative
reactions (Meyerhof et ai., 1938; Dupuy, 1959). Rehm (1964)

54
indicated that at least three steps in glycolysis in Escherichia
coli and Saccharomyces cerevisiae were strongly inhibited by
sulphite. The blocking of the glyceraldehyde-3-phosphate ^ 1,3-
diphosphoglycerate step is probably the most crucial in yeasts
whereas the NAD-dependent formation of oxaloacetate from malate
is more important in Escherichia coli (Wallndfer and Rehm, 1965).
Several other reactions involving sulphite have been mentioned
in the literature. For instance sulphite may inhibit growth by
reacting with products of intermediate metabolism thus shifting
the reaction equilibria to thermodynamically-inefficient states
(Rehm and Wittman, 1952), or reversibly inhibiting g-galactosidase
induction, RNA and protein synthesis in Escherichia coli. (Robakis,
1980). Sulphite may also break disulphide bridges by replacing
the sulphur group from the bond of cystine or cystine peptides
(Clarke, 1932) and as sulphonation of disulphide bridges in enzymes
can lead to loss of enzyme function it is a likely site of sulphite
toxicity. Some of the reactions of bisulphite and sulphite with
important cell constituents are outlined below;
Reaction of HSO^ with 4-thiouridine
4-Thiouridine occurs in many bacterial tRNA's between the
dihydrouridine stem and the CCA stem (Nishiraura, 197 2) and HSO^
reacts via an O^-mediated free radical mechanism:

55
S — SO-
s
HN
R
R
so
SQ-
R

Addition of HSO to uracil
56
0
R
0
HN HSO. HN
R
Deamination of cytosine by HSO,
NH.
H N
R
0
R
NH.
H NHSO
SO
R
H2ONH.
0
HN
N SON
R

57.
Reaction with carbonyl compounds
\ \C ^ O HSO: y
R R'
Finally it has been demonstrated also (Tanaka and Luker, 1978) that2_killing of bacteria and yeasts by SO^ (up to 24 mM) was more
effective under aerobic than anaerobic conditions. They found that2 -when Escherichia coli was exposed to SO^ , leakage of g-methyl-
glucoside occurred, followed by rapid loss of ATP. Furthermore2 -they noted that SO^ (15.6 mM at pH 6.0) killed Salmonella
typhimurium and Bacillus subtilis by breaking DNA as indicated by
a more rapid kill of DNA-repair mutants than their prototrophs.
2 -In summary, it is evident that SO^/HSQ^ or SO^ can act
in several ways to inhibit or kill micro-organisms.
Reaction with disulphide bonds (sulphitolysis)
R-S-S-R ^HSO~^ RSSO^ + R-SH
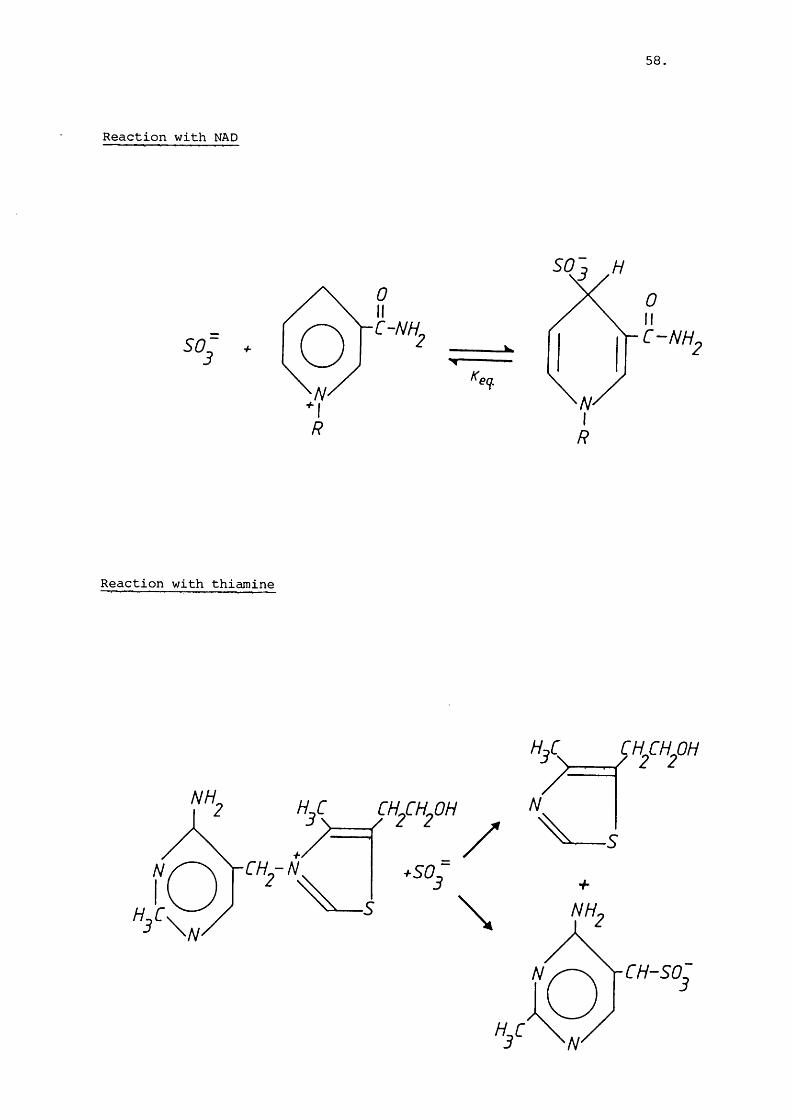
Reaction with NAD
58.
so ~ +C-NH,
R
K .eq.
C-NH.
R
Reaction with thiamine
CH.-N
H X CHSH^OHJ \ / 2 2
«/N
/ V■so
■S
3
\cH -so:

59
As the reaction of the preservative is influenced greatly by the
nature of the environment, e.g. pO , pCO , a , pH, nutrient status,A 2 wavailability of binding compounds, temperature and types,numbers
and physiological status of the test micro-organisms, the action
of sulphite in such a complex system as the British fresh sausage
will be difficult to resolve. It was for this reason that the
present study concentrated on the action of sulphite on microbial
growth, measured by changes in opacity or colony-forming units.

60
MATERIALS AND METHODS
Sausage manufacture
Commercial scale
Sausage ingredients were weighed into a bowl chopper (Laska,
Austria) in the proportions of a commercial sausage recipe
(Appendix, p.329 )• The total capacity of the mix was 500 Z.
When appropriate, the first batch was made without preservative,
the second included preservative (sodium metabisulphite) in the
seasoning (Rank-Hovis-McDougall 599,England), at the "commercial
level" which was predicted to give a final concentration of ca.-1600 yg SO2 9 sausage. The meat/rusk slurry was extruded
mechanically into reconstituted collagen casings (Devro, England),
linked by hand, packed in eights and wrapped in food grade
cellophane film (25 ym thickness). The oxygen permeability of
the latter at 25* 0 was 400 cc/m^/h/0.025 mm; water vapour
permeability at 25°C and 75% relative humidity was 5 g/m^/24 h/
0.025 mm.
Laboratory scale
Sausage ingredients obtained from material awaiting use in
sausage manufacture in a factory of a large-scale manufacturer
were minced (Kenwood A901), and weighed into a clean stainless
steel bowl in the proportions of a commercial sausage recipe
(Appendix p. 329). The total capacity of the mix was 2 Kg. The
ingredients were chopped vigorously (Kenwood A901). Additives:
sulphited or unsulphited seasonings at the commercial level.

61.
potassium sorbate (2,000 yg/g) or solid carbon dioxide pellets
(20% w/v) were chopped into the slurry in various combinations
(Appendix, p. 309 ). The sausage meat was extruded through a
sausage stuffer (Kenwood) into reconstituted collagen casings
(Devro, England), linked by hand, packed in eights and wrapped
in film (vide supra).
Seeding of sausage meat
Rifampicin-resistant Salmonella were inoculated into Brain
Heart Infusion broth (BHIB, pH 7.0; Difco) and incubated at 37°C
for 18 h. Freshly manufactured sulphited or unsulphited sausage
meat obtained from the factory was inoculated with Salmonella
at a level calculated to give a final concentration of ca. 1 x 10^— %
Salmonella g sausage. The inoculated meat slurries were mixed
thoroughly (Kenwood A901) and samples (100 g) put into square
Petri dishes (Sterilin). Uninoculated samples were prepared
also, sterile distilled water being added to the sausage meat instead
of a suspension of Salmonella.
Storage of sausages and sausage meat in Petri dishes
As with factory practice, all sausages and sausage meat were
stored aerobically at 4* C for 24 h. Thereafter packs of sausages
were kept at 4°, 10°, 15° or 22°C for up to 14 d. Petri dishes
containing seeded sausage meat were kept at -20°, 4°, 9°, 15°,
20° or 25°C for up to 8 d.

62
Sampling of sausages and sausage ingredients
Samples of pork sausage, pork and beef sausage and all the
ingredients used in their manufacture were obtained from a large
factory during normal production in the period 1.10.79 to
31.9.82. One Kg of pork meats were taken at random from material
awaiting processing. The meat came from pigs slaughtered at the
factory; butchered meat had been stored at 4°C overnight.
Cattle were neither slaughtered nor butchered on site and 1 kg
samples were taken from blocks of frozen, deboned beef. Samples
of rusks, seasoning, polyphosphate (Pibrisol VIO), pasteurized
and dried rinds, mechanically recovered meat and linked sausages
were examined also. All samples were stored at 4°C and examined
within 3 h. To minimise sampling errors (Elliott, 1977), all
meats were comminuted in a sterile mincer (Kenwood). Ten sub
samples (each of ca. 2 g) were taken at random from a minced
ingredient, bulked, and homogenised in 180 ml of ^ strength
Ringers diluent (Qxoid BR52) with a Colworth stomacher 400
(Seward, London) for 50 s. Further serial dilutions were done
in Ringers diluent (9 ml). With linked sausage four sub
samples were taken from either the surface (depth up to 0.25 cm)
or "core" sites (innermost 1 cm diameter cylinder) or from a
transverse section, and, as with meat ingredients, a 20 g
sample was used.
Determination of sulphur dioxide (SO^) in sausage
Monier-Williams (1927) method, modified by Shipton (1959)
Specimen preparation. A sample (20 g) of sausage was taken from a

53
standard pack (454 g) using the sampling scheme described on p.62
Total SO^ determination. Analysis was done according to the
modification of Shipton (1959). The specimen was acidified
with orthophosphoric acid (2M, 50 ml) and distilled for 50 min.
Hydrogen peroxide was used to collect the SO^ (Burroughs and
Sparks, 1953).
Electrometric ion-selective probe method (Bailey and Riley, 1975;
Brown, 197 7).
Specimen preparation. A sample (5 g) of sausage was taken from a
standard pack, added to distilled water (45 ml) and homogenised
in a Colworth Stomacher 400 (Seward, London) for 60 s. The
homogenate was centrifuged (MSE Super Speed 18) at 35,000 g for
10 min. When the set speed was achieved, the compressor was
regulated to cool at 2°c. Chilled centrifugation separated a
fatty layer from the aqueous layer and a solid base, comprising
of starch, meat and rind particles (Brown, 1977). The aqueous
layer was decanted and stored in a stoppered volumetric flask
(50 ml) in an ice bath to reduce SO^ loss before analysis.
Total SO determination . One part of sodium hydroxide (IM) was
added to one part of specimen solution and stored at 25°C for
30 min to dissociate bisulphite-addition compounds. A sample
of alkalised specimen (25 ml) was placed in an Erlenmeyer flask
(100 ml) containing a magnetic stirring bar and an equal volume

64
of sulphuric acid (1 M) added. The flask was put in an ice bath
on a stirrer bed (Chem Lab SS3H) and the probe (EIL, Chertsey)
immersed to a depth of 1.5 cm. The peak deflection (mV) after
2 min as registered on an EIL 7030 pH meter (Monovalent cation
setting) or as indicated on a flat-Joed pen recorder (Servoscribe)
noted.
Free SQ determination. Analysis was done immediately after specimen
preparation vide supra. No alkalisation procedure was used and
acidification with sulphuric acid released the free SO^ from the
specimen which was then estimated as for total SO^ determination
vide supra.
Spectrophotometric method
Specimen preparation. A sample (5 g) of sausage was taken from
a standard pack, added to chilled distilled water (25 ml) in a
screw-capped bottle (100 ml) which contained glass beads, and
shaken 60 times through an arc of 0.5 m for 30 s. The homogenate
was stored in an ice bath before analysis.
The specimen was placed in a three-necked round-bottom flask
(Q&Q FR 250/35/22A). One side neck was connected to a supply of
nitrogen (Air Products, Bristol) via a modified sparger (Q&Q
MF 27/3/13). A vertical condenser (Q&Q Cl/13) was connected to the
centre neck and two Drechsel bottle heads (Q&Q 27/3/13) were
attached in parallel to the condenser outlet by means of a "y"

65
junction adaptor (Q&Q MF 10 2B) with taps. The dip tubes of the
Drechsel bottle heads were placed in boiling tubes (Q&Q MF 24/3)
below the surface of solutions of 5,5'-dithiobis 2-nitrobenzoic-3acid (DTNB) (50 ml). DTNB reagent (final concentration 2.5 x 10 M;
-2pH 8.0) was dissolved in phosphate buffer (1.6 x 10 M Na^HPO^ +-31.8 X 10 M KH^PO^) containing ethanol (10% v/v). Nitrogen was
3 -1then passed through the distillation flask (20 cm s ). Hydro
chloric acid (2 M, 50 ml) was added via the remaining side neck
and the flask was heated. On the completion of distillation
(15 min) DTNB was added to the phosphate buffer (50 ml) and the
absorbance measured at 412 nm on a Pye Unicam SP6-550 UV/VIS
spectrophotometer (Wedzicha and Bindra, 1980).
Free 50^ determination. Analysis was done immediately after
specimen preparation by transfer of the specimen to a distillation
flask (Q&Q 250/35/22A) containing a magnetic stirring bar. A
condenser was not used and the tubing adaptor (Q&Q MF 10 2B) was
attached directly to the flask. No heat was applied and the
reaction was allowed to continue for 15 min after addition of
hydrochloric acid (2M, 50 ml). Nitrogen was passed through the3 -1distillation flask (20 cm s ) as above. Analysis of DTNB was done
as for total SO^ determination.
Standard sulphite solutions
Sulphite solutions were made up from potassium metabisulphite-4and contained ethylene-diamine-tetraacetic acid (EDTA, 1 x 10 M)
to prevent oxidation. The solutions were standardised iodimetrically

65,
by volumetric analysis.
Standard curves of sulphite concentration
Electrometric ion-selective probe analysis. Standard solutions of
sulphite were treated in the same way as specimen solutions to
give a standard curve (Fig. 2).
Spectrophotometric analysis. Standard solutions of sulphite were
added directly to buffered DTNB reagent and analysed spectrophoto-
metrically in the same way as specimen solutions to give a standard
curve (Fig. 3). A freshly prepared reagent blank was used for each
set of readings.
Determination of sulphite concentration of sausage seasonings
1. A sample (0.01 g) of seasonings was added to DTNB (25 ml),
diluted with buffer to a final volume of 50 ml and analysed
spectrophotometrically.
2. A sample (0.01 g) of seasonings was used. The concentration
of free and total SO^ was determined by the method described.
3. lodimetric method (used by one large manufacturer of sausage).
A sample (1 g) of seasonings was added to distilled water
(25 ml) in an Erlenmeyer flask (250 ml). Two ml of
sulphuric acid (10% w/v) and 3 drops of soluble starch were
added and mixed thoroughly. This solution was titrated with
iodine (0.1 N). The percentage SO^ = titration value (ml) x 0.32%

67.
Determination of sulphite-binding capacity of sausage ingredients
Sterile lean pork was excised from large (300 x 200 mm) blocks
of meat, minced aseptically (Kenwood A901) and samples (25 g)
added to sterile Erlenmeyer flasks (250 ml). Samples (25 g) of
minced lean, head and belly pork, rinds, fat or mechanically
recovered meat (MRM) were added to flasks, as was rusk also. Solutions
of filter-sterilised potassium metabisulphite (100 ml, ca. 600 yg
SO^/ml) were added aseptically to the flasks which were stoppered
with rubber bungs. The flasks were incubated at 25°C in a shaking
(88 reciprocal excursions over 5 cm) water bath for up to 6 h.
Uninoculated sulphite solutions were used as controls. Concentrations
of free and total sulphite of the slurries were determined by the
method described on pp. 64 - 66.
Sulphite-binding agents
Preparation of detection solution. An aqueous solution (pH 8.0) of
basic fuchsin (0.02 w/v) and sodium metabisulphite (0.125% w/v)
was made in deoxygenated distilled water and stored under a
blanket of scrubbed (Appendix, p.335 ) dinitrogen at ca. 4°C.
Preparation of standard curve. Samples (1 ml) of various concen
trations of acetaldehyde were added to the detection solution (10 ml),
sparged with scrubbed dinitrogen and the increase in absorbance
(A = 538 nra) measured immediately against a blank control (Pye
Unicam SP UV-VIS spectrophotometer). The presence of sulphite-
binding agents in unsulphited culture fluid was determined as
above, and the concentrations, estimated by reference to the

68
standard curve (Fig. 4). In examination of sulphited culture
fluid, correction was made for the presence of "endogenous" sulphite,
2 -Sulphate (SO ) determination
A modified method based on that of Bertolacini and Barney
(1957) was used. Deionised water was used. Micro-organisms were
removed from culture fluid by centrifugation (7,000 g; 15 min)
and a sample (1 ml) of the supernatant was added to 1 ml of
0.12 M potassium hydrogen phthalate (COCH.C_H .COOK) in ethanol6 4(50% v/v). Five rag of solid barium chloranilate (BaC^Cl O.)6 2 4was added, the mixture "whirlimixed", centrifuged (7,000 g;
15 min) and the absorbance of the supernatant (chloranilic acid)
measured (A = 332 nm) on a Pye Unicam SP 6 UV-VIS spectrophotometer.
A standard curve, using various concentrations of sodium sulphate
(Na^SO^.I2 H2 O) was made using the same method (Fig. 5). Where
there was a possibility of interference from cations or components
which would absorb near 332 nm, a sample (30 ml) of culture super
natant was passed down a column of Dowex 50X8 resin (10 ml bed
volume in the form) at ca. 5 ml/min. The effluent was brought
to pH 4 with ammonium hydroxide (NH^OH), adjusted to volume and
a sample (4 ml), mixed with 0.05 M potassium hydrogen phthalate
(1 ml). After vigorous agitation and centrifugation (7,000 g,
15 min) the absorbance of the supernatant was measured at 530 nm
against a deionised water blank.
Concentrated solutions of freshly-prepared sulphite (2.5 -
12.5 mM) also gave sane colour in this assay so a calibration curve

69
was prepared (Fig. 6) with sodium sulphite (Na^SO^.TH^O). From
this curve, absorbance corrections at 530 nm due to sulphite
(determined independently with DTNB reagent) could be made in2 - 2 -samples containing both SO^ and SO^ moieties.
Removal of thiol compounds from samples
Interference from thiol compounds was removed in two ways
(1) formaldehyde (O.Ol ml; 50 mM) was added to the sample (1 ml)
which contained both sulphite and a thiol. The sulphite was
complexed as formaldehyde sulphoxylate so that it did not react with
the DTNB reagent. Residual absorbance at 412 nm was due to thiols
and the appropriate correction made (Fig. 7). (2). The sample
(1 ml) was diluted 10 fold and allowed to stand in air at 25°C
for 20 min. This completely oxidised thiols to the corresponding
disulphides after which gave no colour with DTNB (Fig. 8). Residual
colour was due to sulphite.
pHA sample (20 g) of sausage or an ingredient of sausage was
homogenised in distilled water (pH 7.0; 180 ml) in a Colworth
Stomacher 400 (Seward, London) for 60 s. The pH of the
homogenate was measured with a glass pH electrode (Russell,
England).

70.
Isolation, enumeration and maintenance of micro-organisms from meat
and sausage
Total viable count (TVC): From appropriate dilutions, samples
(20 yl) were pipetted onto the dried surface (37°C, 60 min) of
PCA. Using the technique of Miles and Misra (1938), 32 replicate
drops were accommodated in one Petri dish (diameter, 9 cm). Incubation
was at 20^0 for 3 d, and colonies picked at random were purified
by streaking on PCA. Pure cultures were stored on PCA slopes at
4°c with sub-culture at 4 week intervals.
Pseudomonas
Samples (0.2 ml) of an appropriate dilution were spread over
the dried (37^c, 60 min) surface of Cetrimide-Fusidic acid-
Cephalodrine (CFC, Mead and Adams, 1977) medium. Incubation
was at 15°C for 48 h and 3 replicates at each dilution level
were done. Colonies picked at random from the lowest countable
dilution of CFC agar were purified by streaking on PCA and
maintained thereafter on PCA slopes at 4°C with subculture every
4 weeks.
Lactobacillus-
A sample (1 ml) of an appropriate dilution was added to 10 ml
of MRS (de Man et al., I960) medium at 50°C. When set, an overlay
(10 ml) of MRS was added. Three replicates were done at each
dilution level and incubation was at 25°C for 5 d under anaerobic
conditions. Colonies picked at random from the lowest countable

71.
dilution of MRS agar were purified by streaking on Heart Infusion
(HIA; Difco B44) agar, and maintained in Robertson's cooked meat
medium (Appendix p . 333) at 4°C. The isolates were subcultured every
4 weeks.
Group D streptococci
Kanamycin aesculin azide agar (KAA; Kanamycin aesculin azide
broth + 1.5% w/v agar) was sterilised by autoclaving at 121°C for
15 min. A sample (1 ml) of an appropriate dilution was mixed with
KAA (20 ml) at 50^0. Three replicates were done at each dilution
level and incubation was at 37°C for 24 h. Colonies picked at
random from the lowest countable dilution of KAA were purified
by streaking onto Todd Hewitt agar (TDA, Appendix p. 333 ), and
maintained in Robertson's cooked meat medium (Appendix, p. 333)
at 4°C with subculture every 4 weeks. ,
Brochothrix thermosphacta
A sample (0.1 ml) of an appropriate dilution was spread over
the dried surface (37°c, 50 min) pf streptomycin and thallous
acetate actidione (STAA, Gardner, 1966) medium. Three replicates
were done at each dilution level and incubaticn was at 20°C
for 3 d. Colonies picked at random from the lowest countable
dilution of STAA were purified by streaking on PCA and maintained
on PCA slopes at 4°c with subculture every 6 weeks.
Yeasts
A sample (O. 1 ml) of an appropriate dilution was spread over

72
the dried (37°C, 60 min) surface of Rose Bengal chloramphenicol
(RBC, Oxoid CM 549) medium. Three replicates were done at each
dilution level and incubation was at 25°C for 3 d in the dark.
Colonies picked at random from the lowest countable dilution on
RBC were purified by streaking onto malt extract (MA, Appendix
p. 333) agar. Isolates were maintained on MA slopes at 4°C and
subcultured every 6 weeks.
Enterobacter iaceae
Lactose fermenting Enterobacteriaceae. A sample (1 ml) of an
appropriate dilution was mixed with 10 ml of violet red bile
(VRB, Oxoid CM 107) agar which had been cooled to 46°C. When set,
the surface of the agar was overlaid with VRB (10 ml). Incubation
was at 37°C for 24 h.
Glucose fermenting Enterobacteriaceae. A sample (1 ml) of an
appropriate dilution was mixed with 10 ml of violet red bile glucose
(VRBG Oxoid CM 485) agar cooled to 46^C. When set, the surface of
the agar was overlaid with VRBG (10 ml). Incubation was at 30°C
for 24 h. Six replicates at each dilution level were done and
colonies selected at random were picked from VRB or VRBG and purified
by streaking on PCA. Isolates were maintained on slopes of PCA
at 4°C with subculture every 4 weeks.
Rifampicin-resistant Salmonella
Salmonella^^^ ^ were recovered on Desoxycholate citrate (DCA,

73
Difco B274) agar supplemented with rifampicin (Sigma) at a concen--1 otration of 50 yg ml medium. DCA was boiled, cooled to 50 C
and a filter-sterilised solution of rifampicin in methanol (BDH)
added and the medium (DCAR) mixed. Petri dishes were poured
immediately and the surface of the medium dried (37°C, 50 min)
before use. A sample (0.1 ml) of an appropriate dilution was
spread over the surface of the medium. Six replicates were
done at each dilution level and incubation was at 37°C for 48 h.
Isolates were maintained on PCA slopes at 4°C with subculture
every 2 weeks.
Selective enumeration of drug-resistant Salmonella
Rifampicin and Nalidixin resistant Salmonella
Drug-resistant Salmonella , Hafnia alvei
(E 58),Escherichia coli (E16), Enterobacter agglomerans (E85) and
Citrohacter freundii (E91) were tested for growth on solid and
in liquid media.
Test organisms were incubated at 30°C for 16 h in TSB after7 8 -1which time cell concentration was ca. 10 - 10 c.f.u. ml
broth in all cases. Samples (1 y 1) were multipointed (Denley)
onto the dried (37°C, 60 min) surface of DCAR, PCA + rifampicin
(50 yg/ml) PCA and MacConkey agars and into TSB or TSB + nalidixic
acid (30 yg/ml) contained in Petri dishes (Sterilin).

74.
Influence of recovery medium on selective enumeration of Salmonella^^^ ^
mutants
Rifampicin-resistant strains of Salmonella goerlitz, virchow^
kedougouf hadar, anatum, an unidentified group E4 Salmonella and
strains of Enterobacteriaceae isolated from British fresh sausage
(E 58, E 16, E 85 and E 91 vide supra) were incubated at 30°C for
15 h in TSB (10 ml). Decimal dilutions were made in sterile
distilled water (9 ml). Five replicate samples (0.1 ml) of
appropriate dilutions of single strains or mixed (in the ratio 1:1)
strains were plated onto TSA, TSA + rifampicin(50 yg/ml) or
DCAR.
Yeasts
The yeasts were characterised by the methods given below in
order to assess whether or not sulphite was having a selective
role in sausages.
Morphology
Gross. Colony appearance, pigment and texture on malt extract agar
(Malt, 6% w/v) after incubation at 25°C for 5 d was noted.
Microscopic. The size and shape of cells and occurrence -of
capsules, arthrospores, pseudo- or true hyphae was noted.
Production of urease. The method of Christensen (1946) was used

75.
Assimilation of nitrate
The method of Wickerham (1945) was used. Difco carbon base
(B391; 11.7% w/v) was used as the basal medium.
Fermentation of glucose
The method of Beech et al. (1968) with filter-sterilised glucose
(6% w/v) was used.
Assimilation of organic substrates
The basal medium (Difco yeast nitrogen base, B392; 6.7% w/v)
was filter-sterilised, dispensed into sterile universal bottles
in 25 ml volumes and maintained at 55°C. Organic substrates were
dissolved (4% w/v) in distilled water, filter-sterilised, dispensed
into sterile universal bottles in 5 ml volumes and kept at 55^C .
Purified agar (Oxoid L28; 4% w/v) was dispensed into 100 ml bottles,
sterilized by autoclaving at 115°C for 10 min and cooled to 55°C,
The basal medium and organic supplement was added aseptically to the
agar base, mixed and kept at 55^C. A sample (250 yl) of the
complete medium was dispensed into wells of sterile Microtitre
plates (Sterilin) and allowed to set.
Test strains were incubated in 10 ml of yeast nitrogen base
(6.7% w/v) containing D-glucose (0.1% w/v) at 25°C for 2 d. The
suspensions were centrifuged (7,000 g, 15 min) and the pellets
were resuspended finally in sterile distilled water (2 ml). Samples
(200 yl) were pipetted into the wells of a sterile Microtitre plate
(Sterilin).

76
Inoculation
Ninety-six map pins (Rexel Co. Ltd.) were glued (Araldite)
into the wells of a Microtitre plate. This inoculation plate was
soaked in ethanol (70% w/v) for 2 h, dried (45^C, 6 h) and used
to inoculate suspensions of washed yeast strains into the wells
in Microtitre plates containing single organic substrates.
Incubation was at 25°C for 14 d in a moist air environment.
Identification was achieved by reference to traditional
works (e.g. Lodder, 1970) or to a numerical data base (API 20C
Auxanogram).
Lactic acid bacteria
For the preliminary characterization of organisms isolated on
MRS agar (presumptive lactobacilli) or KAA (presumptive streptococci),
the following properties were used:
Growth on modified (Dowdell and Board, 1968) Keddies (1951) medium
(Appendix, p. 33]} was tested. Incubation was at 25°C for up to
14 d. The Gram reaction according to the method of Kopeloff and
Beerman (1922) was assessed. Production of catalase was.tested.
Hydrogen peroxide (10 vols.) was applied to a nutrient agar
slope culture (72 - 96 h) of the organism. Growth at 15°, 37°
or 45°C in MRS broth ( 9 ml) was assessed using 0.1 ml of an
overnight culture grown in MRS at 30°C as inoculum. Incubation
was for up to 14 d. The homo- or heterofermentative action on
glucose in the medium of Gibson and Abd-el-Malek (1945) was
determined; incubation was at 25°C for up to 14 d. Evidence of

77
oxidative or fermentative breakdown of glucose was tested with the
OF medium (Appendix p.332) of Whittenbury (1963); incubation was o
at 25 C for up to 14 d. Further characterisation of the lacto
bacilli involved the monitoring of the following properties:
The type of isomer of lactate which was formed from glucose
breakdown was determined using the methods of Barker and Sommerson
(1941) and Garvie (1967). The production of carbon dioxide from
the breakdown of gluconate and the hydrolysis of aesculin were
tested according to the methods of Collins and Lyne (1976).
The formation of ammonia from arginine was tested (Appendix
p. 332). The API 50 CHL scheme (Paule, 1971) was used to study
the fermentation of 49 carbohydrates (see below). The methods
used were those recommended by the manufacturer (API Systems,
Basingstoke); incubation was at 25°C for up to 72 h.
Organic substrates used in fermentation studies
0 Control 10 Galactose
1 Glycerol 11 Glucose
2 Erythritol 12 Fructose3 D Arabinose 13 Mannose4 L Arabinose 14 Sorbose
5 Ribose 15 Rhamnose
6 D Xylose 16 Dulcitol7 L xylose 17 Inositol
8 Adonitol 18 Mannitol
9 3 Methyl xyloside 19 Sorbitol

78-
20 a Methyl mannoside 36 Starch21 a Methyl glucoside 37 Glycogen22 N acetyl glucosamine 38 Xylitol23 Amygdalin 39 Gentiobiose24 Arbutin 40 D Turanose25 Esculin 41 D Lyxose26 Salicin 42 D Tagatose27 Cellobiose 43 D Fucose28 Maltose 44 L Fucose29 Lactose 45 D Arabitol30 Melibiose 46 L Arabitol31 Sucrose 47 Gluconate32 Trehalose 48 2 Keto gluconate33 Inulin 49 5 Keto gluconate34 Melezitose35 Raffinose
In addition to the preliminary rests used for the identification
of the lactic acid bacteria vide supra,the following tests were
used for classification of the streptococci: The ability to
reduce tétrazolium and to grow in the presence of 0.4% potassium
tellurite were tested using the methods of Facklam (1972). The
tests described below were done under aerobic or anaerobic conditions
in Repli-dishes (Sterilin ): Tolerance of 40% bile, ability to
grow at pH 9.5, 10°C or in the presence of 5.5% NaCl in MRS broth;
production of haemolysis on a modified Kneteman's (1947) medium
(Appendix p. 332 ); oxidation or fermentation of carbohydrates (see
below).

79
Carbohydrate (final concentration, 1% w/v) in basal medium
of Whittenbury (1963; Appendix, p. 332)
Azelate; Arbutin; Citrate; Raffinose; Maltose; Salicin;
D^trehalose; Mucate;Ad onitol; DvMele.zitose; D+Melibiose;
Sorbitol; Inulin; Arabinose; Cellobiose; Sarcosine: Glycerol;
Mannitol; Pimelate; Inositol; D- ribose.
Preparation of inoculum
Strains of Streptococci were incubated at 30°C for 24 h in
10 ml of Streptococcus medium (SM, Appendix, p.332). Cultures
were centifuged (7,000 g; 5 min) and the pellet resuspended in
sterile Ringers (10 ml). The washing procedure was repeated and
ca. 1 ul placed in the liquid or, on the surface of solid media
contained in Repli-dishes (Sterilin) by multipoint inoculation.
For carbohydrate utilization studies, each well of the Repli-
dish contained ca. 5 ml of Whittenbury's (1963) medium and
provided aerobic and anaerobic conditions. This was indicated
by the site of growth and by the reduction of methylene blue
below the surface layer (2 cm) of uninoculated medium. Reducing
conditions existed throughout the incubation period of 7 d at
30°C. Inoculated Repli-dishes were incubated in moist chambers
or in anaerobic jars.
Serology of Streptococcus antigens
The counterimmunoelectrophoresis (CEIP) method was used.

80.
Extract preparation
Pure cultures of the test strains were inoculated into 20 ml
of sterile Todd-Hewitt broth (Oxoid DM 189) in plastic centrifuge
tubes (capacity 25 ml). Incubation was at 37°C for ca. 18 h
after which time the tubes were centrifuged at 7,000 g for 15 min.
The pellets were resuspended in 1 ml amounts of 0.05 N HCl in
saline (0.85%), and the pH was adjusted to 2.3. The suspensionso owere heated to IQO C for 10 min in a water bath, cooled to 25 C
and brought to a neutral pH with NaOH (1 M). The suspensions
were centrifuged (7,000 g; 15 min) and the supernatants were used
as antigens in the CEIP procedure.
Gel and buffer preparation
Hydrochloric acid (0.2 N; 38.2 ml) was added to barbitone
buffer (sodium methyl-barbiturate, 8.25 g; glass distilled water,
1000 ml) to give a final pH of 8.6. Agarose (1 g , Sigma)
was dissolved in this barbitone-HCl buffer (100 ml) by heating in
a water bath, maintained at 70°C and poured onto preheated
(50°C) glass slides (300 mm x 200 mm) to a gel thickness of
ca. 2 mm. When the gel had set holes (5 mm diameter) were
punched 10 mm apart along the electrophoretic axis. Sixty-
eight wells could be accommodated on a slide. Streptococcus
antisera (30 yl; ZJ series, Wellcome, Beckenham) were
pipetted into anodal wells; antigens (30 yl) into cathodal wells,
Electrophoresis
The gel was placed in an electrophoresis tank which was filled

81
-1with barbitone-HCl buffer. The current was adjusted to 2.5 mA cm
width; voltage to ca. 300 V and the gel was "run for 2h".
Fixing and staining
The gel was fixed in an aqueous solution of perchloric acid
(8%, v/v) for 2 min, rinsed in distilled water (1000 ml) and
stained with Comassie brilliant blue (0.04%, w/v) in perchloric
acid (3.5% w/v) for 1 h at 37°C.
Enterobacteriaceae
The following tests were used to characterise the organisms
isolated on Violet Red Bile and Violet Red Bile Glucose agar,
and to determine whether or not sulphite preservative influenced
the composition of the Enterobacteriaceae: The Gram stain was
determined by the method of Kopeloff and Beerman (1922). The mode of breakdown of glucose under aerobic or anaerobic
conditions was determined using the method of Hugh and Leifson
(1953). The presence of catalase was tested by the application
of hydrogen peroxide (6%, v/v) to a PCA slope culture (18 - 24 h
at 37°c) of the test organism. The production of oxidase was
tested using the method of Kovacs (1955). The ability to grow
on MacConkeys medium (containing crystal violet and bile salts)
and Eosin methylene blue (EMB, Oxoid CM 69) agar was noted;
incubation was at 30°c for 48 h.
The motility of strains incubated in TSB for 18 h at 30°C was tested by the hanging drop method (Collins and Lyne, 1976).

82.
The API 20 E scheme (API, Basingstoke) was used to assess the
following characteristics;
The ability of a strain to produce 3-galactosidase, arginine di
hydrolase, lysine and ornithine decarboxylases, urease, tryptamine
deaminase, indole from tryptophan, hydrogen sulphide from sodium
thiosulphate, acetoin from sodium pyruvate, nitrites and/or
dinitrogen from nitrates and gelatinase. Incubation was at
30°C for up to 48 h.
The ability to grow using citrate as a sole carbon source; incubation
at 30°C for up to 48 h.
The ability to ferment mannitol, sorbitol, rhamnose, sucrose,
inositol, melibiose, amygdalin and L (+)arabinose. Incubation was
at 30°C for up to 48 h.
The API 50 CHE scheme (API, Basingstoke) was used to determine
the ability of a strain to ferment the following carbohydrates:
1 Glycerol 13 Mannose 25 Esculin2 Erythritol 14 Sorbose 26 Salicin
3 D Arabinose 15 Rhamnose 27 Cellobiose4 L Arabinose 16 Dulcitol 28 Maltose
5 Ribose 17 Inositol 29 Lactose6 D Xylose 18 Mannitol 30 Melibiose7 L Xylose 19 Sorbitol 31 Sucrose
8 Adonitol 20 a Methyl Mannoside 32 Trehalose
9 3 Methyl Xyloside 21 a Methyl glucoside 33 Inulin10 Galactose 22 N Acetyl Glucosamine 34 Melezitose
11 Glucose 23 Amygdalin 35 Raffinose12 Fructose 24 Arbutin 36 Starch

83,
37 Glycogen 42 D Tagatose 46 L Arabitol
38 Xylitol 43 D Fucose 47 Gluconate
39 Gentiobiose 44 L Fucose 48 2 Keto Gluconate
40 D Turanose 45 D Arabitol 49 5 Keto Gluconate
41 D Lyxose
Incubation was at 30°C for up to 96 h.
Data from all the tests mentioned above were converted into di:
binary data and analysed by the Clustan numercial analysis
package (Wishart, 1978). The data were sorted by the similarity
coefficients of Jaccard (S ), Gower (S ), and Simple MatchingvT G(Sg^); clustering was by single linkage (nearest neighbour),
complete linkage (furthest neighbour) and average linkage
(unweighed pair group - UPGMA) methods. Identification was done
by reference to culture collection strains or to a data base
(API Floppy Disk).
Culture studies
All media used are listed in the Appendix (pp. 334 - 335)
Turbidometer culture
The test organism was inoculated into the test medium (90 ml)
in an Erlenmeyer flask (250 ml) and incubated until a cell concen
tration of ca. 1 X 10^ bacterial cells/ml medium or ca. 1 x 10^
yeast cells/ml medium was attained. Samples (1 ml) were transferred
to sterile medium (500 ml) in modified flat-bottomed Quickfit
flasks (2 2). The flasks were maintained at the desired temperature

84
in a through-flow water bath and were stirred with magnetic stirring
bars. For aerobic studies, filter-sterilised air (Air Products,3 -1Bristol) was used to sparge the cultures (20 cm s ) via Dreschel
bottle heads (Q&Q 27/3/13) fitted to the necks of the flasks.
Flasks were fitted also with modified fermentation locks containing
'Lysol' (20% v/v) to prevent aerosol production (Fig. 9). The
optical density (A = 600 nm for bacteria, 540 nm for yeasts) of
the cultures was monitored continuously using a spectrophotometer
(Pye-Unicam, UV-VIS) and recorded on a flat-bed pen recorder
(Servoscribe). Culture fluids were drawn through sterilised 'through-
flow' glass cuvettes by a peristaltic pump and returned to the
culture flasks via side-arm attachments covered by rubber seals
(Subaseal). Narrow bore P.T.F.E. tubing was used to draw the
culture fluids out of the flasks and silicon rubber tubing was
fitted around the peristaltic drive shaft. Samples (1 ml) of
culture fluid were withdrawn aseptically from the culture flasks.
With yeast cultures, the 10 dilution in h Ringers was sonicated
for 15 s (Pringle and Mor, 1975) at an amplitude of 6 - 7 microns
(peak to peak) in order to disrupt clumps of cells (P. Thompson,
pers. comm.). Serial dilutions were made in h Ringers (9 ml) and
from appropriate dilutions, samples (20 yl) were pipetted onto
the dried surface (37°C, 1 h) of PCA. Using the technique of
Miles and Misra (1938) 32 replicate drops were accommodated on
a 9 cm diameter Petri dish (Sterilin).
Batch culture
The test organism was grown in the test medium to a cell6 —1 5 —1concentration of ca. 10 ml medium (bacteria) or ca. 10 ml

85.
medium (yeasts). Samples (0.1 ml) were transferred to sterile
medium (90 ml) in Erlenmeyer flasks (250 ml). Flasks were maintained
at the desired temperature in a water bath and left static, or
stirred with magnetic stirring bars or shaken (88 reciprocal
movements/min through a distance of 5 cm). For aerobic studies,
loose-fitting cotton wool bungs covered with cotton gauze were
inserted into the flasks whereas for anaerobic studies, 250 ml3 -1Erlenmeyer flasks (Q&Q FE 250/4) were sparged (20 cm s ) with
filter-sterilised, scrubbed (Appendix, p. 335), white spot dinitrogen
(Air Products, Bristol) via modified Dreschel bottle heads (Q&Q
27/3/13). A sample (0.5 ml) of culture broth was withdrawn
aseptically from the culture flasks and the optical density (A =
600 nm, bacteria; 540 nm, yeasts) read in microcuvettes (Sterilin)
on a spectrophotometer (Pye-Unicam, SP 600 UV-VIS). Cell viability
was estimated as in Turbidometric culture (vide supra).
Preparation of sulphite solutions
Stock solutions of potassium metabisulphite were prepared in
oxygen-free distilled water, filter-sterilised and kept at 4°C
under a blanket of filter-sterilised dinitrogen. The concen
trations of total sulphite were determined iodimetrically and
by addition of stock solutions to Ellman's reagent - 5,5'-dithiobis-
2-nitrobenzoic acid (DTNB) using the spectrophotometric method
described previously (p. 64 - 66)• Sulphite solutions were
added aseptically to cultures in mid-exponential phase, 2 fold
dilutions into batch culture flasks or incrementally until stasis
was observed on the flat bed recorder in turbidometer culture
flasks. Thereafter, samples (1 ml) of culture fluid were withdrawn

86.
aseptically from flasks, free and total SO^ concentrations determined
as before (see p. 66 ), or after filtration or centrifugation (13,000 g-,
2 min) of the broth to remove micro-organisms.
Hydrogen sulphide (H^S) production
Filter paper strips (Whatman) were soaked in a saturated
solution of lead acetate, dried at 70°C, and clipped onto the
outlets of modified fermentation locks or filters of culture flasks.
Binding or neutralising agents
Solutions of acetaldehyde (BDH, Poole) or hydrogen peroxide
(Sigma, Poole) were made up in deoxygenated, distilled water,
filter-sterilised, and kept in tightly-stoppered bottles at 4°C.
In order to bind completely the free sulphite in culture flasks,
acetaldehyde was added in excess whereas for neutralisation,
hydrogen peroxide was added in an equimolar concentration to
sulphite.

87
5x10
1x10
S 5x10roOE
5x10
400 300 200Electrometric deflection ( mv )
Figure 2. Influence of sodium metabisulphite concentration on
electrometric deflection as measured by the SO^ probe.

88
>1(0yiUlfdu
• H
4J
is8-14
uaUlTJQ)•H*44-r4
fgg01j::4J
■S143Ulfd01BUlfdCo
• H4J(dVsuc0u
8U40
143UT314
1-UCO
OosO
8CNJ
oo CJIR( N
O
nO
fSI
%3Cn•Htui
oo( N
<t( N
O CM COÔ
•-t o cbAbsorbance (412 nm.)

89
>1rdininIdc•rHin•g04-1
U•r-lin5IQ)■u•Har-43in
T30)kId(UEinId
c0•H■Pfdu4J§0u
%gT )r4Id+ j0)u(d
4-1o
£3UT3Pg§4J
w
ouoCn
•H[14
Oo
o
LO
O
tQjU
( 9 £ S = \ ) ^4!^^ d D u e q j o s q y

90.
O
LO
LH
•H
tnCTi
•H
CNJLD mo o{ tiJU OES) a D u e q j o s q v
oo o•rH
OJroSI
3Ln
"oeE

91
co■H+Jk4Jcmuc80)-P(T3•§,r-43to0 -p
1P3M<3i(DÆ4JC•H
§-H-Pfl3Ppig 8 OUQJ-P■Px:aI— i3to
(Puc%euips8M
VO01p3•S'tp
o(/)oo
oo o
<po
ooo oo o
( O E S ) S D u e q j o s q v

92 .
Figure 7 . Interference by thiols in the measurement of sulphite
concentration. Detection by means of formaldehyde
treatment.
V U
EClrsi5
1-0
<uucro_oS 0 • 6
0-2
0
0 200 400-1
600
j jg SO-ml
Key : O sulphite and thiol; # thiol;
□ sulphite and thiol + formaldehyde (50 mM)
■ sulphite + formaldehyde (50 mM)

93.
Figure 8. Interference by cysteine in the measurement of
sulphite concentration. Quenching of cysteine by
oxidation.
1 '4 r -
Êccvi
OJucroJDL.O\AXD<
1 0
0 6
0 2
0-o
Key:
0 400 800/jfDol Cysteine ml"
1200
# Freshly-prepared solutions of cysteineo
O Solutions of cysteine stored aerobically at 25 C
for 20 min.

94
inQ)•H1Vin
Q)sr 4au>14J•H>•H-P•HWc<uI•Ha
f-43tnc•pm3cn3Sa
SEi53w0i•H
BrH(X,
CTi0) I P&•P
(_QJ
OWOJa:c_m-Cw
Q.E3Q_w
m
Q)c/)
u(U■4' ■OJEPoJCCL8w(UQ l00
a;COCOzS;
<u
3W

95
RESULTS
Spectrophotometric method - theory and practice
The sulphite ion reacts with many organic disulphides to dis
place a thiol anion thus forming the organic thiosulphate, commonly
referred to as a "Bunte" salt.
2- ^(1) RSSR + SO^ ^ RSSOg + RS (see Fig. 10)
A large excess of sulphite ion is normally needed to estimate quanti
tatively the disulphide and the thiol anion is usually titrated
with silver nitrate (Cecil and McPhee, 1954).
Some disulphides e.g. 5,5'-dithiobis(2-nitrobenzoic acid),
"Ellman's reagent", and 4,4'-dithiodipyridine (Grassetti and Murray,
1957) are used also for the determination of thiol groups. The
disulphide is reduced to the corresponding thiol which absorbs
at a different wavelength from the disulphide. As the disulphide
is in excess, a mixed disulphide is probably formed:
(2) R'SSR' + RSH ^ R'SSR + R'SH
The disulphide absorbs maximally at 324 nm and the 5-mercapto-2-
nitrobenzoic acid at 412 nm (Fig. 10). When the concentration-5of the disulphide was ca. 5 x lO M, a mole ratio plot (Fig. 11)
of SO^^ indicated a stoichiometric reaction, 1 mole of SO^^
reacting with 1 mole of DTNB. The accentuated inflection seen
in this plot suggests that there are no intermediates formed and

that the 5-mercapto-2-nitrobenzoate moiety is produced as one
of the products in each reaction.
96
5 NO.
COOH
S NO.
COOH R-SH
COOH
R-S-S NO.
COOHreduction
HS NO.
COOH
5,5 - dithiobis (2-nitrobenzoic acid) Pale yellow
5 -mercapto-2-nitrobenzoate Dark yellow
Concentration range for SO determination
At a wavelength of 412 nm and a light path of 1 cm, 0.04 ug
SO^/ml was equivalent to 0.001 absorbance units (SP6-550 UV-VIS
spectrophotometer; Pye-Unicam). A straight-line plot (Fig. 12)
was obtained up to an absorbance of 2.04 units, this being
equivalent to a concentration of 950 yg SO^/ml. To avoid saturation
of DTNB and loss of SO^ from the system, the flow of SO^ in the
carrier gas was diverted by means of a 'Y ' junction adaptor to
duplicate receiving vessels containing the reagent.

97
Stability of sulphite and formation of thiol-4Sulphite solutions containing EDTA (1 x 10 M) maintained
under a blanket of dinitrogen at ca. 4* 0 were stable for at least
60 rain. The coloured complex was formed within 2 min of the
addition of SO^ to DTNB and, although there was no appreciable
loss of colour for 60 min (Table 10), the absorbance decreased
significantly with prolonged (18 h) aerobic storage due to
oxidation of the thiol anion (Humphrey et ai., 1970).
Rate and efficiency of SO^ uptake by DTNB
For free and total SO^ determinations in samples (5 g) of sausage
meat, a 900 s reaction time gave a 95% and 100% recovery of SO^
respectively (Table 11). Several receiving vessels containing
DTNB connected in series to the primary vessel did not recover3 -1any measurable SO^. A carrier gas flow rate of 20 cm s
-1(equivalent to ca. 100 bubbles min in the receiving vessel
containing 50 ml of DTNB) allowed complete harvesting of the SO^
coming from the reaction vessel.
Stability of DTNB
The observation that DTNB per se showed no tendency to form
the highly absorbing thiol over a storage period of 28 d (pH 8.0;
25^0) is in accord with that of others e.g. Wedzicha and
Johnson (1979); Wedzicha and Bindra (1980). The sulphite ion
reacts quantitatively with DTNB over the range of pH 6 - 9
(Humphrey et ai., 1970); above pH 8.0, however, there is

98
UinCN
fdܕHJO2CDId%uscn
IU-lOSAki8
§
§•H
IUc•Hcw0(UB5iwCw
CDr—I-g&H
Q CN r- TT O CN 'd'o 0 r~ r~ 10 CN m CN cnin O q O CN cn 1—1 r"<T>
Q d d O O O I—1 1—I
Q Q cr> r-H Q m CNO o rH lO 0 r- mo O q rH m m in cn CNm o rH o O O O p CN
C7> cn 1—1 lO in 10cn CN lO cn r- TTq r-H m ’d* Z m CNin d O O Q 1—4 CN
cn r—( r-H r-H m in in(U o CN 10 cn r~ «d*P fe! r-H f-H cn "sT m cn CNg oi ZC o o O O O P CN•H2
CN CN CN 'd'o r-H 10 cn 'ïf lO mrH r-H cn m cn CNr—io o O o O rH CN
*o cn r-H rH cn Ip lOQ cn r-H in CN oo O q f-H m in pd d d d d d P CN
r—I0P< CQ U Q k o
0 73V PidDc tifd Idf—4 PCQ w
UIdCB58a
T30)■Ptn(Up-P§

99
Table 11. Rate of SO^ harvesting by DTNB (5 g sausage)
Time (s) Free SO^ (%) Total SO (%)
0 0 0
150 32 47
300 90 96
600 94 99
900 95 100
1800 100 100

lOO
some evidence of breaking of S-S linkages, probably due to attack
by OH radicals, leading to the formation of the highly absorbing
thiol. As it was considered that a DTNB solution buffered at
pH 8.0 would be consistent with efficient trapping of SO^ from the
gas phase, it was adopted in the present work.
Influence of sample alkalization on total SO determination
Bound sulphite compounds are dissociated very slowly in acid
solutions at room temperature. Vas (1949); they ^re released
rapidly under alkaline (pH 9 - 11) conditions (Ponting and Johnson,
1945; Brown, 1977). Although extended (60 min) distillation would
be expected to dissociate bound SO^ moieties .(Shipton, 1959) it
has been suggested that short (15 rain) distillation periods may
result in a reduced yield (Wedzicha and Bindra, 1980). Alkalization
of sausage homogenate for 30 min before acidification and
distillation did not result (Table 12) however in a significantly
greater (p = 0.05, paired t-test) yield of total SO^ with subsequent
measurement by the method developed in this study.
Interference with DTNB
Unsulphited sausage meat
Using the spectrophotometric method developed in this study,
no interference from components of freshly produced or stored
unsulphited sausage was detected with sparging (free SO^ determin
ation) or distillation (total SO g determination). If sausage (0.1 g)
was added directly to DTNB, an inflated blank value resulted; the

101
0aeu
•HU4-)Q)
g4J
1kl4JU<U&i-PC
•H
So■p(H0
gou0)klcog
•H•pIT3N
•HI—I3
(p04-)UO
« pipw
JQ5
O C r o O
•H <U 4-Jr—I nfl À r-iE I—IW 4 J
WO ^
4 J D
ilLO C CN O
■P 4 J
; âip•Hffi O
O " PttJ u2 tO(PP5Z
1I. au4 JQ)g43
!4 JU
&
i*ftjw
OQ m CN TT 1 0 iH CN 1—1 CNr ~ CN LD CN r » 00 lO O CNCN r o r o r o r o •U* UO m
<
M
CN 10 0 CN 10 f—I en CN Oen rp 0 0 UO 0 0 oo r oCN r o r o r o r o 'd i UO
lÛ 00 CTi
Q)
W
S
tn
St n3 .

102
interference by these unknown components of meat or microbial origin
increased during storage at 4°c( Table 13 )
Thiols
Cysteine reacts with DTNB to produce a complex. There is a
straight line plot up to at least 1050 yg freshly prepared cysteine/
ml (Fig. 8). Such interference was rarely encountered, however,
in routine estimation of SO^ concentration in sausage or broth
cultures. Most thiols will oxidise in air after ca. 20 min
at 25°C and thus fail to form a chromogen with DTNB (Fig. 8).
The addition of 0.01 ml of formaldehyde (50 mM) to a sample
containing sulphite and a thiol leads to the formation of
formaldehyde sulphoxylate which does not react with DTNB (Fig. 7).
Electrometric ion-selective probe method
The theory and use of gas-sensing membrane probes for the
determination of SO^ in solution has been discussed by Bailey and
Riley (1975). Such probes incorporate an SO^ permeable membrane
so that a hydrogen ion-sensing electrode is separated from the test
solution by a buffered aqueous solution of hydrogen sulphite.
In this study, the concentration of SO^ was usually maximised
by poising the test solution below pH 1.0 before analysis.
The equilibria associated with the hydrolysis of SO^ in aqueous
solution can be summarised (King et al., 1981) :
(3) SÜ2 + H^O ^ HgO.SOg

103
Table 13. Interference with DTNB from components of stored
(4^0) unsulphited sausage
Days Apparent SO^ concentration(yg/g sausage)
0 8* 0^
1 6 0
2 0 0
3 21 04 14 0
8 41 0
* Direct addition of sausage (0.1 g) to DTNB reagent
-j- Method for total SO^ determination
Table 14. Interference in total SO^ determination of unsulphite
sausage*
Storage at 4°C Apparent SO(days) 1(yg SO2 g sausage ;
0 12
1 4
2 0
3 26
4 17
5 34
6 0
7 12
8 15
* Shiptons (1959) modification of Monier-WilUams's (1927) method

104.
^ +(4) HgO.SOg + H^O ^ HSOj + H^O
(5) HSO]" + H^O ^ SOg^" + HjO*
The use of the probe in this study was based largely on the methods
of Brown (1977). Two technical points need to be stressed. Firstly
chilled centrifugation of homogenised samples was needed because pork
fat tended to coat the membrane of the probe. This resulted in
(a) a loss of sensitivity and progressively lower estimations of
SO2 concentration and (b) a short membrane life. The poor recovery
of both free and total SO2 in samples can be explained in part
because the SO2 level of sausage serum only was measured without
regard to SO2 trapped in the fat or solid phases of centrifuged
sausage homogenate. Secondly, a deflection (mv) was registered
on an EIL pH meter (monovalent cation setting) following immersion
of the probe in the acidified specimen. At very low concentrations
(< 50 yg/ml) of SO^, the peak deflection after 2 min was often
difficult to measure because of (a) a slow response period and (b)
interference by unidentified factors. At high concentrations
(> 400 yg/ml) of SO2 , the peak deflection after 2 min was easily
measured but the recovery period (dialysis against distilled
water) required to remove excess SO^ from the internal buffer
solution of the probe was seldom less than 10 min.
Concentration range for 50 determination
The probe could detect as little as 5 yg SO^/ml and a plot
of the logarithm of preservative concentration against the-3electrometric deflection (mv) was linear up to at least 5 x 10 M
Na2S202 (Fig. 2).

105.
Steam distillation method
The modification (Shipton, 1959) of Monier-Williams's (1927)
method was used for reference purposes in this study.
Interference
The steam distillation method (Table 14) was prone to inter
ference. This resulted in an inflated blank value of ca. 15 yg -1SO 2 g sample from unidentified compounds in unsulphited sausage
Influence of sample size
Although analysis of ten replicates of sausage meat gave
identical mean recoveries (412.6 yg SO^/g) for 10 g and 20 g
samples, the variation in the former (standard deviation = 38.18)
was greater than in the latter (standard deviation = 22.84).
The better reproducibility obtained with the 20 g sample led
to its adoption for all assays based on steam distillation.
Comparison of methods
Total SO2 determination
In direct comparisons, the spectrophotometric method proved
to be less variable than the steam distillation method (Table 15)
There was no significant difference (Students t-test p = O.Ol )
in the level of recovery of SO2 and an excellent correlation
between the two methods (Spearman's rank correlation coefficient
(r = 0.99 ). A regression line (least squares method) fitted2to the data had a coefficient of determination (r ) of 0.96

106
wT303343(UE0)g0klOiTOc<dܕHU43(UE0430A0ki43U0)aM
(30•rH43IdrHrH•rH43tn•rHTDE QJfd CP(U Id43 tXtn d
Id(U tn3343 C3
•rH(3(U CN
O> w43QJ r—133 Id
430) 0U 43cId <4H•H 0f3 c> 0
•rH144 430 IdEtn •H•H 43tn tn>1 QJr-HId klC 0< 144
IT)
I
O'in
co•H 43 <0 ♦ f—I ^—- I—I O'43 O W CN
•H ^ T3
U •rH kl 43(U§43
1A in2 t;&
0)id0•HrH1
o 1— 1 CN in cn m O in CN O COOCOr- in cn CO cn VD 'd' ID CO ID 1—1
m m m m m m m m m m m
IDm Q cn O ID O O in o CNCN O m •d* cn 1—1 «d* O cn CO
'd' m «d* m m
COCNCN
IW
mCT>m
in 1—1 r—1 CN O O CN r~-1—im
cnr—1t—1 CO o 1—1 00 cn cn CO cn i-H
'd* 'd' m m m m m
CO cn I X
Q>Wdfd[/)
CP
38CP3.

107
(intercept (y) at 0.08, slope (m) of 0.98). A more detailed
comparison between twenty duplicate samples confirmed that there
was no significant difference (Paired t-test p = 0.01 ) between
total SO^ recovery and a regression line fitted to the data was 2described; r = 0.99, y = 5.03 and m = 1.00 (Fig. 13). Comparison
of the steam distillation with the probe method showed that the
latter was less variable. The differences in SO^ recovery between
the methods were significant (p = 0.05) when analysed by the
t-test and correlation between the methods was poor, r^ = 0.67.
Comparison of methods for total SO estimation after freezing of
the sample
A limited study showed that freezing (-20^C) of samples of
sausage meat for up to 48 h had no appreciable effect on recovery
of total SO^ by the steam distillation or the spectrophotometric
methods. Although the latter recovered on average more SO^
(x = 401.66 y g SO^/g) before sample freezing, this level was
not significantly higher (p = 0.05) than those achieved by either
method after freezing (Table 16). The error, as indicated by
the standard deviation (on-l), was very small (an-1 = 2.89)
for the assay of fresh sausage but relatively large (an-1 = 12.65)
for the assay of stored freezer material.
Free SO determination
The spectrophotometric method recovered significantly more
free SO than the probe (t-test, p = 0.05) and was less variable

108
c0 CP•H c43 -Hfd N1-4 QJ1-4 QJ * O <0•H k4 CM CO o O <D rH CN43 <44U) m CO <D CO <Ti CP ro•H kl O GO CP O 'd' 00 CN'Ü QJ Tf m m 4" ro ro
43E <44CM fd <CO QJw 43
W1-4fO4-)04J
U40>4uQ) CP> (30 •HÜ N * O mQJ QJ CN CN CD CN CD ^ <pki QJ
k4 m n in r4 r4 00 CNC <44 CP o rH CO O rH0 m Tf Tf m "ff ro
k4QJ33 43<44
CO 0 c'd* •Hkl43
U QJ0 ’ EO 0CM 431 O33ACP 0c kl•H 43N u 5QJ QJ CQJ Oi -Hkl CO N * ID144 QJ CN «fj* ro ID <T>QJ 00QJ kl .H <n <3) 0 Q r4 •rH <44 O <Ti O 0 O 0 CNA ro 'd* ’d' 'd' 'd'QJ
k4œ 0<44
<44 QJ0 PQQJUa0)01-4<44cH
. QJID 43rH fdÜ rH0) •r4 1r-H 1-4 (333 04 rH CN ro 'd' m 1 X ofd Q)E4 (X
QJ
W0(0w
Cr>
8O'3.

109
(Table 17). Correlation between the methods was very good (r =0.99)
according to Spearman's rank correlation test and a regression line2fitted to these data had an r value of 0.92, (y = 0.44, m =0.88).
A more detailed study of 20 duplicate samples confirmed that the
spectrophotometric method recovered significantly (p = 0.05,
paired t-test) more free SQ^ than the probe method. A regression2line (Fig. 14) described these data (r = 0.98, y = 4.41 and m = 0.88)
Estimation of SO^ concentration in sausage seasonings
Analysis of sausage seasonings (RHM 599) used commercially in
the preparation of sulphited or unsulphited batches of sausage
is summarised in Table 18. The iodimetric method, used by the
large-scale manufacturers for determination of SO^ was prone to
error due to unidentified moieties in the unsulphited seasoning.
On average this error accounted for ca. 14% of the apparent SO^
concentration and explains the consistently higher values obtained
using the iodimetric, rather than the spectrophotometric, method.
Sulphite binding
Sterile solutions of sulphite maintained under a blanket of
oxygen-free dinitrogen did not bind significant amounts of free,
or cause the loss by oxidation, of total SO (Table 19). Aerobically2
stored solutions of sulphite led to an initial binding of 15%
of the free SO^, a level which was maintained during the period
of incubation. All meat ingredients of sausage bound significantly
(p = 0.05) greater amounts of free SO^ and catalysed also the loss

110.
0)en(tJw5cfl<w0
en 0) I—j1cnQ)•p(0o•HrH6 M
8Q)dJktMH<440
g8Q)kU8enT3243i<440C
8•H
1
•§H
I04 tT> LT» S om CP r~ en<Ti
CD nUN CN) O CDO O f—1 rH rHrH rH rH
(US'en3
S
tpCN)8S
0 •H U 43
1I438S!
CPin
O NT* CP UN CN O lO O rH UN<x» NT m ro UN UN CD UN UN NTr-H rH rH rH rH rH rH rH rH rH
euSü•HrH
Iin <0 CD CP
o CPI—I ON
CP o 00NT rH
I
I X G D
Ien•K

Ill
tn
•H§cn(C3Q)cnQJS'tn3ftJcnMH0
0•H
k43C8GOU
CO
QJ1-4•s54
U•Hko)g43
Ik43U&
aGO
■§tn3
■85Q)2
m
I8g•H43•H
43Us•HQ
I43
U-r4k43
I8
CPCP -H CP G LO 0
tnI $CX CO
roNTLO
oNT<P
CP00
sCOCN
r-CNLO
OCOCPoro
CNlOLO
SNTCNro
CPNT
SCNCPCN
COroLO
oLDLD
CO O CNo ro 00LD LO V)
Q 0 o o OLD LD UN CN UN LDr—1 00 CN LDLO LO (-4 ro NT NTro ro ro
CN CN CN CN CN co o o o O oCO CO CO CO CO CO+ 1 + 1 + 1
3>dPO'
TJ cnQJ bT! 0T) 33It) 43
QJCP EG•H (3G •H0 kcn 43(tJ QJQJ Scn 043CP 0G 33
•H &S 03 kcn 43cn C3(CJ Q)
Û4X cn•He QJ
33QJ 43CPfCJ 04cn 03(CJ kcn QJ4 8
•HCP QJCN 33
Q 43CO •HCP3. bQJ
43
cn 8QJ CP 43cn G QJQJ •H33 G+3 0 cnG cn fljQJ Id 5k QJ(CJ cn QJA (3
r— I GG 1 QJ•H CP k
QJcn CN <44QJ o kk CO QJ3 43CP CP G•H 3 •H
0* 4— Z

112
w4Js•H%U•H
Q)S'M33Ü)
<4-10&•HU3aÜg•H'Oc•HXII
•HÛ4rHa
CP
854
430)01—4CN
o (P NT LD LO c~ 00 ro O 00 LDCO lD vD LD LD ro CN 00 00 NT r—4X1—4 1
(d 043 X05-tcK>
T3G30 CP UN ro 4^ ro CN LD UN f-HX LD LD LD NT r- UN CN CN rHàP
O)LD (U in CN NT 1—4 ro 1—4 UN O c~ Ok LD 00 O CN LO CN NT O' LD CN
k rH 1—4 CN CN ro LO
f—1(d43 LD NT 1—4 CP O LD CN o 1—4 o-P o ro 00 LD NT r- O r- CP 005- CN CN 1—4 ro NT rH ro NT LO
T3GG LD CP UN ro 1—4 r- CP CP O' CPQ LD UN NT UN ro LD NT ro 1— 4X
33dP
<U05-t CO (U LD O' CN 00 NT CN ro CN 1—4 CPk NT LD O CP LD CP 00 CN LD NTIh 1—4 rH CN 1—4 ro ro NT in
r—HfOX X O CN NT 00 ro LA LD NTp ro Q CN CN C'- LD CN LO o54 NT NT ro NT UN CN <—) LO LO LD
13Q UN 13N O NT LD CP UN ro LO NTX CN CN ro 1—4 1—4 LO O' CN rH 1—4dP
Q)O 0) CN ro r—H CN NT ro o NT O'k O 00 CP O LD NT 1—4 roEu LT) NT ro «d* LO CN 1—4 NT UN UN
1—4id MX O UN CN 1—4 ro ro NT CN CN rop r" NT ro O NT 00 1— 1 O CN54 LD LD UN UN LD LD LO LD LD LD
X3xC 3^ X id ük k (d eu •r4Q) 0 0 Q) S U rH X X
r-4 A Ci 6 •H 0 0 0•H >1 en X k k kk c G V rH T) 4 - 34 0 X eu Xm 3 (d fd rH G X cn k G fd GX QJ eu ÿ g •H fd 3 eu 0 G Ocn rH 1—4 X X k X 2 k fd U fd u
8
I43JB
II8G•HS0%V
1•H
I
CN
8CP3

113
of greater (p = 0.05) amounts of total SO^ (Table 19). With the
exception of mechanically-recovered-meat, binding and irretreviable
loss of SO^ increased with storage time in the presence of meat
ingredients. On average, rusk bound only 30% of the free SO^;
after 6 h, however, the absolute level of SO^ had fallen by 40%.
^ 2 loss from stored British fresh sausage
Influence of site
There was no significant difference (p = 0.05) between the
levels of total SO2 recovered from the centre and from the surface
layer (0.25 cm) of sausage at the storage temperatures used in
this study (Fig. 15 a,b,c). The level of free SO^ recovered from
the surface was significantly lower (p = 0.05) than from the
centre at all three storage temperatures, 4, 10 or 22^0 (Figs.
15 a, b and c respectively).
Influence of temperature of sausage
Total SO 2On average 24% of total SO2 added to the ingredients in the
bowl chopper was lost irretreviably; loss of total SO^ occurred
at a rapid rate from samples taken immediately following manu
facture to the 3rd day of storage but at a much reduced rate
thereafter (Fig. 16). Although the initial level of SO2 in the
freshly manufactured sausage did not influence the rate of loss
it did dictate the final level on day 8. Loss was related
directly to temperature. Thus after 8 d at 4°, 10° or 22°C,
82%, 79% and 76% of the total S0_ level (at t ) remained.2 o

114
Free SO^
On average 27% of the total SO^ added to sausage ingredients
in the bowl chopper was bound reversibly; thereafter the loss of
free SO^ from day O to day 5 at all storage temperatures conformed
to the equation:
(6) log^^ SO^ = mx + b
where x = storage time in days, m = slope/gradient and b =
log^Q SO^ at t^. After day 5, the rate of loss diminished
(Fig. 16). The rate of loss was significantly different (p = 0.05)
at the 3 storage temperatures (22° > 10° > 4°C). After 8 d at
4°, 10° or 22°C, 27%, 18% and 11% respectively of the initial level
of free SO^ remained.
Binding of free SO^
The rate and extent of binding was related directly to the
storage temperature. Thus only storage at 4°C prevented substantial
loss of free SO2 during the period day 0 - day 5.
Loss of SO^ from sausage meat stored in Petri dishes
Although in these experiments 600 yg SO2 were added per gram
of sausage ingredients in the bowl chopper, 150 yg were lost
irretrievably The absolute level of total SO2 in sausage meat
(100 g) stored in sealed square Petri dishes at 4°, 9°, 15°,20°
or 25°C diminished rapidly from day O to day 4; thereafter the rate
of loss decreased (Fig. 17). The initial rate of loss was related

115
-U•H3
8M-l0C0•H-Uu(T3Q)kl§klT)•HU(T3U•HONC
2
CMI04J&UklE
u-i0C0•H4JnjEkl2
0)kl3en•Hk
L OCN
8+CQZg
S
CQZËO oL O
K.0NS-§kl■p•rHC
OomCNU)•rHX!0•rHs :HJ•rHV
OLOrsi
unLD
LO O Oun
Ec
_0L X juay^ao] uoipui^xa jejoiu

116.
*CQ2gT3§S•Ha1-4dwtuo■Uor4O,0•rHHJ(Qkl
%3en•Hk
m
ovOô ô
ooo o
00
tu0cO
•HHJ3Ud)ucou
( U J U 3 t ^ 7 ) B D u e q j o s q v

117
■H
CM
<4-1
oovO
8CSI
co(N
O008vO
CnJ
CMI-1(Uua
•H[I4
CO(N CVI
O(N
vO (NI OOÔ Ô
Absorbance (A12nm. )

118.
Figure 13. Comparison of methods for the determination of
total SO^ concentration in fresh sausage.
400
1 30 _C
E S,3001 iro
-4- C71
i ^ 0 0
I 5c/)
100
0 100 200 300 400
/ug SO» g” sausage
Specfrophofometric method

119.
Figure 14. Comparison of methods for the determination of
free SO^ concentration in fresh sausage.
OJcnro{/)3ro1/1
"OoJZ<DÊcuJDa
OincnR
uOJQJ00co
2 0 0 r
150
100
50
0 50 100 150 200
yug so g"”* sausage
Spectrophotomefric method

120.
Æt/î0)u4-1
k0Vin
04-JOJ-U
•r-tx:ar-l3wQ)Q)
4-Jnc•T3
w (U0 r-H (UrH rHÛJ 04eu r4 (U& co r-H4J B A COro eu EC W u ro 0)0 ro CO uO) 144 roc P k <U 4-t0 0 3 P P•H Ü CO 0 34-1 U COroO rsi CM0 O O CM CMr—1 W W O Oen enD 1—1 1—!C rO rO (U (UfO 4-1 4-1 (U (U0 0 p(U E-r E-r kk3-P3 O e □ ■uq;a
c (U(U Cn3 3 u Ur—1 co u 0 04-1 3 0 O CMC 3 i-H CMM co. 3 1 X!( U
(Du3 , Cn •H
(D+Jg4J00
oo
oo 8 O
oo
oo
oo
oo§ o o
35esnes _5 ^qS 6rr

121
Figure 16. Influence of temperature on the loss of free
and total sulphite from stored fresh sausage
500'
o — o
300lo
N
200
100
0 1 U 5TIME (dl)
7 8
O Total SO , 4 C
Total SO , 10°C
# Free SO , 4 C
jL Free SOi , 10°C
□ Total SOU, 22 C Free SO , 22 C

122
Figure 17
6001-0
500
H- I l l I I
- I
400
ro eu
I booDroi/)
CJ1
f 200
en
100
o*-
0
Influence of temperature on the loss of free and total
sulphite from sausage meat stored in Petri-dishes
O Total SO,
< :o
4°C9°C15°C20°C25°C
3 4 5
Time ( days )
# Free SO) , 4°C
I : &■ " 20°C
▲ 25°C
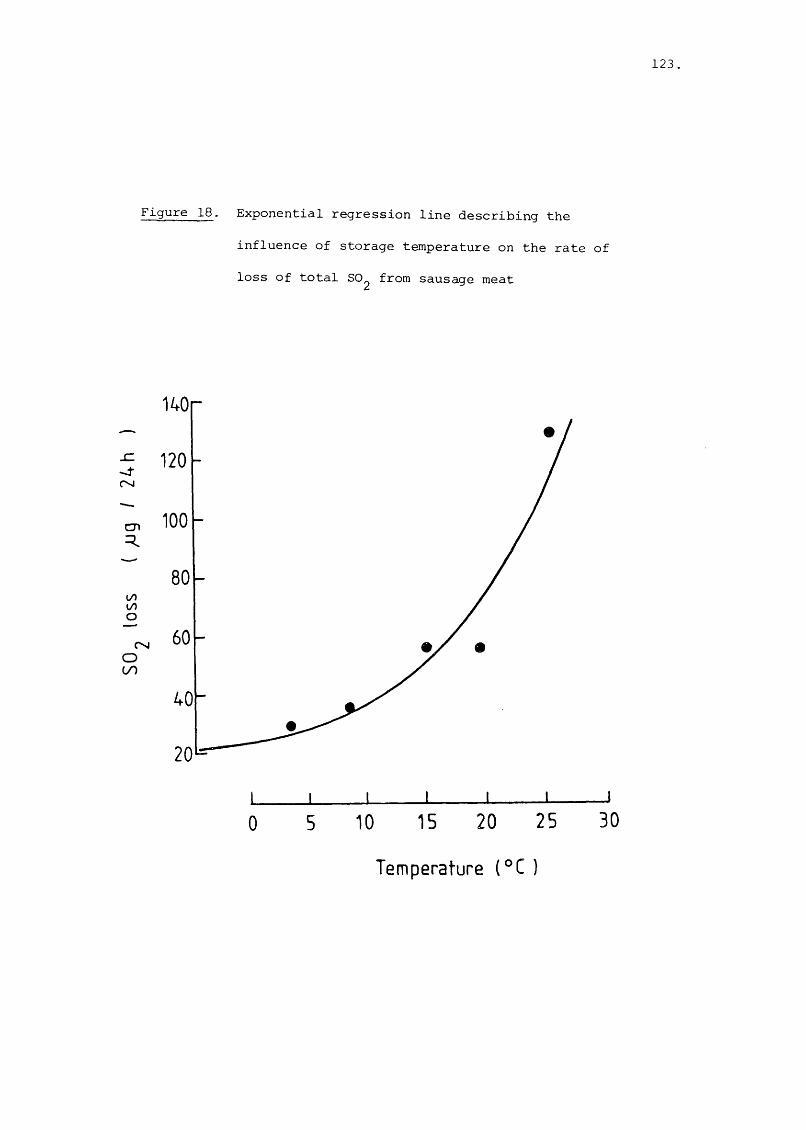
123
Figure 18. Exponential regression line describing the
influence of storage temperature on the rate of
loss of total SO^ from sausage meat
140
120
100cn
rsjO00
0 10 15 20 25
Temperature (°C )
30
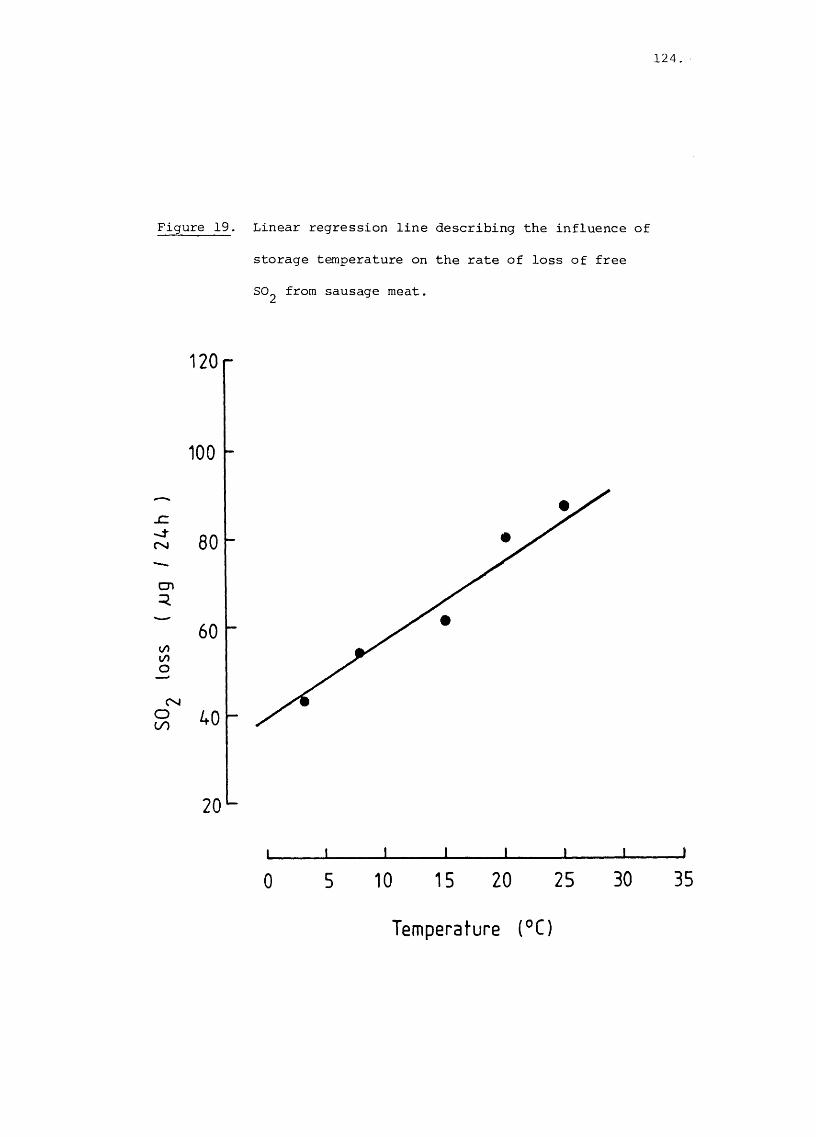
124.
Figure 19. Linear regression line describing the influence of
storage temperature on the rate of loss of free
SO^ from sausage meat.
120 r
100
_C
cn
0000o
80
60
CMR 40
20
10 15 20 25
Temperature (°C)
30 35

125
directly to storage temperature, a linear regression line (Fig. 18)2fitted to the data had a coefficient of determination (r ) = 0.98,
intercept (y) 2 34.5 and slope (m) = 1.92. Free sulphite levels
declined rapidly (Fig. 17) and loss was related exponentially (Fig.219) to temperature, r =0.87, y =21.76 and m = 0.06.
Sources and extent of contamination of British fresh sausage of
factory origin
Analysis of 35 batches of freshly made sulphited pork sausage
over the period 1.10.79 - 31.9.82 showed that Brochothrix thermo-
sphacta^ lactobacilli, Enterobacteriaceae and yeasts were always2present at levels greater than 10 /g; streptococci and pseudomonads
were present in low numbers (10^/g or less) only (Table 20). In
all cases the sausage microflora was dominated numerically by
Brochothrix thermosphacta (isolated on STAA medium), substantial
contamination with lactobacilli or pseudomonads was of infrequent
occurrence. Of the organisms noted above only the pseudomonads
and Enterobacteriaceae were present in significantly greater
numbers in the freshly made unsulphited product. With the
exception of the streptococci, most raw meat ingredients awaiting3use in the factory harboured large (> 10 /g) populations of all
of the members of the microbial association (Table 20). The
minced ingredients, belly and head meat, were usually more heavily
contaminated with all types of micro-organisms than were whole
pieces of lean pork. Back fat contained relatively low numbers
of contaminants, especially Enterobacteriaceae, whereas rinds,
which had been cooked, minced and chilled, were always heavily
infected (Table 20).

126.
c-H-HM0>1Su14-1
4-tO<Denn}cnP(pcn
CenCD44
4-1
x :en•H+j•H4lCQ4-1OCO-H4JfOC•rH
(T3•H
•§
Ü■HS4 - t0-ps■p
s
fOcnCD
P0co
CDI—IA5
en-p CD CNtel O r~ 1— 14-1 40 CD O O uo 1— 1
m o 1 • sf » CD • 40 1 Id.i-i O O 1 CN 'Ct* uo O CN rH 40 CD oU CN • r-4 • CD • 1 • 1 • 1 . Tf prcJ en LD . ro CN r-l r-4 ro CN CN • 0CQ CN ro V V V #— 4 4H
CDen rH73 Qtc in en CD E•H TT M ’ 'ey cdP CD cn
40 CD 40 M* CD 4073 O O 1 CN 1 CN • en • r- 1 ro 1 1— 1CD CN CTt CN CN ro O 40 CD M* r- CN CD rH 1
• CTt • r- • 1 • 1 ' en • CD en0 M • 40 • CN CN r-l TT • uo •0 CN Tf CN CNU P
< u
X ID CN uP in r» CD 400 40 ro OQt ir> CN . . CN UO ro uo • rH
r—4 1 en M CD 40 en 1 CN CD 40 en73 O • CN • 1 • 1 • CD • 1 0tC3 CN CTt uo en ro 40 1— 1 r"~ uo sf 40 p— 1CD 1Œ m in CN 1— 4 CN CN
V cn■Pa
.X pP c o 00 c \ en uût CN • r~ M* uo r— I r- Tf .
'ey ^ 40 • r-4 CN O uo 'cr 40 uo (p>1 O • 1 • CD • 1 0 enrH CN r o ^ uo 1 ro r-H CN ro 40 <;Jt Tf CDr-l 40 TT 1 1 1 CD rHCD CN 1— 4 CN CN en QtCQ CN V V V G E
cd cqttî en
AC r-~ en -W-P r— 4 CDO m • 40 ro uo • i— H en CD TfQt r - 40 «a» 40 UO CN rH CD rH
C O m Tf ^ 40 ro CN rH CN ro UO ro uo(C3 CN r— 1 1 1 1 1CD CN CN rH CN rHPI CN V V V V
73CD■P in r-l en CN•H 40 CD enC CD CD • ro CN r-4 • O CN CN • CN •Qt ^ CD M* en • r- 40 40 ro ^ uo ^ Tf1— 4 n3 CD • 1 • CD • 1 • 1 > 1 CDp en ro r-4 uo 1 ro 'cT CN M* ^ CD 40 r—1en g m ro 1 en M* QtC fC3 rH ED en CN sf CN CN ro rd
en
-H- rHCN CN 1
CD 4— en CN r~ •vT en■P CD CN 'D en CN ro M* O uo ro en .•p (ji •K CN 'CT CD • 40 • en 40 ro rH uo 'cr fdn ce) in • 1 • r~ . UO • 1 Q 3Qt en m r-4 uo 1 ro 1 CN ^ ro 40 ro Tf pr-t Q CN ro CN 1 1 en ct_iP (p rH CN CN CDco cn CO 'd' CN V V 4-> u
cn oCD CD rH
co Qt Qt iq rH m■u Qt Qt CD Qt 0ü en cn Qt U E rHId Qt (drc; en en en •H enQt 3 3 p •Pen M ü en CD cp c
M ü iq ■P 0 P•p 0 C U 0
P ü ü cd p uQ) fCJ 0 A CD
en rq rQ •p 0 0 A g■P -p 0 Qt 73 P ^ Î3cn ■u CD 3 CD p CDçd ü P CD ■P Z SCD p cq •P en C>H CQ A CD Ût w * -f-

127.
Influence of day of manufacture
Although the composition of the microbial association of the
sausages produced at the factory was always dominated initially by
Brochothrix thermosphacta and, with storage at a variety of temperatures,
by Brochothrix thermosphacta, lactobacilli and yeasts, the chilled
storage of carcass meat over the weekend influenced the extent of
the initial contamination (Fig. 20). Thus sausages produced on
Mondays or Tuesdays with meat which had been stored for upwards of
96 h at ca. 4°C, contained significantly (p = 0.05) more micro
organisms (x log^Q 7.27; Q = log^Q 0.62) than those produced on
Wednesdays or Thursdays with meat which had been stored for 30 h
(x = log^Q 5.85; a = log^^ 0.62).
Influence of initial population size on extent of growth of micro
organisms during storage
The initial level of contamination with micro-organisms was
related inversely to the extent of microbial growth during storage
at various temperatures (Fig. 21), a regression line fitted to2the data had a coefficient of determination, r =0.91 and y = 3.25,
m = -0.28. The data of Brown (1977) and Dowdell and Board (1968,
1971) were analysed also; those of the former were described by2a linear regression line r = 0.84, y = 3.44, m =-0.31 and the latter
2by r = 0.63; y = 1.97, m = -0.11.

128
Influence of sulphite and temperature on the size and composition
of the microbial association
Loss of sulphite
Analysis of the seasonings used in five experiments (3 commer
cial, 2 pilot scale), indicated that of the 580 \ig or so sulphite
added per gramme of ingredients in the bowl chopper, ca. 26% was lost
irretrievably and 23% bound reversibly during mixing. Likewise
comparable rates and extents of loss of free and total sulphiteo o o o ofrom sausages stored at 4 , 10 , 15 , 20 or 22 C were noted in
all the other four batches examined. For purposes of clarity, the
fate of free and total sulphite in one batch of sausage only is
shown in Fig. 22.
Development of the microbial association
As measured by the total viable count (Fig. 23), the presence
of sulphite reduced significantly both the rate and extent of
growth of the principal bacterial contaminants at all storage
temperatures (Fig. 30 a). The size of the climax populations
of yeasts was not affected significantly by sulphite (Fig. 30b).
Its presence enhanced however the rate of multiplication of these
organisms especially with storage at 4° and 10°C, (Fig. 24).
The preservative influenced the dominant bacterial contaminant,
Brochothrix thermosphacta, in as much as larger populations were
formed in unsulphited sausages as compared to sulphited ones
(Fig. 31a). The time taken to attain a climax population of the
organisms was influenced by both temperature and sulphite; the
most rapid growth occurred in unsulphited sausage at 22°C, (Fig. 25).

129.
The growth of Lactobacillus spp. in sulphited sausages at 22°C
was similar to that of Brochothrix thermosphacta; at 10°C or less,
the rate and extent of growth of the former, especially in pre
served sausages, was markedly reduced (Figs. 26, 31b). The rate
and extent of growth of members of the genus Streptococcus were
only affected by sulphite with storage at 10°C or less (Figs. 27,
32a). Judging from changes in the size of the populations of
pseudomonads and Enterobacteriaceae, sulphite was more effective
as a preservative, especially at low storage temperatures, against
the Gram-negative fraction of the initial contaminants of sausages.
Thus with the pseudomonads, no demonstrable growth occurred in
sulphited sausage stored at 4°C, whereas growth occurred at 10°
or 22°C. The rate of increase and eventual size of the populations
in sulphited sausages were always less than in unsulphited ones
(Fig. 32b). Moreover, the temperature of storage did not influence
appreciably either the rate or extent of growth of these organisms
in the unpreserved product (Fig. 28). Members of the Enterobacteria
ceae appeared to be the most sensitive indicators of the preservative
role of sulphite. These organisms formed large populations in
unsulphited sausages stored at 10° or 22°C but remained quiescent
at other storage temperatures, other than 22°C, or when free-1sulphite was present at concentrations > 25 yg SO^ g sausage
(Figs. 29, 33).
Influence of temperature and sulphite concentration on pH drift
There was little change in the pH of sulphited sausages at
4°, or 10°C (Fig. 34) during storage for 7 d, but a marked acid

130,
drift with storage at 22°C (Fig. 34).The marked changes in the pH
correlated with the formation of large populations of lactic acid
bacteria. In other experiments (see pp.157 - 164) the rate of
pH drift in sulphited sausage meat was associated exponentially
with the storage temperature, (coefficient of determination,2r = 0.79) thus supporting the notion that sulphite was effective
in preventing acid formation only up to a certain temperature
threshold. Changes in pH were invariably greater with unsulphited
sausage meat and were significantly larger (Students paired t-test)
than those in sulphited ones at 4° (p = 0.05); 9° (p = 0.05);
15° (p = 0.01) and 20°C (p = O.Ol). At 25°C, however, there was
no significant difference between the pH of the preserved or un
preserved product over the storage period of 2 - 8 days.
Influence of a high concentration of sulphite and temperature
During the course of examination of sulphited and unsulphited
batches of sausage produced under commercial conditions in a
factory, a particular batch was exceptional in having an extremely
high concentration of sulphite. Analysis of this batch gave some-1interesting trends. Of the 660 yg total sulphite g sausage
present initially, 270 yg were bound reversibly during manufacture
and, as with other batches tested, the absolute content of sulphite
decreased slowly with storage (Fig. 35). The rate of decline
of free sulphite was slower than in sausages containing normal
levels of the preservative (Fig. 35). Indeed, on day 7, the
concentrations of active preservative were 2 - 3 fold higher than
in commercially-made sausages containing legally-permitted levels

131
of sulphite. These differences in the rate of loss of the active
form of the preservative were reflected in the results obtained
from a microbiological analysis, especially with the populations
of the sulphite tolerant organisms, the yeasts. In contrast to
sausages containing legally permitted levels of sulphite, the
rate but not the extent of growth of the yeasts was suppressed by
the presence of abnormally high concentrations of the preservative
(Fig. 36a). The temperature of storage also influenced the doubling
times and size of the populations attained by day 4 or 7 (Fig. 39a).
The rate and extent of growth of Brochothrix thermosphacta were
severely curtailed by the high level of sulphite (Fig. 36b).
in the absence of the preservative, the doubling times and size
oÈ the climax populations (Fig. 39b) were of the same order as those
noted in other batches of unsulphited sausage. As noted previously
( p. 129), the temperature of storage influenced the rate and
extent of growth of the lactobacilli and streptococci. The former
group was not affected, however, by the high concentrations of
preservative (Fig. 36c). Indeed, the climax population formed at
4°c was significantly greater than in sausage containing the
normal levels of sulphite (Fig. 40a). The streptococci likewise
formed substantially larger populations in the highly sulphited
product stored at refrigeration temperatures when compared with
those developing in normal batches of sausage (Figs 37a, 40 b).
The response of the pseudomonads was similar to those in sausage
containing the legally permitted levels in that their growth
in the sulphited product was not evident until the 5th day of
storage at 15°C, at which time the level of free sulphite had
diminished to ca. 110 yg/g sausage (Figs. 37b, 41a). The marked

132
sensitivity of the Enterobacteriaceae to sulphite, which was noted
above (p. 129), was reflected by the significantly greater doubling
times and smaller climax populations (Fig. 41b) in the highly
sulphited product stored at 4° or 15°C (Fig. 37c).
Composition of the microbial associations
Influence of sulphite concentration
The identification schemes used in previous studies of the
microbial associations of British fresh sausage (Brown, 1977;
Abbiss, 1978; Leads, 1979) were such that subtle changes, if
any, in the composition of microbial groups would have been un
detected. As the results presented above suggested that the
Gram-negative bacteria were the most sulphite-sensitive members
of the microbial association, it was surmised that the changing
concentrations of free sulphite may exert a subtle selection such
that the composition of the populations of these minor members
of the association would vary, particularly between sausages
made with or without the preservative. This hypothesis was tested
by detailed analysis of the composition of the Enterobacteriaceae
and Pseudomonas isolates.
Enterobacteriaceae
Thirty seven attributes were used for the initial character
ization (Table 21) which assigned the isolates to twelve distinct
groups. Sixty-four representative strains from these groups were
characterized further using the API 50 CHE scheme. The Clustan

133.
<u
CO
CO
73Ctq-P<qS
§
r —I
8-H
%Q)Ütq•H
■p
As(DPCWIH0COÜ•H4-)CO•H
-PPcqÆu
CD1-4Eh
CN
OrH CN
CTi CNun
œ <T\
r- CN•3*
40 'T
in CNCO
'IfCO
CO 'q*CO
CN CO
CO
p p CD CD •p -p 10 CO p prH rHu u
O O inO CN
§
8 ° 8
8 ° 8
o o o o o
8 °
CO CO 40 CO CO
O O Oin m o
O
rH (3"» O00 rH O
CO CN o00 rH O
CO r ~ . «;)<CN r>. m
O O O O
«>c0CD
g&orH
8
P^ CD JP I
«q* -rlCN 73
CO CNA
•H73
CNV
•PA
UHo
o o o
° 8 ° 3
° 8 ° 8
>
I
8
Oo
8 S
in O
CO O CO
o 40 o 'q* OCO o cnrH
o O Q 40 O00 O COrH
o Q O CO oO o1—1 rH
o 40 Q O oCO O Of— 1 rH
o in O Q oo O1—1 rH
o CO o in 1—1in o CO cnrH
o O o O COorH
o O Q in oO CNrH
ms0 w•H G73 0CO CDCD 6 73p CD0 E
Ù 4 u PCO e cq 00 p s ip73 •pC 73 G GCD CD 0 CDS CDip 73 X i0 CO CD COG >1 SCD >1 0 CD P ÜÜ ■P X 0 •PC •H X i G ip 1—1CD rH -P 0 rHCO •H :? U 73 cqCD 4J 0 0 •P pP 0 p cq O CDCU S Ü s < s

134.
04 «q"
I—1 LD
o M*r-l CN
CT» CNin
00 CT» «q*
0~ CN'q'
40 Tf
LD CNm
«qf o- m
ro 'q* ro
00
P p QJ QJ 4-) 4-)m m P P
73QJ
g•H4JCoo
r—ICN
•9Eh
8 °
8 °
8 °
8 8
§ §
§ §
§ §
8 8
8 8
8 8
8 8
8 8
8 8
O O O O Oo
o o o o oo o 40
o o
8
§
8 8
O O O Oo o
o o o o
§ 8 °
O O Oo
o
o o Q o o in O O
rP o <M o o en 00 o
o- o oCN 40
O M" CN o (33 ro o
CN O O O O 03 O O
LDCN
o o
° §
o O
O O
O O
O O
O O
O O
O o
o o
o LD CN
O O O
o o o
o CN o
o o o
o o
o o
o o
o o
r- O
O
O
O
ror-
r-4 QJ Q) pu UJ G >1cq QJ cq QJ e uQJ QJ P UJ Xi 44G G UJ UJ >1 cq e u cq■P 0 G cq cq X G 0 .G•P 0 QJ ip 1—1 0 ■p 44 0 4 • p
UJ 44 • p UJ 0 A s e u p 73G ü 44 0 QJ p X P cq >1 p 0•P P ü U UJ 73 0 cq QJ p UJ UJfd 73 P P ip cq >1 JP ü 73 44 0P 0 73 rH G 0 73 JP p QJ ■p E
4 J P 0 o 0 •P •P cq 73 QJ E x: 0W Û4 P ■P G UJ 73 ü G 0 44 P QJtu G G 44 0 0 QJ QJ cq P ip U)
0) 0 0 cq •P P Q) 73 G Æ ip cqUJ CD •P 44 4J ü G •P eu G Gcq UJ G 44 G U cq •P QJ JP Q) 0 QJ p •P •Pip cq 0 cq QJ P 1—1 G G P UJ 4 4 iH ip 0 44cq 73 •P 73 E 73 cq •P •P •P cq e u 0 44 cq
4 4 •P 44 • P P 0 cr> On UJ G Q) >1 73 CD QJ »Hcq X ü X QJ P P >1 P P p G C N U QJu o < O k eu C û cq r—1 0 P 44 •P en < O

135
73
•5-pc8OO<ur—jAg
OO -q'
u o
Op OO
03 OJ U3
00 03
O ' CN
CD «q
U O O Om
o-m
m 'd' 03
CN m
00
P P Q) OJ •p -p co cn D GrH iHU ü
c•H
enGp73p4-1CD
8
8P
uo00
§
oO'
03
o
5 "2 ^13 en
O O O
8 8
O u o
O O
O O J
O O u o
O' pCD 00
8 8 8
O 03
œ
<33 o oro
U3O-
O O O O O
8 8 8 8t—i P P I—1
U 3 O u o r o 00
o <N| +O
O c n I + O
O C D | +S
8 r o O O r oI—I 03
CD 00 CN (30 0003 03 (33 <33 CJ3
R 8 8 8P P P
p r " r or o C D r o
8
8
8
03 8 8 8
p ro p p p03 uo C'' (53 cy>
C'' P r ~e n «q" (53 p (33
O O O 00 O
uo p 00C N C D p
§
8
(D G1—1 f-1 CD en P0 0 en CD 0 1—t4-) 44 0 en P rcJ•H P G 0 XP 73en JP s p p (530 P <C3 u p >1G 0 43 G (DH CD « CD S
00
co
co I
CN 1 +
001 +
O col + o
col
CN
73CD44en(Dp
ücn PG Ap 075 Pp (D44 73CD G
73
P • # 1—1JP 073 rH PP CD 0 0 P< en p p P0 (D 4:u 73 U 44+ G P >1 >1rH U rH pXI ü C C5 w

o CO + + + + + + + +
03 CO > + + + + + 4- +
CN CN + + + + + + +
136.
+ + + + + + + + + + 4-
4- 4 - 4 - 4 -
00 00 4 - 4 - 4 - -t- -|- -f -f- -H -4 4- 4-
r' 00 e> 4- 4- -4 -4 4 - 4 - 4 - 4 - 4 - 4 -
CO CN 4 4 4 4 4 4 4
CD CO 4 4 4 4 4 4 4 4 4 4
'q* CO 4 4 4 4 4 4 4 4 4 4 > > 4 4
P CO 4 4 4 4 4 4 4 4 4
4 4 4 4 4 4 4 > 4 4 4.. pP CD CD P P cn cn CDG 4-> 1—4u cn•pGpPCD
CD CDcn cn0 0c c CD CD 1—1•p •p cn cn 0p JP CD 0 0 pG G cn 1—4 1—4 •pP P 0 c< < XI X X 01 1 •p 1 1 73Û XI « Û XI <
CD
cn0f—IS’1fHI’9E
CDCD CD cn CD CDcn cn 0 cn en CD r-H 1— 1 1—1 1— 10 0 P 0 0 en 0 0 0 04J u Ü c JP 0 41 4> 41 41o G G G p G •P •P •P •PG rH P G 0 Ë O en G XIrH O k S CD G rH 0 G pG 1 1 i 1 rG G G G 0Ü Q Q XI « Q H S CD

137
+ + + + + + + + +
+ + + + + + + + + + + + +
03 > > > + +
CO + + + > + + + + +
+ + + + + + + + 4
4 4 4 4 4
4 4 4 > 4 4 4 4 4 4 4
«q* > 4 4 4 4 > > 4
4 4 4
4 4 4 > 4 > > 4 4 > >
9g•HPC8CN
CDP9Eh
QJ QJ QJ73 73 GP P P P(D W U)P 0 0U) G 0 enG G G 0rH G P uu S O G
rHQ Q urH P rHX >1 >1X Æ PP P QJQ) Q) U2 E G3 3 2
GP
i
QJenG 0G P G P GP P p JP enP G U 0 0G ü p P 4JA en 1—1 1—l PP QJ G G G< C CD U S
GG en G
G en G 0 enen 0 en G 0
G 0 P 0 P Pen P G P G P p0 XI X! G P P N4J •H U X: P G Gy rH U G G Di1 rHG G G p G GXI S CD Eh H Q S

138.
+ + + + + + + +
03 + +
G.3PGtnGGtnG•HGÜwI—I t— I <
co > > > > >
4 4
> 4 4 4
GrHGP>
lO 4 + 4
4 >
4 + 4
4 >
4 4
G>P+>Penaü)GPGPPen
> > > > + > +
73GBppG8CM
Gr-H
GE"
GG rHÆ 03 0U 0 pP U pG >. rH+1 rH >1CO ü X
p G G rH rHX en en 0 0 G0 0 G 0 G G p p P•H G en P en en p p Gp G Q G 0 0 X) X GG p 03 ü u G G 0G G G G G P P UU Eh XI EH fU k < < G
Q 1 1 1 1 1 Pco. Q Q Q XI Q XI U
G
G8GrH03OpX
G+)Soug03
ops,
G>P+1PenOeu
enGpGpPen
àP4

139.
numerical analysis package (Wishart, 1978) was used to sort the
binary data. Sorting was by the Jaccard (S^). Gower (S^); or
simple matching (S ^ ) similarity coefficients, clustering was by
single complete or average, (UPGMA) linkage. The dendrogram
(Fig. 42) supported the initial conclusion that 12 distinct clusters
occurred, these joined at a level of similarity of 59.5% (69.5%S).
Three hundred pure cultures of presumptive Enterobacteriaceae
were assembled from Violet Red Bile Glucose agar (VRBG) inoculated
with dilutions of sulphited or unsulphited sausages or ingredients
used in their manufacture (Table 22). The following seven stock
cultures were added to this collection: Citrohacter frenndii,
ATCC 11102; Citrohacter intermedius, NCIB 4143; Enterohacter
cloacae, ATCC 13047; Escherichia coli, ATCC 25922; Hafnia alvei,
ATCC 25927; Serratia liguefaciens, NCIB 9321; Serratia marcescens
ATCC 13880. As a result of initial characterization by traditional
methods, (Edwards and Ewing, 1972) as well as with the API 20 E
and 50 CHE schemes, 15 strains could not be assigned to a cluster
nor a genus. The remainder, which fell into 12 clusters, were
identified with Hafnia alvei (76 strains); Enterohacter agglomerans
(49 strains); Enterohacter cloacae (42 strains). Citrohacter
freundii (37 strains); Escherichia coli (34 strains); Serratia
liguefaciens (32 strains); Serratia marcescens, (14 strains)j
Yersinia spp. (21 strains) or Klebsiella spp. (5 strains).
Analysis of the results indicated appreciable differences
between the types of Enterobacteriaceae which occurred in the
sulphited or unsulphited product (Table 22). Thus, Escherichia

140,
w SVc<u >1T)01U tt)O' CÛCu •Hu
K (U cO O ' Iflo (0 tuw in h3o(0t/3
1(d(N O
CN CO O ^
1 I I n
0 ^ 0 0 0 0 0 0
O - l O O O O O O O ' ^ O O
O t N O O O « N O O n c M f s i O
i-l O tM
—< 0 0 0 0 0 0 0 c s r ^ o c N
O r o o o O - H O O C M O t N O
mc: 03 1H <i) m 73 e73 M c Ü 0q 0 H) 733 ,7: u 0 &)Q) tn Cn0 0) 03 CJ 73u P 3 ü
a eu tj! k k ka a m L, 03 03U) in <l)•c CJ ü<0 IQ ü Ü 7J 73 73 73S •H Jû rQc: c k A 0 0 tnD 0 73 73 k k 3w tn •C P k, k 03 03 73k Ü k k -U •U km m y> H3 03 C 3 03X ce o CO Co te te
T3<ucO'

141.
coli, Enterohacter cloacae and Yersinia spp. were isolated frequently
from the former, whereas Enterohacter agglomerans and one biotype of
Hafnia alvei from the latter type. These findings contrast with
those obtained in a detailed analysis of the pseudomonad flora
which indicated that no such selection occurred, (pp. 258 - 293) .
Lactohacillus
Of the 92 cultures isolated from sulphited or unsulphited
sausage on MRS agar, all grew on a modified (Dowdell and Board,
1968) Keddie's medium (1951), were Gram-positive catalase-negative
and occurred typically as bacilli in pairs or chains. Furthermore
all strains produced acid in Whittenbury's (1963) medium although
strains L64 and L65 grew poorly. On the basis of these and
other tests (Table 23), 90 strains were assigned to the homo-
fermentative and 2 strains (L64 and L65) to the hetero-fermentative
group (Sharpe, 1979). Of the homofermentative group, 10
(LlO, 37, 46, 55, 72, 73, 79-82) were assigned to the thermobacterium
sub-group lA, and 80 to the streptobacterium sub-group IB (Table 24).
The four major fermentation profiles which were typical of 81 of
the strains are summarized in Fig. 43. The majority of the homo-
fermentative streptobacteria were identified with Lactohacillus
casei var. casei (43 strains Fig. 43a; 28 strains. Fig. 43b), 4
minor profiles described the remaining 9 strains (L2, 14 ,22, 28,
33, 47, 69, 78, 101) which were left unassigned to species. Two
distinct profiles were typical of the 10 strains of homofermentative
thermobacteria, 6 strains (L72, 73, 79-82) were assigned to
Lactohacillus salivarius (Fig. 43c) and 4 strains (LlO, 37, 45,

142.
CMena CN
r"
CM00I
cn
1—4i-q0)
T)OUc0
•H4JUQ)1-41-40ÇJ
LT)
IT)
VOminVO
CM 00 r' Im cn
m
CM
inin
VO
ro
CM00I
cn
m
CM
mVO
inm
VO
3 mv4 r-TTVO
Q)
WPto
E2iw
■nQ)
f-40CO•H
•HU
IurO
COCD-Pm1-40CO•H
<4-10
54
1
om
CNcn VO § g CM00 o CO
00
<440toOJp
•MV4PPtd
tdÜ•HCnOr-4o•HCO
cu
nCM
E • •CO 73E p CU
CO r-4 Ep 0 U1—1 X3 00 fd PA P<d o 73P E Pcu U
p E (U fdfd >
cu p u:? > p p0 p fd Pu p P ocn fd c fdu u U P (U r400 0 0 C + 1P in r~ in 0) 144
r-4 M E (U 0 PI Q>1 V4 pP cu 0 pp p p cur-4 0 cu Ep E p 0XJ 0 cu to< m H
Q)PfdC0UPrHCn
S2P
TOQ)
OP
CDT3•HX•S73POA13U
c•Hr4PuCOcufdp0
CO•HCO>1rH21

143
730)3.5VcoumCN
•§
CNenPI1
d
0)730U
C0 m p•H VO 00PU(U 'cf or-4 VD 00#— I P P0U
eup0pf—4DU
U)<üPfOr-4oenp
p CN 00 (N0 00
p03JQ
Z
03PPCPenPfd
g0p ^p fd
0373 503
P Ew P 103 0 \
P + 1 +
fdpC0

144
03
CO
sCO
x:CO03PP§PP
'Spfdr —4OCOp
pCJfdPOPufd
poco
pp
sppppc037373CfdCO03Üpp0no
CM
Pfd
p P03 0)CO COfdu u
CO . . CO CO 73 733 p p 3 3 03 030 M fd fd P 0 G GP M > > P P cn Cn
P Id P P P73 0 p > 03 CO CO03 fq 03 03 p > CO COC3 X3 CO CO p P fd fdCr> 0 fd fd fd 03 G G
P p Cj 0 CO X3 D DCO Üto m< P
fd fd fdp p fd fd pp P p p p03 03 P p 03p P 03 03 pu U p P U
04 fd fd C3 C3 fdP X3 X3 fd fd XI0 0 0 X> XI 0 1
•• P P P 0 0 pX: cn 04 04 E E 04P 03 03 P p 03P X3 P P 03 03 P5 p P P X: X3 Pw CO CO E h E h CO
73 03 P P P P C 03
73 AH 0OPÜ
COPPfd 03 p p C P 03 P G 0 P P 03 Dj IP
03S’CO
sCO
§pp
rH0CO
p0p
EI
*0 0 0 0 0Ë E E E S p0 0 0 0 0 0353 53 53 53 53 53
(Tf I—I
"vrCN
cn'sT
CD
COCN
LD
VO
CO03 03 s
P C P C•P ^ P ?P O P O0 x3 o x:P to P CO
fd X) u 73p
Pp
Pm m m n P 0 P 0
«d* Tf 0 G 0 GG Gp cd P fd
Cn cn Cn Cn S p E pp p p P fd fdk k tP k 73 pH 73
in o
cn
03
•S
«
Iip
I
03
P
1P
203P0353

14'
55) to Lactobacillus helveticus (Fig. 43d). One carbohydrate fer
mentation profile was typical of the two heterofermentative
lactobacilli (L54 and L55).
The presence of sulphite preservative did not influence
significantly the range of Lactobacillus spp. isolated from
sausage (Table 24).
Streptococcus
All of the 48 isolates from KAA agar - 24 strains from sul-
phited sausages (SI - 824) and 24 from unsulphited sausages
(S25 - S48) - produced visible growth on modified (Dowdell and
Board, 1968) Keddie's (1951) medium, gave a positive reaction
in the Gram stain test, a negative catalase reaction and were
typically arranged as single, paired or chains of cocci. All
strains grew at 10 , 15°, 37° or 45°C and were homofermentative.
Furthermore, all initiated growth at pH 9.6 and in 6.5% NaCl and
were tolerant of 40% bile. On the basis of these and other tests
(Table 25) they were assigned to the "faecal" group (Sharpe,
1979) of streptococci. Thirty-four strains were identified
tentatively with Streptococcus faecalis and 14 with Streptococcus
faecium . Results from a screen of oxidative and fermentative
breakdown of sugars (Figs 44 , 45) suggested however, that there
were 3 groups. Thus 25 isolates could be identified with Strepto
coccus faecalis, 10 with Streptococcus faecium and 13 with an
'intermediate biotype' which was left unassigned (Figs 44, 45).
The presence of sulphite preservative did not appear to influence

146.
03 CP (0 t/3%wIpp%p(T3rHs•p•puu0uop04%pwp0U303PPP•PPPP(UiHs•pCPoI—Io■p>1r CO,
LDCM
•§Eh
CO
CO
03730U
GO•PPU030Ü%PPrHpu
COG
• P(dpPCO
p0p03
1
V£>CO
OCMIdpp03pp0GO•PP
B7303D3
inCP>CO
00CO
03CM
inCMCOCM
VÛpinPVDinCMCO
1—1
(TiCO
00CO
<73CM
inCM 0 0
COCM 'd'1—1 VO 'vr
'v r 'd’VOP m P
'd'in r-'P 'VJ' O CO
VO ■'d'CO CO COin CM
CMCM CM VO COcn HO* cn cn
03p•psrHrH03P
•PCOCOIdPâ
+
G•p
I6
VOm
PIdS’730OrHX)§CO•prH0§5P003aEh
Omc/3
>1Pcn7303 G G GG 0 0 Id
§ a ca a
CO
Idp03CO-ppcId
73 03tH X :03 p"p 0p03 x:U pG -pId
730 GX : 0
04 p •Pp 03 P0 E UP Id<73 Q 03cn p1—1 Û4Id H P COu 0 CO•p w P 0<73 Ü p0 u uop03CO

147
significantly the composition of the Streptococcus microflora and
isolates identified with the different species or biotypes noted
above originated from both sulphited and unsulphited sausage
(Table 26).
Yeasts
Of the eighty-nine yeasts isolated from ingredients of pork
sausage and from the unsulphited or sulphited sausage which had
been stored for upwards of 7 d at 4°, 10° or 22°C, 67 were
non-pigmented (white, cream or buff) and 13 pigmented strains
(pink or orange). With subculture on malt extract agar, 48
were mucoid, 35 were smooth and 6 were rough in texture (Table 27).
All strains showed multipolar budding from circular or oval cells,
a minority possessed arthrospores, a pseudo- or true mycelium
and none produced chlamydospores (Table 27). On the basis of
morphological and biochemical tests the strains were assigned to
8 groups. The majority (53%) were assigned to the genus Candida
or Torulopsis, 25% to the genus Debaryomyces/15% to the genus
Rhodotorula and 7% to the genus Trichosporon (Table 28). This
survey of a limited number of yeasts suggested that sulphite did
not influence significantly the composition of the yeast flora in
sausages (Table 27).
Culture studies
The observations discussed on pp. 1 2 5 - 1 32 identified the
bacteriostatic role of free sulphite in sausages. This led me
to the hypothesis that the contribution or otherwise of a particular

148
euCP(t!Wwsopp■puü0ü
sCueuppwp0co■ppfdü•pp■ppceu73H
noCN
PedE-f
eu » *73 p 000 Cu r~' enu eu ro
eu enG X LT) CN0 eu m•p LDp eu CNü > coeu •prH en m PrH P CN0 rH vOu ü 1— 1 vO 1—l
G peu •P enP cr> - pp 00 " «vrP sT LDp 1 - lp rH 1—1 1—1 CNU e/3 53 'd' in
eu
coPfden
enG CN•p ofd 53pP 1enp0
euC en■p fdCn. en•P PP fdO en
enC•pfdpPenp0PeuI5-p
'S•pp•ppceu73
IPÜ
inCN
en■ppSeu'Pen3B8Op%P53
00
73
•PenenfdGD
uo
O
1•p
8'Pen388O
253
LT)
"d*enen■pkeueu03

149
(Uen(dcnDfden
73cfdcnPc03
■p7303penc
•p
03< rfdenDfden
0P
P
enC
•pfdppen
Penfd03>1
pO
en03PPP•ppPPfd
fdo■p
r—j028
73Cfd
en03UPPOen
oCN
E-f
CPo
(-4
&8U
■p
§■uenOPeu■pS
•Hr4
03 03G üP >1Eh E
E1 G0 •P73 rHG 0303 Üen >1a E
en03P
RenOP
PPpC
en03P
&073>1
U
03
&x :en
*&Or-4OU 03SEh
CNO
03 en en I fden CN 3 O fd en en +
enPC03•P73
g:enG
enp G03 -pXJ fde PG p PZ O en
aP0po
o o o o o
o o o V) c^ CNCN i— I
O
o o o o o
> > >O o o\P i-H 1— 1 r H r H > 1— 1 >U u u u U o u o
CQS S "u O[2 s: "-v oCu
PO cn cn\ \ \s s s c n s s s c n
CN m cTi O
L n v x v D O f ^ c N i r i o
O 'P LfO p o f^
O c N C T v m m c N P C T f p CN CN p CX3
c^
C D
p CN f ^ V LD X ) 00
pfdrHGU rHP rd•P >U o
rH >U o
0303 E enp fd P X G• p 03 P G f3X p G • P p12 u CQ a o
S U CÛ Pu o
X pen oo e PO en
s oo en
03POGpootu

cnPG03
•p7303Pcn 73G 01-P P
cn03 0)cn pfdcn PG Qfd ccn
73G'fd
0303 >cn •Prd PV) •PG cnfd 0V) aE cnO GP •pP fd
P73 P03 cnpfd pr—I 0ocn p•H 03
XcnPcn Gfd03 4 ->1
p 03o >
■Pcn P03 •PP cnG 0
X a•PP cnP GP •pfd fd
p1—1 Pfd cnu•p PC71 P0 fdp0 11•pcn +ÏPXa
00
CMp
LO If)
'q' CTi CM
m CM CM
CM O
cnC•HfdpPcnp0pa 03II
I + + + + + I + + I + I + I I + + + + + 0 0
I + i P P 0 0 " ; f G 3 I O OM I I I O + + + + O O J C M P i—I p P P p
l + l + l i l t l l l l l l + t l l l i m r o
I + + + I I I I I l + l + i I I l + l l O O O
1 + 4 - 1 I + I + + I + I + I I I I I I l O O O
I + + + + + I 1 + I + + + + + + + I + + 0 0
I + + + + + + + + I + + + + + + + + + + 0 0 0
+ + + + I 1 + I + + + + + + + I + + O r -
03Ps0UGr—I CDtn COi aO I
P 0 03 P U 03 >1 X
IU O U CM
03p cn I—I I—I0 p O 0 0
03 P O P P PW P P U -H -P0 G -P fd cn X)P O P P O pÎP 73 >1 fd C OX C X O H CO
a•pE
03 fd 73 cn •p 0 cn Ü 0 G0 P G U)I—iCJ Q 031 cn
Q P 0I ÏP -p
P P XI^ S . 3P fd P 03 03S Z CJ
03 03 CO cns scj P fd fd X S
Go•pp
5G
ËP03 03
03 cn 03 Pcn 0 cn0 P 013 'S '.s
Ü X 03 P U 03 P P fd P 03 fd CO Eh s CJ
pH
EHcn 73 cn ofd p
a03 03 cn p oG
03 fd cn P fd P 03 •p p Z D
150.
fdpGP0P0 7573 P0 XX GOO P
fdPGP cn0 •pP G0 •p73 P0 GX PDO Cn
fd73•p73GfdO
cn0373•pfd 073 G•P 7573 PG G)75 03CJ N
7573 75•p cn73 0G Cn75 GCJ pcn03ÜE •p0 •pG) GP 0375 cnX G03 75a X
cn•pcna Id0 73p •PG 73p Gp 75Eh ÜG0P0a Ecn G0 03X GÜ fd•p Pp GEh Ü
cn•pG
•p « P
P G
S
7303GCn•Pcncnfd
PCJ

151
group of organisms to the microbial association was perhaps associated
with their relative tolerance of sulphite. This hypothesis was
tested with isolates from sulphited and unsulphited sausages.
Batch culture
Preliminary studies were done with actively-growing static
or agitated cultures in 90 ml of a chemically-defined medium
(pH 7.0) in 250 ml Erlenmeyer flasks to which sulphite was added.
The concentration of sulphite which inhibited growth was monitored
by viable counts. Thus, for example, a concentration of 250 yg
free sulphite/ml MGGM caused growth stasis of Hafnia alvei straino
E 34 at pH 7.0 and 30 C whereas 125 yg free sulphite/ml reduced
the rate and extent of growth; 500 yg free sulphite/ml resulted in
death (Fig. 46).
The rate of growth of the six strains of yeasts and 8 out of
the 9 strains of Brochothrix thermosphacta was not influenced by
ca. 500 yg free sulphite/ml at 20°C (Fig. 47). The growth rate
of the exceptional strain of Brochothrix thermosphacta was retarded
by 470 yg/ml. A lower range of sulphite concentration (255 - 415 yg
sulphite/ml) inhibited
the growth of Gram-negative bacteria and in some instances the
concentration causing bacteriostasis was very close to that causing
death. Thus with 9 strains of Pseudomonads and 24 strains of
Enterobacteriaceae the minimum inhibitory concentrations (MIC's)
ranged from 160 - 330 and 15 - 270 yg free sulphite/ml respectively
(Fig. 47), within the latter family some members - Enterobacter

152
agglomerans (MIC 50 - 100 yg/ml), Yersinia enterocolitica (MIC
50 - 100 yg/ml) and Salmonella spp. (MIC 25 - 125 yg/ml) were part
icularly susceptible, whereas others - Hafnia alvei (MIC 210 - 265
yg/ml), Serratia liquefaciens (MIC 200 - 250 yg/ml) - were less
susceptible to free sulphite.
Pure cultures of dominant members of the microbial association
of sulphited sausage were substantially more tolerant of the preser
vative than were those of organisms (e.g. Pseudomonas spp.,
Enterobacteriaceae) which did not contribute significantly to
the association. As such experiments were done with "physiologically-
fit" micro-organisms at temperatures which allowed good growth,
the situation is not analogous to that in a freshly prepared,
chilled sausage. Thus these preliminary experiments with batch
cultures were extended to studies of the influence of high initial
concentrations of free or bound sulphite at temperatures of 4° and
12°C on Salmonella^ these organisms of public health significance
being chosen because of their apparent sensitivity to low concen
trations of free sulphite.
The 6 isolates of Salmonella did not grow in modified glucose
glutamate medium (MGGM) at 4°C but did so at 12°C. As the response
of all the 6 strains to sulphite was similar at both temperatures
(Fig. 48), only the results obtained with Salmonella virchow
rif^ - the serotype used in sausage-seeding experiments (pp. 157
- 164) - are described.
When 900 - 1000 yg sulphite was added per ml of sterile MGGM,

153
ca. 200 - 300 yg were bound reversibly within minutes; inoculation
of the medium with Salmonella did not cause appreciable binding of
the residual free sulphite (Fig. 49). The latter was easily bound
by the addition of an excess of acetaldehyde without influencing
appreciably the absolute level of total sulphite (Fig. 49). The
loss of free and total sulphite from uninoculated MGGM was faster
and occurred to a greater extent at 12°C (Fig. 50) than at 4°C
(Fig. 49). Moreover, the transfer after 7 days of flasks ofo osulphited medium from 4 C to 12 C resulted in a rate of loss of
free and total sulphite that was greater than that occurring in
flasks stored only at 4°c (Fig. 51). Actively growing or
quiescent Salmonella did not influence substantially the rate
or extent of loss of free or total sulphite (Figs. 49 - 51).
Sulphite protected to a large extent all the strains of
Salmonella inoculated in MGGM and stored at 4°C from death.
There was a gradual decline only - ca. 10^ c.f.u./ml to ca.
10^ c.f.u./ml in 20 days - in the size of their populations
(Fig. 48). In the absence of sulphite, the populations
of all the strains of Salmonella declined at a significantly
faster rate, those of Salmonella anatum, goerlitz and kedougou
from ca. 10^ c.f.u./ml to ca. 10^ c.f.u./ml in 20 days and those
of Salmonella hadarftyphimurium nal.^ and virchow (Fig. 52) from
ca. 10^ c.f.u./ml to ca. 10^ c.f.u./ml in 3 - 6 days (Fig. 48).
The addition of sulphite to slowly growing cultures of
Salmonella at 12°C also caused rapid decline in the number of3 4viable organisms from ca. 10 - 10 c.f.u./ml to numbers below

154.
the limit for detection (ca. 10^ c.f.u./ml) by days 4 - 6 (Fig.
53). Sulphite bound by acetaldehyde reduced both the rate and
extent of growth as compared with the unsulphited cultures. It
was notable, however, that large populations (ca. 10^ - 10^ c.f.u./ml)
were attained after 7 - 1 3 days (Fig. 53).
When cultures of Salmonella stored at 4* c for 7 days were
transferred to 12°C, the following observations were made:
(1) Salmonella in sulphite-free MGGM initiated growth; (2) the
protective effect of sulphite was negated and rapid death ensued,
and (3) the sulphite-acetaldehyde complex did not prevent multi
plication of Salmonella, (Fig. 54).
The observations made with batch cultures supported the
hypothesis that the sulphite tolerance of members of the microbial
association was such that this property can be considered to be
a major cause of the selection of the association described on
pages 125 - 151. Moreover, the evidence supports the conclusion
that sulphite , in the free but not bound form, acts mainly as a
bacteriostatic agent at neutral pH. The results obtained with
Salmonella are of particular interest because they suggest a
protective role for sulphite with susceptible organisms stored
at temperatures preventing growth.
Turbidometer culture
As the batch culture system relied on determination of viable
counts to index the action of sulphite there was no opportunity for

155
critical assay of sulphite tolerance particularly as the range of
tolerance was extensive. Thus a novel assay system was developed
(Fig. 9). The preservative was added incrementally to a continuously
monitored culture in the exponential phase of growth. When the
trace of optical density plotted on the chart recorder indicated
that growth had been arrested, the concentration of free sulphite
in the medium was determined immediately. Moreover, the method
allowed constant adjustment to the concentration of free sulphite,
in the culture medium thereby compensating for irretriev able - by
oxidation - or reversible - by binding - loss of active sulphite.
This dynamic approach to the assay of sulphite sensitivity confirmed
the preliminary results obtained with batch culture experiments.
Thus stasis was not induced in any of the 8 strains of yeast
or 2 strains of Brochothrix thermosphacta by a concentration of
500 yg free sulphite/ml whereas 125 - 270 yg free sulphite/ml
inhibited the growth of 2 strains of Pseudomonas fragi and 1
strain of Pseudomonas fluorescens (Fig. 55). With the exception
of members of the genus Salmonella, growth of representative
Enterobacteriaceae was suppressed by 50 - 270 yg free sulphite/ml.
A lower range (15 - 109 yg free sulphite/ml) inhibited Salmonella
obtained from culture collections (e.g. Salmonella virchow rif.^;
Fig. 56) or from sulphited sausage (e.g. Salmonella derby; Fig 57).
It needs to be stressed that growth stasis as indicated by
turbidometry was confirmed by viable counts. Sulphite-induced
stasis of the Salmonella cultures persisted for ca. 20 h at 30°C
after which time the organisms died (Figs. 56, 57). The addition

156.
of a binding (acetaldehyde) or neutralizing (hydrogen peroxide)
agent at this time neither released Salmonella from growth inhi
bition nor prevented their death (Figs. 56, 57). With the notable
exception of Enterobacter agglomerans, an organism which responded
to sulphite in the manner described above, other members of the
Enterobacteriaceae were inhibited but not killed by sulphite in
the course of the experiments. Indeed the addition of a sulphite-
binding or neutralizing agent led to the growth of organisms
which had been inhibited by sulphite for 20 h at 30°C (Fig, 58).
The results obtained from batch and turbidometer culture
permitted a ccmparison of the sulphite-sensitivity of common
contaminants of commercial sausage (Fig. 55) tested under aerobic
conditions in buffered, defined media at pH 7.0 and at near
optimum temperatures for growth. Further studies showed that
the actual range of sulphite concentration causing growth
inhibition but not the rank order of the various ranges discussed
above was influenced by incubation temperature and redox
potential. Thus an organisms'tolerance of free sulphite diminished
as the incubation temperature moved away fran the optimum for
growth. Free sulphite was generally more effective under anaerobic
conditions although 4 strains of Salmonella - anatum, derby,
kedougou and virchow - were notably tolerant. Indeed, in the
absence of oxygen, high concentrations (> 250 yg free sulphite/ml)
were required to inhibit growth at pH 7.0 and 30°C.
There was no demonstrable difference noted in trials which
compared the sulphite sensitivity of isolates grown in chemically-

157
defined or complex media, even though the latter had a much larger
potential for binding sulphite.
Fate of a selected pathogen in sausage
Because of the particular importance of Salmonella in food-
borne disease and the good public health record of British fresh
sausages with respect to this group of organisms as well as their
apparent marked sensitivity to free sulphite, a series of experi
ments were done to establish their fate in sausage. The low and
unpredictable levels of natural contamination of sausage with
Salmonella (pp. 230 - 257) meant that samples taken from a
factory could not be used in studies of the fate of these
organisms during storage. Thus sausage meat was inoculated with
low numbers (ca. 10^/g) of Salmonella and other Enterobacteriaceae
which had been isolated from sausage,grew on PCA, MacConkey agar
or in TSB whereas, with the exception of Salmonella typhimurium
340 nal^, only the former grew (Table 29) on PCA or DCA suppl
emented with rifampicin (50 yg/ml). In contrast strains of
Salmonella typhimurium 340 nal^, Enterobacter agglomerans and
Citrobacter freundii only grew in TSB supplemented with 30 yg/ml
nalidixic acid (Table 29). Moreover, of the Salmonella serotypes
studied, goerlitz was the only one affected adversely by the
rifampicin-supplemented DCA medium (Table 30a.) Preliminary
experiments showed also that yeasts in sausage grew on rifampicin-
supplemented TSA but not on rifampicin supplemented DCA. Thus
the latter medium was selected for use in this study.

158.
03(d03üfd•H
4-3UfdX203XgX0co•H1dr—40cn•rH
G0•rH7303E&>OU03 3404 O03
RCL)GXUHGH
(TiCN
•§Eh
S
1• H7 303E>1ïOU03G
GO
X
22U)
<R
H
enI•HGfdS'o
+1
•KenGGH •K
•G•G
0 73Ht M G"M tn GM 030 G G
* Ü Oh•H G0) G 03 G:> •H X 03M X ü XG 0 G 0•M X G<d G 0 X'M 03 G 0G X 03 GX Ü X Xfd 03 G •G£G kl kl O
sG
• G
iIX
(dM
03GM
to
N GX 0• G 0 & i EX X G G GG 0 0 G X03 G 7 3 r r 73 GP • G 03 w G Gtn X : X G
X
IenZ
XI34en
X
Een r H
Eo enin X
Od iH f
03 • G Een uG • G tjf 73en A • GG UG o Gen X m• G UX G - Gen d XG X • G • GG X u 73X - G • G - G2 D u i HX Ë fden 73 G d
• G G XX X • G X• G d G XG G • Gm E X 2G X73 r H • G 7303 A 3 GX a X
■G d 73 dX en G GQ -P EX G d GG G G Xen d en euG c • G G euG • G G 1—1 d\ G G G A en73 G X X CuG X en G d XX en G en X
• G X X 0X X d • G G GA d G U G XX G X C3'G X en G G Gen en • G X >1• G en G X 0S en G i H d en
0 03 G 0 dG G X 0 GX d u o dd • G >1 073 • G ü X G X
03 X • G 0 X û)X • G A en G > 1G 73 E G 1—1 G1-4 • G G Q PU Eh0 X Xen G • G OP Z
H d G 3 < CQg u CD•K +-txn Q cu EH

159
Table 30a.Influence of recovery medium on quantitative isolation
of Salmonella
Salmonella :TSAR
Isolation medium TSA DCAR
goerlitz 431* 472 216
virchow 230 228 232
kedougou 379 411 360
"E4" 301 337 275
hadar 459 471 460
anatum 226 234 206
* Arithmetic mean of 5 replicate counts at a dilution level of -510 in TSB
TSAR Tryptone soya agar supplemented with rifampicin (50 yg/ml)
DCAR Desoxycholate citrate agar supplemented with rifampicin
(50 yg/ml)

160,
When diluted mixtures of cultures of Salmonella and other
Enterobacteriaceae were incubated on rifampicin-supplemented DCA,
significant (p = 0.05) diminution in the numbers of Salmonella
anatum in the presence of Escherichia coli or Enterobacter
agglomerans, and a group E 4 Salmonella sp. in the presence of
Escherichia coli was noted (Tables 3ob 31). Conversely, signi
ficantly greater (p = 0.05) recovery of Salmonella hadar in the
presence of either Enterobacter agglomerans , Citrobacter freundii
or Hafnia alvei on rifampicin-supplemented DCA, was observed
(Table 3ob 31). As Salmonella virchow was not influenced
significantly by the type of recovery medium or competing organism,
it was chosen for further studies.
Influence of sulphite concentration and temperature of storage
As similar trends were evident in all the three major experi
ments on this topic, for purpose of brevity the results of one
particular batch only are described below. It should be stressed
also that although the unsulphited and sulphited sausage meat was
obtained from a large factory in the course of routine production,
inoculation of meat slurries with rifampicin-resistant Salmonella
was done in the laboratory.
With the exception of storage at -20°C, (Fig. 60) Entero
bacteriaceae grew at all temperatures in unpreserved sausage meat,
the rate of growth was related to temperature (Figs. 61 - 65).
The numbers of these organisms did not change in sulphited sausage
meat stored for up to 7 days at -20°, 4° (Fig. 61), 9° (Fig. 62)
or 15°C (Fig. 63). After 4 days at 20°C (Fig. 64) however, the

161
CwEhC0GXXGI
X
CDX0d0
■GXGI—I0w•G
G>•GXGX•GXCG&0w§
■G
enG010GU-genc
•GXG
§u
X0GucG3f—IXc
H
- 0oPO
•3Eh
Of-4
CD
co
G
SI•G§S'o
vû 00CMCM
ID
I\uoXCM
CMr - -
PO
Z
CMCMCM\(DXCM
POXPO
coCM
I\vOXCM
OPOCM
+O00
XPOPO
VÛXCM
CMPOCM
PO
PO
8PO
POCM
u nCMCM
CMCM
CM 4" X 'Uf un GCM 1— 1 X CM XPO CM PO CM PO CM G\ \ \ \ üun 00 CM un 'si' •G00 PO O X 00 uo r—! wCM CM c o CM PO CM A dG GG >1 GX CD 1—1 X CM G •G Gun CM CM 00 un PO un G •G EPO CM PO CM PO CM > 73 0\ \ \ \ X 0 d XX CO X uo 0 U 3 CD -GX 1— 1 un un un G G CD XCM CM CM CM r o CM d G G G 0G Mh CjG d G •Ge G G G G GVû uo uo X G G X •GCM O CD CD CM O U S X Ü -d XPO CM CM un PO CM •G Ü G ü G\ \ \ CM > \ X 7 3 G A GO un PO \ o CM G G h 0 G Gun 00 un H un UO X 0 G G •GCM X CM Z PO X ü G G -d dX G X X CJ kl
X eu •G d G GG X O Cq kl Cd< wun uo X 00 CD O1— 1 * - h - X CM PO
a-G§UGGCGG6
73>GGW3
unI
O
EG 3G X73 GG d"d GG GX XX XG Gd d
0EX XG GCD CD
CD Or—1
G N 3> X 0G •G 0 CD1— 1 X -d 3
G 0 0C G G 730 0 X G WX CDX3 G G G GX X X X XX X X X X73 G G G Gd d d dG 0 0 0 0E E S sX X X X XG G G G G
CD CD CD CDenX
un UO X 00
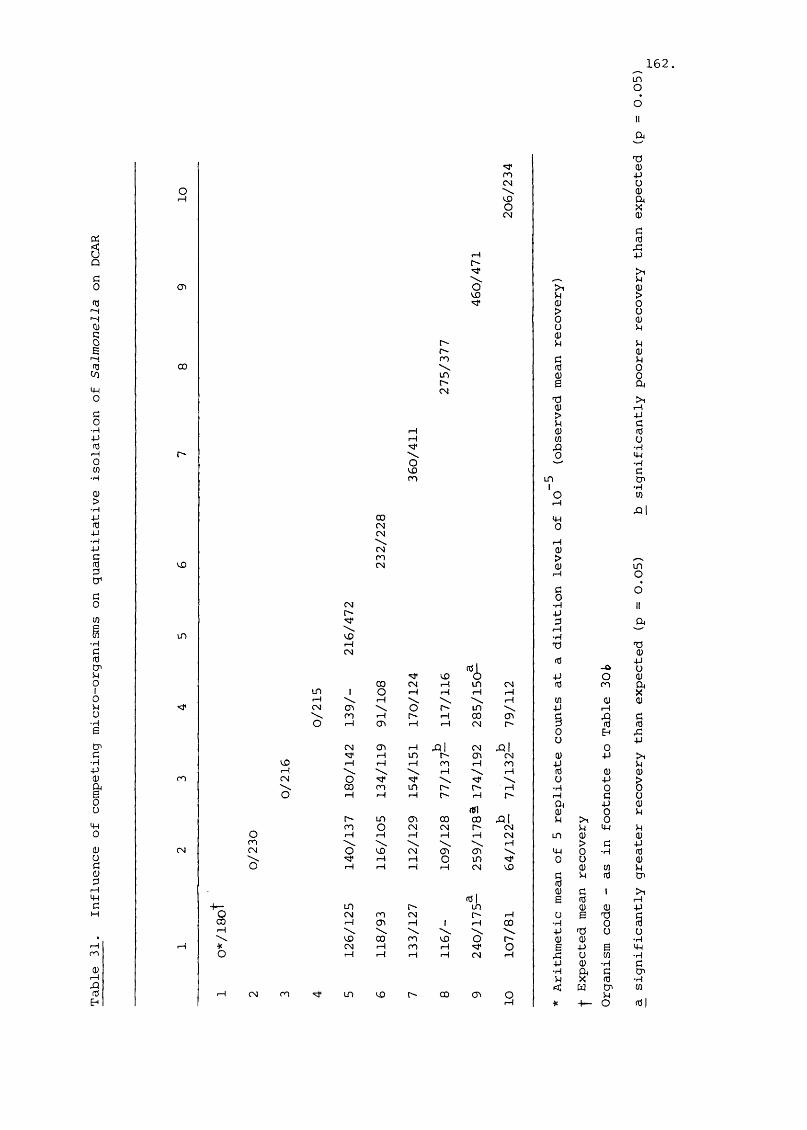
162unO
a
ic0G
XXG§S
X
54-1OGO
•HXGf—IOw
•H
■H
X•HX
§3CDCO
•H
5enG010 U U X6enc
XXG1UXoGëG3
XXd
X
G X ,
3E"
CD
00
Uû
«q*XCM
UOOCM
X•q '
UO«q*
XXX\uoXCM
\OuoX
00CMCM
XiXCM■ Xo
uoI— I CM
OXCM
S*O
X >CM G un
r-4 Od d0
CM X IIX aTT 3 A\ 1—1uo XX 73 73CM GG aGl - O uuo o a O G00 CM X un CM G X A1 O X X 1—1 X X\ X \ \ X w G GCD \ o X in \ a XX X X X 00 CD d a dr-4 CD X X CM X 3 G G
0 Eh au aCM CD X hl CM al 0"q* 1—1 un X CD CM G a >1X X X X X X a G\ X X G G GO 'cr 'cr \ <cr \ u a >00 X un X X X X 0 0X X X X X X X d üA a GGl G 0 GX un CD 00 00 al G >1 0X O CM CM X CM G a GX X X X X CM IT) G G\ \ \ ■ \ X > d ao uo CM CD CD X a 0 X G
X X o un 'd ' 0 u GX X 1—1 X CM uo G en Gd G G enGG d 1 >1G| e G rHX X un G G aCM X CM X X u E 73 d1—1 CD X 1 X 00 X 0 G\ \ \ \ \ a 73 U uUO 00 X uo o X G G XCM X X X o s a Ë aX X X X CM X a U en Xa G X d
X A d CDG X G X< w en enIT) uo X 00 CD o G
X * + — O G|
&GêoGG
36
•SaGW
3
O
XO
73GXUG§■GS£a->oüGGGGG0â
XSu
XXXdenXw

153
decline in the concentration of free sulphite to non-inhibitory
levels (ca. 50 - 60 yg free sulphite/g) was associated with a
small increase in their numbers (Fig. 66). Salmonella virchow
rif.^ present in unsulphited sausage meat stored at -20°, 4° or
9°C did not grow, it did so to a limited extent at 15°C and
extensively at 20° or 25°C (Fig. 65). Growth of this serotype
in sulphited sausage meat at 25°C or 20°C was not observed until
after incubation for 2.5 and 4 days respectively, by which times
the concentrations of free sulphite were 20 and 45 yg/g sausage
meat (Fig. 66). It was noteworthy that these values were of the
same order as those noted in the experiments with pure cultures
in broth even though there was a progressive acid drift in the
sausage meat and hence a possible accentuation of sulphite
toxicity (Fig. 67). There was a pronounced acid drift in the
pH of unsulphited but not sulphited sausage meat (Fig. 68).
With the former the data were described by a linear regression2line (coefficient of determination, r = 0.73) whereas with the
2latter by an exponential regression line (r = 0.79), see (Fig. 69).
Influence of a high initial level of microbial contamination
It was notable also that rifampicin-resistant Salmonella virchow
failed to grow at 4°, 15° or 20°C within 8 days in sulphited or
unsulphited sausage meat which had been made from meat stored
for upwards of 96 h in the factory (Fig. 70c). As would be
expected, the freshly made sausage had a very high microbial
population, dominated by Brochothrix thermosphacta and other Gram-
positive bacilli, as determined by microscopical examination of

164.
Gram stained films made from colonies on Plate Count agar (Fig. 70a).
The extent of growth of the dominant microbial group as well as
that of the Enterobacteriaceae was curtailed (Fig. 70b). The last
mentioned group, although present initially in high numbers did
not attain the large populations in unsulphited sausage meat which
would have been predicted from the analysis of sausage containing
normal levels of contaminants (pp. 125 - 130 ) . Thus the extent of
the initial contamination as well as the free sulphite content
influences not only the growth of the Enterobacteriaceae, but also
salmonellae in sausage meat.

165
(UmwDcox:cod)u
CW0C0•H4J(T3C
•rHSrüVC8
AOMÜ•HeUHOQ)>(D
r—4
fO•HJJ■rHC
(U4JC0(UupH-l14-1ëg0
03(+H0euCJC=14HC
en>1tdT3•HH
kUOen
rgenua54enS’"Seuc
gco
'gk5iwppp6eu
en%en
(+H0eneu43u-PPCQ
•H
•rHCoro QJc ene (V
c/1<TJ ZDroC c/)Owen
O d«H—
QJ w>QJ c
enoro
c
OCN
eukPen•rHQh I
a5esnes p sayo^eq jo jaqiunu

166
Figure 21. Regression lines describing the influence of the initial
level of microbial contamination on the final climax
population size in fresh sausage.\\
c o
_rg3Q_OCLTOCcn o
Xro£UJcno
Data of J.G. Banks,this thesis
Q . — . Q Data of M.H. Brown (1977)2-OrData of M.J. Dowdell and R.G. Board (1968, 1971)
5 6 7
In i ta t level of confamination
( IoQ.q c.f.u. g"’’ sausage )
8

167
Figure 22. Influence of temperature of storage on the loss of free
and total sulphite foom a batch of sausage dominated by
Brochothrix thermosphacta.600,-
500
400
QJcnroUO3ro
c n
incnR
300
200
100
tI
0 2 3 4Time (d )
Key : Predicted contribution of total SO^ to sausage by retrospective analysis of the seasonings.
.o_ ^ .o# Total SO , 4°C A 10°C■ 22°C
O Free SO , 4 CA " io°c□ " 22°C

158.
4JcDOu
<urH
•H>
504-1
1c0c0
•H-pÜgÜcoÜo4-)•rH
ArH0mTDcrt3
mS'N441440ÛJpddp
ft14-)
ft0(DUC(Ud1-4ftcH
nCM
0)pdft•wCb
4J0
104-1
XftM
4J0
1
%gcftIOJftdcnddcn
cft
<b CN m e
o -
'•o
u
O -
"O- O oj O
pn E
O -
I O oJ,Lo
01,abesnes ^_6 n y : Bo]

<urC■P
ft0
ft30uftQ)x:ft 0)ftc •p0 ftftc f—10 d•H cnftd 1pft dd ft cn0) Ü 1—(u d 0d -d f t0 f t sÜ d >1
cn(Uf t P 73•H Q) d>d tnf t f t 0rH rHd X uÜ) •pp73 -dd ftd 0-d0) Üf t 0d pp cq0f t > ,w f t
f t 730 Q)
f t0 d Q)p d f td •p •Pf t E ftd 0 ftp 73 rH(D df t <D cne f td d +f t cn
dft d cn0 tn 1—4
00) d ftÜ •HdQ) cn cnd ftr4 cn dft d d)d d ftH >1 O
CM
%&•Hft
169.
--f
1_CO
• oL_
GO
UOJ 0 £ CN
CN
o
(0
aBesnes _6 n j r ] ^^Bo i

Xpft4JI1ftoft3Sftcuftftd0d0• HftdPftd(UÜd0u
■ H
ftI— Idcn
73dddft§ftcn
ft0CDpdft2 0)1ftft00)udrHU-idH
LOCMdPd•S'ft
§•Hddftp0cn•p5
■sftdd•p073dft2ddcn
d•pdftÜIPdrdft
170.
- o'"— (N CN
p ^E u
• P
• O
< H
"o
2
uo
OLaôesnes nyo 6o)

171
c0•H(ÜU4-)C(Uuc0uQ)-P•Hx:Ar-43ü)
T)CrOQJPO-PMluOQ)U
SfOp(D
W-lOQ)UC
U— o 2 : QJ ^E .u
L_o o
_Jm
+J
o.W
I-C-p
•psscq"%4->rCC•H§T30)enfüen0IT3enC■H
4
= °2eue s
ip euc euM en
OJ eueu P
eu P •HP •HD f: Aen A 1-4-p rH 0
D enen+
enrHen QrH0 epE en>1en '0(UC eneu 0a rHO U
OL,aOesnes n p 6o)

172
V)31-Ua
s■uco14-)O
Æ
O>-)CP(D
x :4Jdoco•H4-)fOU+JC(Uu
§u
iJ■HQ)r HS'ifO0)eny■pœp0OJ32 0)1p
po(DUC0)dr HpC
C N(DU3enPk
oo
P- ^ TD
fOP0 (Q1kleu<p
*s-qp
I-n(Uc•H
I(Uen(\SU)d03mc•HI
UC NCN
(UP•Har HDM
enI—IIg
0)p•HX:Û4rHPen
enP
5
T3euen0rHU
I— 1
s
01aOesnes |^_6 ny'D 6o]

A&cncnc0E0133Q)cna.p0x;p30kicn0)XPd0d0•HP(QkiPd0ud0Ü0 pp Ü•H (QX XQj Cu1—1 cnPtnT) (Ud XnJ pOJ Xcn •H(tJ k)U X0 pP 0cn XÜp 00 k cuOQ pcu cu pk4 >1 p XP X p 04P X r—1ft) '0 A Pu rH cn0 p pCk (d tn 1B d0) p +P s tn0 PP X cn Q0 rH XCÜ 0 B0) cn >1u Id cnd cnOJ P cn Xp Id 0)cn d cnp (U 0d d % rHH P 0 U
COCN<uk,pcn•H
# OvO
173
-o""O CNCMOJ ^G U
U-0 00rH0)Gi— X
•• Ô
-o UP O
a6esnes ,_6 n j ] ''601

CTiCN
QJ54DCn•H
4->•Ha ,
0) cnfC<D 1Ufd
•H cnkl f-HOJ 0+J pu EId >1n tnou 'ÜQJ CD4-> cnc 0u f—1
uCm0
cup
•P •H5 P0 04>4 r-4Cn 3
cnOJ
p +p
c cn0 r—1
0c P0 E
■H4-1 cn(d>4 Gp cuc 0401 OÜc0u
fd0) Pp Ü■Hp pCL, O41—1 cn3w 0
cuG p(d p
01 XCn p(d 0>4 p0 4JP 0cn p
0M4 00 0
CQQ)i4 >13 Pp(d Pl4 CUcu p0 , rde Gcu ■Hp E
044 P0
CUcu cnu Gc cncu 33 (d1—( cnpc GM p
174.
uCNCN
L_o
OI—IA
O -
O -
U
O -J o
_J
OL,aBesnes ^_5 ny ] 6o)

176
Figure 30. Influence of temperature of storage and sulphite
concentration on the rate and extent of growth
of a, the total viable count, and b, the yeasts in
sausage dominated by Brochothrix thermosphacta.
o
Mean doubling time up to climax
population in sulphited sausage
Mean doubling time up to climax
population in unsulphited sausage
Climax population in sulphited
sausage
Climax population in unsulphited
sausage
Total viable count
b; Yeasts

Cnc
i
20r
16
12
8
0 L
b2 2 r -
18
U
10
OO
O O
O
4 1010 22
Temperature ( °C )
12
11
10
9
8
8
7
6
5
QjCniJSICnCisc<c?"
c•5C)?CL

177
Figure 31,
20
16
12
8
4 -
Ib ■§ ■§ cI
2S2
7220
16
12
8
(J'
Influence of temperature of storage and sulphite
concentration on the rate and extent of growth of a,
Brochothrix thermosphacta and b, Lactobacillus
spp. in sausage dominated by Brochothrix
thermosphacta.10
9
8
7
O O
O O
10 22 4 10 22
9
6
7
4 6 5
4 3
2
Key: As Figure 30.
Temperature ( ° C )
a ; Brochothrix thermosphacta b; Lactobacillus spp.
ItiH.,-U
-2g-ClX03e

178
Figure 32. Influence of temperature of storage and sulphite
concentration on the rate and extent of growth
of a, Streptococcus spp. and b. Pseudomonas spp.32 r
16
8
I•CJ:d•§cOD0)5:
in sausage dominated by Brochothrix thermosphacta a
O
O
360 r
22 -
0
10 22 4 10
a. Streptococcus spp.b , Pseudomonas spp.
Temperature ( )
Key: As Fig. 30,
I'Tozi'tZubro
cz•2Z)&CLX
CU

179
Figure 33. Influence of temperature of storage and sulphite
concentration on the rate and extent of growth
of Enterobacteriaceae in sausage dominated by
Brochothrix thermosphacta.
30
c 20
1 • ■C3cft)Qj
10
o
10
10 22 10 22
ÎCnt)
o
-2§■CLI
Temperature ( ° C )a Enterobacteriaceae Key: As Fig. 30.

180
Figure 34. Influence of temperature of storage and sulphite
concentration on the pH drift in sausage dominated
by Brochothrix thermosphacta.
6-5 r~
60OJro
S. 5 5OEoJZ
CL
50
15
10
0 3 1 8
Time (d )
+ sulphite, 4* C / \ + sulphite, 10°Cn + sulphite, 22°C
- sulphite, 4 C- sulphite, 10°C
H - sulphite, 22°C

iOi.
Figure 35. Influence of temperature on the loss of free and
total sulphite from sausage dominated by yeasts
700 r
600
500OJcnoto=}otocn(No00
400
cnR 300
200
100“-
0
O Total SO , 4°CA " 15°C
3 4Time (d)
-A
Free SO , 4°C 15°C
-AJ7

p
m4-1M(d(U>1CD5
4-10x:
tuen0)5C0co•H4 J(T3VC8C0u<u
4-)• HCur-iDUJT)Cn3(DtiD4-1(13V403a
4-1003UCr—I 144C
m
Den•HCl.
W4JCO(T303>1
T3034J(dd■He0p03
t/3D(dtoC•iH
ât/3to3MM" MS镧4 4Ü3
(d(d
4 4Ü(d
iI03
4 4
X(4- d
4 4
I
&033MM" MÜI4 4Ü
u I
(d4 4Ü(d
i- q4 4
Xk- q4 4
Ig
t/34J10(d03
> 4
rd I
Ô •
182
_TD
U |
U 0 ^O U")Tf rH
5•H
g
034J•H
310
• <-( m -
-I r-
H -
J O rQ I
UOo in«Cr rH < O.
034-1•Hqar-43to
034-)•H
303
O <J
3 I
05esnes^_5 ng : ^^6o]

âenCO30BB-ua%-uoo440q4-)g!henCD
q4->
Coco•H4 JfdH
4 J
sucouCD
4-)• r4
ar—I3toTSC3(D
32CDa§
4 J
4 40CDUC
BI— I 4 4c
m(Du3a•Ha
to4Jto3CD>.>4PTD3
4 J3C•HE
3a3333toC•H
333U3•H3434JU3XI0343
4 JCM
TiC3
âtoto3q
O33a
XI
333U3•H343
4 JUI3
4 JCWU|
(03q
i73Sa
rQl
i "u
3 34-1 4 J•H -Hq qa a
• <
183
a iEpn -
Xll
atoto3B8O
4 4
as4 4oo3 I
34J•H
arH
S
CD
34 J•H
â3to
_ lm
0<3pn E
3|
. 7 ou'^aoesnes 6 'ng'D 5oj

184.
Figure 38 Influence of temperature of storage and sulphite
concentration on the pH drift in sausage dominated
by yeasts
Q + sulphite,
/ \ + sulphite, 15°C
^ - sulphite, 4°C
- sulphite, 15 C
7 r
QJm 6 cQJcn o E0
1
X 5CL
4
0 3 4Time (d)

105.
Figure 39. In f lu e n c e o f te m p e ra tu re o f s to ra g e and s u lp h i t e
c o n c e n t ra t io n on th e r a te and e x te n t o f g ro w th o f a ,
th e y e a s ts and b , Brochothrix thermosphacta in
sausage d o m in a te d by y e a s ts . 20
O
Y e a s ts
c
16
12
0 L-
O
Ch•SI*§cI
b Brochothrix thermosphacta
32
16
o
4 15
9
-I 8 7
6
5
1 8
]
IChZiu
5§■aXfO5Cj
O
15
Temperature (°C )
Mean d o u b lin g t im e up t o c l im a x p o p u la t io n i n s u lp h i te d sausage
Mean d o u b lin g t im e up to c l im a x p o p u la t io n in u n s u lp h ite d sausageC lim a x p o p u la t io n in s u lp h i te d sausage C lim a x p o p u la t io n in u n s u lp h ite d sausage

186.
Figure_40. Influence of temperature of storage and sulphite concen
tration on the rate and extent of growth of a,
Lactobacillus spp. and Streptococcus spp. in sausage dominated by yeasts.
76
8
O8
7
6
5
■ I
'§c:I
56
32
24
76
O
75 4 75
^ Lactobacillus spp.
h Streptococcus spp.
Temperature ( ° C )
Key: As in Fig. 39
Itn
-IIISc

187.
Figure 41. Influence of temperature of storage and sulphite
concentration on the rate and extent of growth of
a. Pseudomonas spp. and b, Enterobacteriaceae in
sausage dominated by yeasts.
I
oo
1
O
0
15 15
8
7
6
5
4
9
8
76
5
4
C:•5
§■CL
I
a Pseudomonas spp.
b Enterobacteriaceae
Temperature ( )
Key : As in Fig. 39,

Figure 42. Simplified dendrogram of isolates of Enterobacteriaceae
from sausage and sausage ingredients
Footnote : % similarity calculated with the
S coefficient, clustering by UPGMA SMCluster:
1 Yersinia spp.
2 Yersinia spp.
3 Escherichia coli
4 Citrobacter freundii
5 Serratia liquefaciens
6 Serratia marcescens
7 Enterobacter cloacae
8 Enterobacter agglomerans
9 Hafnia alvei
10 Hafnia alvei
11 Klebsiella spp.
12 Unassigned

189
70Percentage Similarity 80 90 100
— I
56
8
1 0
111 2

Figure 43. Fermentation characteristics of Lactobacillus spp.
isolated from sausage.
Footnote ; ^ Homofermentative streptobacteria group IB (Sharpe,
1979). Isolates (43) identified with Lactobacillus
casei var. casei
b Homofermentative streptobacteria group IB (Sharpe,
1979). Isolates (28) identified with Lactobacillus
casei var. casei
2 Homofermentative thermobacteria group lA (Sharpe,
1979). Isolates (6) identified with Lactobacillus
salivarius (L72, 73, 79-82).
^ Homofermentative thermobacteria group lA
(Sharpe, 1979). Isolates (4) identified with
Lactobacillus helveticus (LIO, 37, 46, 55)
I 1 No fermentation
Fermentation of organic compound after 3, 5, 24
or 48 h incubation
Substrates0 Control 21 a methyl-D-glucoside1 Glycerol 22 N-acetyl glucosamine2 Erythritol 23 Amygdalin3 D-arabinose 24 Arbutin4 L-arabinose 25 Aesculin5 Ribose 26 Salicin6 D-xylose 27 Cellobiose7 L-xylose 28 Maltose8 Adonitol 29 Lactose9 gmethyl-xyloside 30 Melibiose10 Galactose 31 Saccharose 41 D-lyxose11 D-glucose 32 Trehalose 42 D-tagatose12 D-fructose 33 Inulin 43 D-fucose13 D-mannose 34 Melezitose 44 L-fucose14 L-sorbose 35 D-raffinose 45 D-arabitol15 Rhamnose 36 Starch 46 L-arabitol16 Dulcitol 37 Glycogen 47 Gluconate17 Inositol 38 Xylitol 48 2 Keto-gluconate18 Mannitol 39 6 gentibiose 49 5 Keto-gluconate19 Sorbitol 40 D-turanose20 a methyl-D-mannoside

191.
3tM
l l

Footnote to Fig. 44.
^ Aerobic incubation
b Anaerobic incubation
Reaction positive in all strains
0 Reaction variable
Reaction negative in all strains
i . Streptococcus faecalis
i i Streptococcus faecium
i i i Streptococcus Group I
i v Streptococcus Group II
V Streptococcus Group III
Substrates :
0 Basal medium
1 Salicin
2 Sorbitol
3 Glycerol
4 Azelate*
5 D (+) trehalose
6 Inulin
7 Mannitol
8 Arbutin
9 Mucate*
10 Arabinose
11 Pimelate
12 Citrate*
13 Adonitol
14 Cellobiose
15 Inositol
16 Raffinose
17 D(+) Melizitose
18 Sarcosine
19 D-ribose
20 Maltose
21 D(+) melibiose
* No reaction due to
endogenous acid in
organic supplement

193
Figure 44, Aerobic and anaerobic metabolism of organic
substrates by Streptococcus spp. isolated from
sulphited sausage
1ii
iii
iv
â 10 11 12 13 U 15 16 17 18 19 20 21
0 1 2 3 4 5 6 7 â 9 10 11 12 13 /4 15 16 17 w 19 20 21
i • • • e • • • • • • • # • # • # # •11 # # # e e e •
iii e e e # • • e # • • e e • # • e e •
iv • # e # • # e • • # # e # • e • e e •
V e e # e e e e e

194
Figure 45. Aerobic and anaerobic metabolism of
organic substrates by Streptococcus snp.
PI isolated from unsulphited sausage
0 1 2 3 ^ 5 6 7 Ô 9 10 11 12 13 14 15 16 17 IS 19 20 21
1ii
iii
iv
0 1 2 3 4 5 6 7 S 9 10 11 12 13 14 15 16 17 18 19 20 21
i e e e e m • • • • • • • • • • • e •i i e e e • • # •
i i i • # • # • • • • # •
i v • # • e • • • e e e
V e • e e # #
Key; As in Fig. 44.

195
Figure 46. Influence of sulphite on Hafnia alvei in
culture medium
^ Modified glucose glutamate medium (MGGM) pH 7.0, 30°C
MGGM + ca.125 yg free SO^/ml
MGGM + ca.250 yg free SO^/ml
I I MGGM + ca.500 yg free S02/ml
8 r
E
ZD
o 6cno
4
0 10 20
Time (h )
30 40 50

x:u4JA
c•H
Q)+J■H£CUr—1 0 cn0)CD!h4-1
O4Jcn§■8cntü•rH
Ü■rH6CD■^-WsscnCD
ACD
4-t0&•rH
-p•HcncCDcn4-10CDS’ItPd't'
CDp
g.•rHÏU
§•H-P•HA•H
c•H
4-1O
CD
rPS
•H%ECDPSrHeI
CD-PS■P0s
CD
-P2CDg'CD 4-1 Eh 0
T3CDcnd
I•HTDCDS
4-)cnCD-p
c•HItP-PCO
pO
It•PA8Ü•P2
U
Oro
-P0
CDcn0 u 0 '—Icn1CDcn5sS’p-p•pa-pcnItCD>H
MD
£O
a
>1pI-p■p•p§■8PCD4-14ha
CO
Icn
I8»PHJCO
cn3MM•pÜ
I■uÜ3
-p0A
CNcri
4H4-1
8ggQ1It•P§S
en
Benft
ISs
pA
CD•P
ipaenCDcn83rHenT)CD•pp•p
I
CN
UoCN
§en
en
CD
ftpü
I0ftCDU gCDÀ ftA •p Pcn pCD %t P •pM U PM ftCD P Pq 0 0Q P -d6 (D ÜM P 0cq es pco H CQ

197.
OOun
£2 ■XD<uÊ o o3QJ
3U
Een o3 ^
rs i O ui
QJ QJt §CN
OOun
oo
oom
ooCN
o o
O L _
(/)inm
S5Umt01Ck:QCi
CLOLl/î
i8-S8-L.S..
o_Q.l/î(/)
k»'Q-Sk»OD
CLCLVO(/)§Oe
Qjo!
SQJUro(_QJWro-Qo(_QJ
OO
CLOutoroQJc:Oe
CN un vO

198
T3M"HQJC
M
tou-iO
enQJû .> .4-)OuOJw
X■HUJCoQJ-U•Haf—Ioen
Dc
gX!nc
Q)euXIW
cr>•HXU-l0euDc
?I— 1u-iC
4JrUeus-l3I— I
sxu■p(UJQC
•H
enQ)A>1-P2euen
X•pen
peu
3-P :
X•pen
Ceueu34-1eu en
X (U
c &O 4-J•P o4J Pnj eu
•p en p
•p
O Q•pIII
O O '■p
o O O
•pCD O O
•P
OCsl
LO
-O
u n
sO LPi - i-
_)uj n;:^^5oi
30-q
Ü'2
?fOIPiI-u
3OCr,0%
N4J•pMP&E3-P§eq
eneu
g;4-1Opeuen
œ
eup3en
(t)MMQ)q
M
(%

199
et;
u
-prOiguW-l■ps:OarHS(H
-pDc(d(Ud)py-i<4-10wœ0P]
d-i(% «•PP p P•p •Pp P0p; lâü 0 0P P: -q•p ü üP P•p •pfdMM id fdCD M Dd rpc; M rp0 CD ip CDS c •P qM P 0SCO M rpfd 0 fdip 10 -q CO0 üPd P Æm p •p piH •p > •p(H 3 30) fdu T3 M TlCD M CD-P CD Po id q fd1—(rH 0 rH3 S 3U M Ufd 0 fd 0Ü C co C•P •Pp; Æ■p CD p> CD•H TS •p T3s 3 >1S >1o x: O Æu T3 CD CL? CDs CD 13 S $ 13P 1—1 4-1rpfd fd 3 30) rp P rH 4-1-p 3 CD -p 3 CD (Id U U Id U O(—1 0 fd rH 0 fd 10 C 3 Co •P + U •P +l0 0c S S C S 2•H ü o •P C2 Üc ü o C o CL?n s s 3 s 2 1< OCN CN CM *■ o
O O o CN CN CN ow co w O o O
co co CO1—j1—1 pnJ fd Id CD 0) CD■p -P +J CD CD CD0 0 0 P P PH H H ïn k, k
o -1Orsi
0 - tn
O -
O - LT>
O -I
Oi_n Ooun
in' ' os
oLTIrv
OrT
'C CDP
CD 0p c3 pen 0
•p 0t , (p

200.
U
■pn3§gI5444
■H
'âI— IoWI—103O+JCrd
<U(U
4 40MU)0
Seu54Pen•H[4
m
en•H[4C
• H
W
>1
■ e
0 4 <
ofNJ
O - m
O -
Ô - LH
OOO
OLO OOLn
oLHCSJ
^ j \ u O S 6 f V

201
w>1fdn
coI— I
a
CM I—IVfdcü>-U
u
+Jfdw>.fdT)
Mou-l
'Suo4-)m
§S
U-l
ü-Ha
r—IC3U)
fd4 JO4-1
fdeueukl
144
U-l0U)enS
UCNrH
U
r—IU-tU-l
c■H
WC
9 4 <]
o -
ô -
o -
o -J o
Ln
<uE
LA
SO
oLA OOLA
OLArsleuklaen•H ^jui OS 6rif

202
<4-1•H
0-qu ÜD P•p>4J03 OSMt; MQJL-l q-H 0Elà 30 to-c:Ü 440"M inrH03 rHM QJU03q U30 Oe 1—4,03 OSto Üc x:0 +j•pQJ là-p•H 03j: QJA +Jr—1 033 rHCO 3U03 03 33 ■P
St303 U3 S3QJQJ -PP •P04 XC44-1 rH0 3inQJ 3U q>3QJ3 1—1 L-l #3M
CNin QJQJ ■Pp 03 3Cn 4-J•p 00k
«14-1•rH
IIsS03"qJ
Ito5•H!5TlO+J03r—IDUOc•Hsg■B-P■HrCr43
H-t•HP:k0?k•H03MM
1M
•P3
+J03f-43Ü0C■H
•H
3M
(D
I30)u3
I
# o o
on
oCNJ
LO
Ln
un rsi

2 03 .
u0CNr-HV3
cc44•PP
0
üp•pii03MMeuqQSM03toC0Q)-P•PXA
i-H3w
03
0X03q3QJQJP44 CN44 LD0
QJ enU ■pC t,QJ3 Cf—4 •P44a C/JM <
on ..m QJ-PQJ 0p C3 -pen 0■p 0pL, k
O(SI
LA
OJE
Ln
—* o
es OO vo un ^ m01.601
(SI

-pn3(/)>1n3
204
P04-1T3Q)Psm
U
cn4-1-p
o•g
(QMMSsMC0
•H"âr 4Dto
Tl§0
ctd0)(DU4-1
4-10cu
8 r —IU-lcM
m0)p3Cr>•Hk,
to
00
p04-1
uDCNt-1+J(C3CCD-P
•H
U
-PttJ
•PA
I— I 0 to C D
•H
O
O O
o
■O
dJ
o OO m

205.
Figure 55. Range of sensitivity of representative micro organisms
to free sulphite in turbidometer culture medium.Free S(1 (xjgmf culture medium )
0 50 100 150 200 250
Candida sp.sp.Crypf-Qcoccus sp. Rhodotorula sp.Br. thermosphacta thermosphacta
I NOj STASIS
I STASIS
Ps. fragi fragifluorescens C
at freundii diver sus I 1Ent. agglomerans agglomerans
cloacae cloacae
L_ I
I IEsch. coli coliHaf. alvei alvei alvei
I . I
Salm. ana turn derby g o e r l i f z hadar kedougou senffenburg
Salmonella sp. Stanley typhimurium Virchow
L -J
CZZZD[=1LUI' . I
CZZ=I C. Z ,1
1______3I 1
Serr. liquefaciens marcescensYers. enterocolitica I — J
01
50 100 150 200 250-ty— I
750Footnote : ] Range of inhibition
Media and temperatures of incubation used as in Fig. 47.

206
Figure 56. Influence of free and bound sulphite on Salmonella.R o
virchow rif. in turbidometer culture medium at 30 C
X V Point of addition of sulphite
Point of addition of binding/neutralizing agent
6
V_JO
Footnote:Time ( h )
Unsulphited MGGM
^ Sulphited MGGM
n MGGM + acetaldehyde/hydrogen peroxide + sulphite
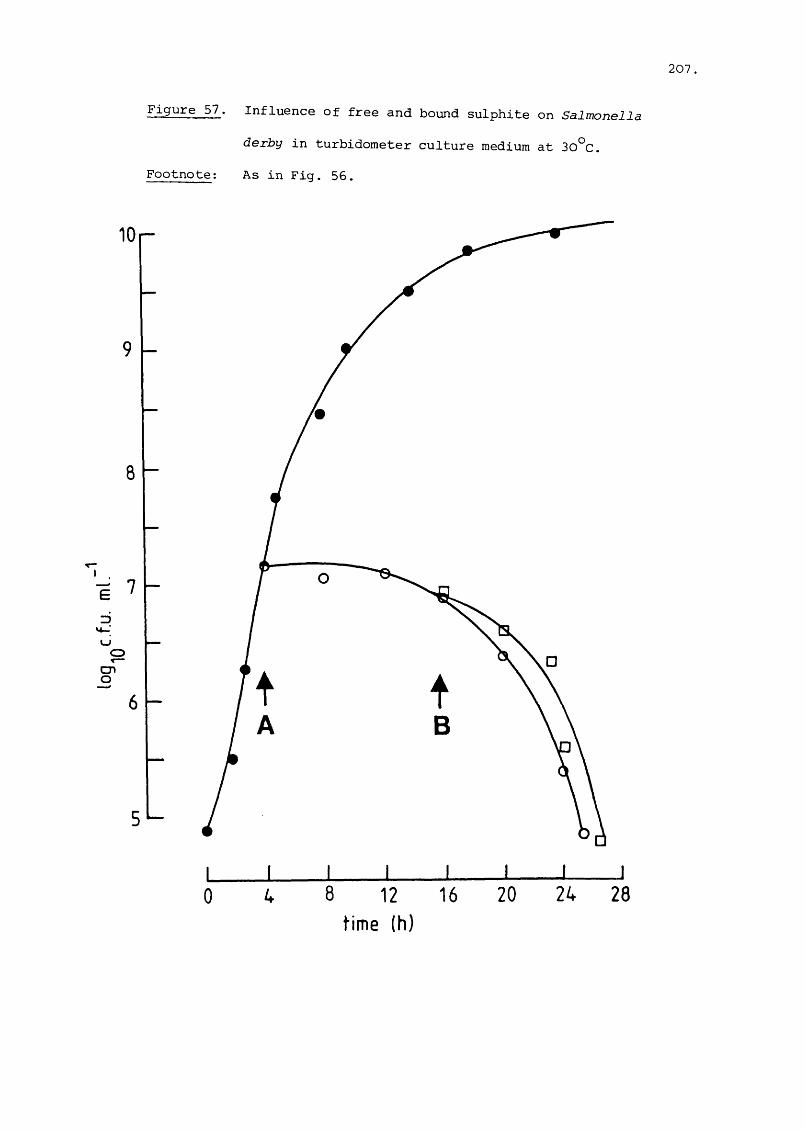
207
Figure 57. Influence of free and bound sulphite on Salmonella
derby in turbidometer culture medium at 30°C. Footnote: As in Fig. 55.
lOr-
8
cn o
0 8 12 time (h)
16 20 24 28

2 08 .
Figure 58. Influence of free and bound sulphite on Hafnia alvei
in turbidometer culture medium at 30°C
As in Fig. 56.Footnote :
8r~
yo
cn
Time ( h )

•H'g•Hb
Oa0•H4-)U
2eu?rO(44o0)
4 JdLlrC
oO'(Ur C4J
codo•H44(dM44d8§od)44•Hx:Ar HDwm(US4144
(44
OoudgrH44d
euS'cndrcJenr)ü•H•Er-Hgiu
144
i-Hg•H44end(U>1
cn44gS'
pd)44df—Idüs•H§•HT3Q)ed)endbddjcn0u44•Hd44cnfO>1%44dSd)
IenId)cn8-H O tn en
P -eu 44 44 cn •H d rC oe u > 1
15D 3
§>4
5•H
%44drH8B•H
i•H%S
d44O44en
gU)
44•H'ârHdco
OLD
>•H44gUeucn(UAX
rd
•HArHg
d)
>WeneuAeu44dueu
1
d)>•Hd£d)end)&g
g>H44end
rd44•H!5peu1SrHdB■H44B•H'ge
(U.£-PdgcneuA1440d0•H44dp
gu§ü
•Hrd
•HErHdcneueup&4
d)>•H44dgcneu&440d.344dP44gUgüd)442«i
>■H44d£eucneupeu44Odo•H
îd0ug
g>4
44end>1X44•H
teuSrH88•H8•H
6
u•Hendrd
5g•HenddO•H44diJgB0U44gS’d-H■B•Hrd
d I rdl
44 cn0 d
044 -Hd 44d) •He Pd) du 0dcn Ud Ud) •HS rd
0deud d d
d deu eu d3 3 euen en • P
dd) eu d d'd 'd cn•H •H en Uts. fd
° o' eu mrdl d I 44•rH 44
d d rd d-H ■H QvrH cncn en d dd d encn en d drH rH "H 0Q 0 U) •H
K II 44rSco co 44 du
ül Pi H
enun
d4dE•H

oo
o
o x :
oom Lu
Ses»
Ooo
o*
•o
•o

211.
Figure 60. Influence of sulphite concentration on the growth of
Salmonella virchow rif.^ and Enterobacteriaceae in
sausage meat at -20 CRFootnote: ^ Salmonella virchow rif inoculated into
unsulphited sausage meat
Salmonella virchow rif.^ inoculated into sulphited sausage meat
m Enterobacteriaceae in unsulphited sausage
n Enterobacteriaceae in sulphited sausage.
5 r-
OJcn ruuo3fDihIcn
3<4— 'wcn o
3
0 4
Time ( d )

212
Figure 61. Influence of sulphite concentration on the growth
of Salmonella virchow rif. and Enterobacteriaceae
in sausage meat at 4°c.
Footnote: As in Fig. 60.
8
QJor»ro/iIDmI/)
I 6 cn
Z)y-ru
cn° 5
3
■
■
0 3 4 5
Time (d )
7 8

213.
Figure 62. Influence of sulphite concentration on the growth of
7djcn mI/)3muoIcn 6
3u
cn° 5
Salmonella virchow rif. and Enterobacteriaceae in sausage
meat at 9°C.
10 1
Footnote: As in Fig. 60.
3 4 5Time ( d )
8

214
Figure 63. Influence of sulphite concentration on the growth of Salmonella virchow rif.^ and Enterobacteriaceae in sausage meat at 15°C.
Footnote: As in Fig. 50.
8
CUcn m00ZDfO00
cn
cn o
3L-
■

Figure 64. Influence of sulphite concentration on the growth of
0»cnro003m00
Icn3
cn o
3 L
Salmonella virchow rif. and Enterobacteriaceae in
sausage meat at 20 C
10
Footnote: As in Fig. 60.
3 4
Time ( d )
8

216Figure 65. Influence of sulphite concentration on the growth of
Salmonella virchow rif. and Enterobacteriaceae in
sausage meat at 25 C.
QJcn m003ro00
cn
3
u
cn o
9 r
0 2 4 5
Time (d )
6 8
Footnote: As in Fig. 50,

217Figure 66
600
500
r - O
— 1
\
400
ro<LI
I 300D œ
Ien rsiO00enq
200
100
O L
0
Influence of temperature of storage on loss of free and
total sulphite from sausage meat stored in Petri-dishes
Open symbols, total sulphite
Closed symbols, free sulphite
1L
2 8
Time ( days )Footnote : Q — Probable fate of total sulphite added from ingredients
to bowl choppero

2 1 8 .
Figure 67. Influence of temperature of storage on pH drift
in sulphited sausage meat stored in Petri-dishes
6 7
QJ
rocOJcnoEo
5-9
5 1
U-3
3-5'-
0 3 4 5
Time ( d )
7 8
Footnote': 4°CO
A
A□ 20°C
9°C15°C
25°c

219
Figure 68. Influence of temperature of storage on pH drift
in unsulphited sausage meat stored in Petri-dishes
Footnote: As in Fig. 67.
6 7
OIm cQJcn o E o .cIo
5 9
5 1
3-5>-
0 2 3 4 5
Time ( d )
8

220.
ft)ft: 44ft •HU44 P0
QJ f tA 4JId 44T3 u 0Ctt! QJ QJ
ft: ft 4-1T3 4-> fd(D L44-1 c O•H 0 QJ
f : II ft!ft QJ ft)rH Li O3 P CM Pw 4-) U 0 IIfdLi 0)fd QJ P Ll CM
ft 0 P Llc E •H 4-1-H QJ 4-1 fd4-1 Id L4 P4-1 p QJ 044 44 •H ft •H•H 0 s ft)QJ fdT3 QJ QJ 4-1 P
U 4-) •HX C QJ 44 Ef t QJ T3 0 UP QJ44 r—1 44 QJ 4J0 44 0 U QJc P P<D UÎ ■r4 4-) QJ4-1 0) P P 44fd Æ QJ QJ rH 0Li tn ft: •H M-4•r4 4J Ü p 4J0) T3 •H •r4 Px: 1 ft 44 QJ4J •H c 44 QJ •HLi ■r4 QJ ft: Uc 4-1 ft) 0 ftj •H0 0) •r4 U 44ft Ll ft 440 u p QJft C cn 4-1 -r4 0Id -H QJ fd ft] ULi T) QJ •r4q E Lt4J (U QJ U ft)W L4 P QJ cn Id0 -r4 f t QJ QJ44 4J --1 fd p E0 cnP P QJ QJ0) 4J 0 fd p f tL4 fd •H cn •r4 fd0 QJ cn r—1 cn4J E cn 'P Pfd QJ P fdLi Q) Ll ft) 0 cncu f t f t •H •Hft fd QJ -p cn PS cn Ll f t cn q0) P —1 QJ 4-)4J fd 1—1 p Ll ■Hcn fd cn ft ft:44 •H QJ ft0 4-) p Ll rHQJ P •H P01 4J QJ Ll cnÜ •H P 4-1 Id Pc rC Q 44 QJ P
f t ft •H p0 1—1 X Ll •H P1—1 p W ■H44 cnC CH P fd 1 ft]|
ftikù 0)4-10) 0Li pP 4Jft 0■H 0ft ft
m
o
CNI
Om
LnCNJ
oCNI
Ln
w001 c_ 3mt_OICLEOI
Ln
'Q.lLnrn
LnCNJ Ln Ln
Ô( 4.'un Hd ) 4^up piDy
ON
oon
inCNI
oCNI
in
wodic_3gQJCl.Edi
in
(T3 I
inrn
inCNI
in in<3
( s j iu n Hd ) IJ i jp PPV

221.O)
rH CDX3 e nfd fd
■H cn> C CD P
fd e n fdr —j fd cnfd e n U)
H) C P X0 ■H fd CD-U > U) X
p •HX : X X
x i CD O i-P - p X 1—1
fd -H PQ) X cn
fd E A CX p
«H CD p0 e n U)
fd cnU) 1—1
■p p cn 03 p 1—1 X0 U) Q sP Xe n c Ë cn
•H SC en X0 cxi CD
c cnC 4-4 CD 00 •H Û4 rH
■H p O U+ Jfdp 0-P X :C ü(D Pu - p u Uc U 0 00 0 LO ou fq rH CN
Mm M
- p CD•Hx : 1 o < □«—I Mp fd(0 co
c ufd T3
T ! P0) C 0e n P rHfdp CD iH0 P p
- p CD ■HU) 0 X
fd 04-1 •H P (%0 p ü
CD •H XCD 4-) E ■HP U pP fd rH
-P XI P X CDP 0 •H fd 0P P X CD XCD CD ■H 0 U ü& 4 J C u fd pE C •H •H ■ p(D W CD P
+ J X l—j CDCD e n X X fd
4-1 X •H p U M0 -P X •H fd M
> X CD(D !>i 0 qU X tH 1—1 p 0C rH fd CD Em P X X MP ■P P 0 C fd
iH C co En W to4-1 p pC 0 cH u p fd 1 XI u |
o
mu&•Hht
0)4-1
g+J0ok
□»
j•9
'o*
• o
ül
E
-Ql
I
01.a5esnes _6 n j y 6oi

222
SALMONELLA CONTAMINATION OF SAUSAGE
The incidence and level of contamination of British fresh sausage with
Salmonella
Farm animals, especially pigs and poultry are the major depots
of Salmonella, one of the commonest causes of food poisoning in
developed countries (Bryan, 1980; Silliker, 1980). Indeed, there
is abundant evidence that food products, for example sausages and
sausage meat (Table 32), containing materials from such animals
commonly have a high incidence of contamination with these organisms.
Major surveys of the British fresh sausage for example, have
demonstrated a wide variation in the incidence of these organisms.
Public health reports, on the other hand, indicate that only
occasionally have British fresh sausages been implicated in out
breaks of food poisoning (Table 33). Of the many reasons that
can be put forward to account for this situation, the general
practice of cooking sausages immediately before consumption may
be the major one. Another might be the actual level rather
than the incidence of contamination, the former can only be
important in view of the observations (pp.157 - 164) that the
bacteriostatic activity of sulphite is temporary in sausages
subjected to temperature abuse. Indeed the recent observations
(Gill et al., 1983) that ca. 10^ Salmonella cells can cause
salmonellosis calls into question the long held view that at
least 10^ Salmonella are needed for an infective dose (Bryan,
1979). The work described in this section sought to establish
the level of contamination of sausages made in a large factory,
and to identify the major sources among the ingredients.

223.
c0■-44JIflcE c<a 0>
w V >E c
0 O'Ü uO' 0)m kU) 0 (U3 £3 SCfln d)c
TD kC £30 (/I VO0)U1~H <u
£ k0) JJ HIc c £0 oE k HIm 'O"Q a4) HIc £0 Ü)U) (U(U O'CÜ fl>1 4Ju c E0 0) 3k u<u k kw (U 3
& Ek0 £
Q.(0 3)z
0 0 Qj IM
CNO CO r- k
VÛ in cn cn (71 HI<7» (7i CO in rH rH £CM rH fH T f cn p ' C'3 ' in rH <71 HI VO(7 \ >1 (71 VO rH HI £ CX (71f-H Tl HI M rH >1 <7i (A HI k
C rH U £ rH cn <0 0 rH iQK] rH "H VO <71 OH U(N HI TJ O <0 <n M Q £CO -H k cn £ HI M HI <71 rH U cn VO HI ■a T l<yi m HI W VO <0 4J o cn c 3 ' <0VO > (7\ cn - 0 ■H HI <0 rH £ rH N IQ TD <QH 4J HI T3 rH H ■U U (71 00 c HI U C
<71 HI <0 C HI HI k C <n 0 k (Q 3 HI*—1 £ HJ <k k HI 0 HI in HI k > iQfH > U 3 X C C HI •u k £ rH HI HI 3 C E cn 3HI k (A ITJ £ u HI 0 C HI </l in «5 HI k HI U £ 0 (A VO 0k > k 4J HI ■U •fH (0 4J HI c HI E 3 £ HI E c M C c <A (71 ink k HI 4J HI c rH fH £ £ c e H £ £ E HI k k kifl <0 £ 3 3 >1 HI HI <0 :3 0 0 0 0 H) <0 0 0 Qj 3 3 HI HIQQ U U Q ffi Q U h o ac a *3 & 0. OH OH cn W H > 3 3
I(US'
IiI
m1
O'O' cn
m V(N 1 in o3
<Q£ Ü
3,0)
I
k,6
cx>o
•2 I
•§
I
3Z H & &0) V)
t o O «N
Q.
-2
>1 Ü1 k•13 2c yX k C
IS. I
in (A <A3 3 3 3 30 0 O 0 0■kk k k k k<Q (Q IQ IQ iQ> > > > >
HI kHI Û4 IQrH E HI CX > HI«TJ HI E(A (X rk iQ CX
E cx 0 E(/) O' iQ IQ(A Cn HICT in (ArH cn CX cnO 1 cn E
o o (Q 3 o'—1 CN (A O1 1 3 •k3 3 3 3 £ 3•k •k IQ •k
HI£ HI HI > £ 0 £3 > > + 3 in 3HI + + HI HI(A HI inHI dP dP vO HI N in HIk k •k vO kCX 00 O CX in CX
in nj IQ HI3 3 HI EiQ IQ E(A (A
0> HICX3
X O' tn 1k k fO IQ HI0 0 HI (0 CA IQCX CX
£ S' s s 1HI HI HI £ £ IQ tn in CAHI cn HI HI cn cn HI HI HI (A HI (A HI D HI HI HIcn IQ cn cn IQ IQ cn cn cn HI cn HI s <d xs cn X) X) cn cnIQ iQ IQ (A IQ iQ iQ k iQ k 0) 3 IQ 3 3 IQ iQin 3 in (A 3 3 <A (A in <k in <k iQ in iQ IQ (A in3 IQ 3 3 iQ IQ 3 3 3 3 in f—( 3 3 3iQ <A IQ iQ <A (A (A HI iQ iQ iQ HI £ IQ £ 3 (0 HI iQ HI HI IQ iQlA in (A 3 Cn (A (A (A cn (A (A (A IQ L, tn tn cn in in
£ 0 £ £ IQ IQ ■k •k <A <U iQ iQ iQj<; CA a <A (A in J<( X <k in £ X £ U in in in X Xk HI k k k HI HI 3 k k HI 3 •k k IQ {/) 3 k 3 3 k k0 k 0 O iQ k k IQ 0 0 HI iQ k 0 k 3 •H IQ 0 iQ IQ 0 0cx Cl, CX (X > Cl, Cb cn CX 0, 00 cn CO CX CO i3 > cn cx cn cn cx CX

224
Xroe
gc•H
H'•Hcocn
•HoA
?0 X
(0M
(D1gXOcn
•SCDUXg
CDUCg:CD
X
S
en XCD Gcn 0Gu G
XX O0 73 X O
CD G inp > CD 1CD rH P OX 0 X irtS >G G2 X
CDX5-r4 r—I
6 •H
OLO<J\
C£CTi
vD mVO CTi
cnOa
CDS'en
§tn
enX
'SXoc
XX(0
eCD
scn
3tn
cnCD
scn
3cn
.Xa
T3G(T3X(Ü
S
IenG X no nO cn CD
ü ®U CD O XOa a
1oaCDsen
3en
en ’vrCD VO G <Ti0 X t )1c X no noCD X m CD
4"I— I CNJ
G X >1CD X cn PG» G 0G G P Xen CD 73 X 73 73 cn UG tn G G 0 p GG X G G >1 p 0 >1 CD Xcn 0 U p 73 X P X en
p G 0 0 ü CDp CD G G CD X 73 X X en cn0 ü 0 ü X 73 ü a G U G G cn X
P X U Q G 0 G G X X G GCD G X X 0 X X X P G ü CD
0 G X X X cn a B X P p GG en G a X en 0 b i 0 0 73 Gcn X X G cn X G P X u 73G CD 73 X 0 X en X X X P CDG fH CD G en p en G X Pen X X X X tn cn CD 1—1 cn cn X 0 cn CD
G G O X G CD X X CD G G X G XCn X 0 CD a X u ü G ü X X X üG 0 ü X en G 0 X X 0 G 73 G G XH P G 0 p p G CD p P G X P G> a H œ 73 a m « a Q G 73 X
en
X X X CD
® X CD G D> O Gen X G X G G en X
CDX pCDCD X X G
73 en G CD G Cn
G•s0O a cn
X E a0 G 73 3 a p
X CD X P 3CD X X X p CD Xa CD G X 3 3 X X G>1 G u p G E P X CDX Q X 0 a X 0 CD 700 G X a X a 73 GP X a a X GCD G E CD X a CD CD Pen en X G en X G ■G X

225
Literature Review
Sources of Salmonella contamination of pigs
Many surveys (Table 34) have indicated a wide range of Salmonella
contamination of pigs examined before or immediately after slaughter.
In general, analysis of mesenteric lymph nodes or some other organ
such as the spleen, rather than faeces, may provide the best index
of contamination of pigs with these organisms because those in
the blood and lymph are probably concentrated by filtration.
Analysis of excrement,either directly or by rectal swab, may fail
to recover low numbers of organisms particularly with animals that
are not persistent excretors (McCall et ai., 1966; Haddock, 1970).
Indeed some have advocated analysis of the previously noted organs
because they contend that there is a good correlation between
their contamination and that of the meat taken from the carcass
(Cherry et ai., 1943; Floyd et ai., 1953; Newell et ai., 1959).
Surveys such as those listed in Table 34 have directed attention
at two topics, the sources of Salmonella to which farm animals are
exposed and practices, both on the farm and in the factories, that
influence the incidence of Salmonella contamination.
Contaminated feed has been identified as an important primary
source of Salmonella (Adam, 1957; Walker, 1957). This view is
supported by the many surveys (e.g. Anon, 1959; Pomeray and
Grady, 1961; Schotts et ai., 1961), which have revealed a high
incidence of Salmonella in feeds or in ingredients used in their
production. In practice, feeds or ingredients appear to be sub
jected to sporadic rather than persistent contamination, contamination
of some rather than all batches is a common finding in these surveys
(e.g. Jacobs et ai., 1963; Clise and Swecker, 1965; Dawkins and

226.
p0)X
s
p(DXXG73GG
S0•HPawO'•HAG•HGO•HX
g•H
XG8GX0I3Xo(DUGG
73-PUGH
Ifmd)X64
û)> mX QJX rHX Û4ü) S0 GA W
dP
GüSSX0)05?G
Q)UPGOW
0)>XX enX (Dcn rH0 AQi
dP cn
(DüGQ)PGXCDP
73S<DUG
S
00■4 CN ro •o cr> X oX 'If ^ X LO
X
VÛ
tn 'f inX CN I
XCN [h
ro
enVÛ
CN O00 00en en1—1 X en X X enX in X 'sf X uo Xen en X LO en en X
X 1—1 X en en X X enen X X X X XP VÛ GG enW X X X X 1—1 X X
G G G G CD GX X
G X X G X X X GG (ü (U CD CD X >, CD
X G CD XS G >1 X en CD G >1 X 1—j CDG CD CD X Q) 0 CD X G iXU > > (D u G X > CD X CD •HCD G P ? G G X P « > GG P G CD G g G G 0 u d) enU U ffi 2 P4 u U ffi g s 2 H
cnG730G
ffi
1>1rH
cnü G•H CDP CDCD iHX AG cnCDen 73CD G2 G
uo Tf en X CNCN 6 X <f en ro o CN
ro X
enroenO rH
XenfH •H73Gp CDCD S
ro ffi G m'f U en "Sf CNen G X enrH S G X en
tH eo XCD04 73
X G XUO TT G G G X
X vn VÛ Gen en X CD Xen X X CD 73 ffi CD X1—1 G ü CD>1 G CD CDP G G GG G G P iH X CD
0 0 0 CD CD > ffiG G G ffi 73 0 CD P5 C C U M ffi ffi 05
VÛroen•H?0§mI— 1GW
%G
0)1 iS
P0ofHX
(D m en G G X
gG G X X X X
^ LD VÛ
wgIGXUOP
enCDüCDGX
73GG
enCD
730G
ffi1rH GînU GX Pp XCD GX fHGCD Gen XGs
en enX >1 >1 >1G G G GX 73 73 73p0 tH CN ro04+ H CN ro

227
Robertson, 1967; Harvey and Price, 1967). Thus from 7 - 34% of
vegetable ingredients, bone and meat and bone meal were found to
be contaminated in one survey (Anon, 1961) whereas Salmonella was
not detected in any of the samples of fish, cereal or vegetable
protein meals, and in 7% only of samples of meat and bone meal in
another (Patterson, 1969). The variety in the range of serotypes
isolated in these surveys is additional evidence of the sporadic
nature of the contamination. Single batches of feed have been
implicated in the introduction of Salmonella heidelberg and
Salmonella st. paul, for example, into swine herds (Newell at ai.,
1959; Galbraith at ai., 1961), and the Salmonella excretion by
symptomless carriers can be important in the subsequent dis
semination of the organisms (Sojka at ai., 1977). The use of
contaminated feed has been linked without dubiety to outbreaks
of food poisoning. The evidence supports therefore the notion
that the use of Salmonella-free feed could play an important
role in protecting humans from Salmonella of animal origin.
Indeed Kampelmacher at ai. (1965) did not isolate Salmonella from
slaughtered pigs which had been fed Salmonella-free feed.
Of the many methods that have been used to achieve feed of
this status, the use of heat during pelleting has attracted most2 5attention. This process can cause a 10 - 10 (Mossel at ai.,
1967) reduction in the Enterobacteriaceae count and the carrier
rate for Salmonella in pigs. Thus only 0.4% of pigs fed with
pelleted feed material compared with 4.7% receiving meal were
carriers (Edel at ai., 1973).
Stress associated with the transport of pigs from farms to

228,
markets or abattoirs can result in an increase in the number of
animals excreting Salmonella as indicated by the results of some
(e.g. Galton et al., 1954; Kampelmacher et al., 1953; Williams
and Newell, 1970) but not all surveys (e.g. Tsai et ai., 1971;
McKinley et ai., 1980). Similarly,residence times in the lairage
has been shown by some (e.g. McDonagh and Smith, 1958; Anderson
et ai., 1951; Bevan Jones et ai., 1964) but not all workers
(e.g. Craven and Hurst, 1982) to increase the incidence of pigs
excreting Salmonella.
Once pigs enter an abattoir, faeces (Childers et ai., 1973;
Stolle and Reuter, 1978), the skin and hair of the animals
(Norval, 1961; Patterson and Gibbs, 1978) and equipment (Lehmann,
1964; Watson, 1975) can become important vehicles in the dis
semination of Salmonella and operations such as scalding at 60 - o62 C or dehairing do not always diminish the incidence of
contamination of carcasses with these organisms (Kampelmacher
et ai., 1961; Chau et al., 1977). Indeed such practices (Fig. 1)
may contribute to Salmonella contamination (Galton et ai., 1954)
because the organisms are afforded protection by hair follicles
or faeces. This protection probably accounts for the frequent
occurrence of these organisms in the dregs in the scald tank
(Kampelmacher et ai., 1961) and effluent drains (Chau et ai.,
1977). Salmonella contamination of porcine carcasses can be
reduced by exposure to naked flames, but the process that is
used to remove the singed appearance, "black scraping", can cause
recontamination, (Kampelmacher et ai., 1961). The potential of
the abattoir environment to be an important depot of infection was

229
shown in a survey (Anon.) published in 1964. It reported a 1.9%
incidence of Salmonella contamination of pig tissue immediately
after slaughter but recovery rates of 74%, 11% and 0.5% from
swabs taken from drains in the abattoirs, meat factories and
butcher's shops handling the meat included in the survey. The
frequent occurrence of identical serotypes and phage types in
this particular survey provided further evidence in support of
the generally accepted view (McDonagh and Smith, 1958; Lo et ai.,
1967; Chau et ai., 1977) that a chain of infection can, under
unsatisfactory conditions, link the farmyard to the kitchen of
the home or canteen.
In attempts to rectify problems associated with traditional
slaughtering systems, attention has been directed at methods
that reduce the number of spoilage organisms as well as Salmonella
on carcasses that are about to enter chill rooms. Spraying the
carcasses with water containing acetate, stannous chloride, hydrogen
peroxide or steam (Biemuller et ai., 1973) as well as chlorine,
antibiotics, g-propiolactone, citrate and succinate have been
shown to be effective, at least under experimental conditions.
The purpose of this study was to establish the actual numbers of
Salmonella in sausages produced in a factory in which traditional
methods of slaughter and butchery ensured that meat used in the
product was liable to exposure to the several depots of infection
noted above.

230
Salmonella in sausages
Few attempts have been made to establish the level of con
tamination of sausages with Salmonella (Surkiewicz et al., 1972;
Patterson, 1959; Pain, 1981). Indeed the major survey (3309
samples) by Roberts et ai. (1975) as well as the minor ones
(Turnbull and Rose, 1982; Barrell, 1982) of British fresh
sausages could be criticized because of the emphasis given to
the incidence rather than the level of Salmonella contamination.
As it is generally accepted that, with the exception of chocolate
products (Gill et ai., 1983), at least 10^ Salmonella are required
to produce overt symptoms of disease in undebilitated humans
(Bryan, 1980), the level as well as the incidence of contamination
of a food should be considered to be of equal importance. It was
for these reasons that a quantitative method for the recovery of
Salmonella was adopted in this study.
Inadequate knowledge about the distribution of micro-organisms
within food and the inaccuracies inherent in the available methods
can give misleading information concerning the incidence and level
of contamination of food products with Salmonella, For example,
the assumption that a log-normal distribution of micro-organisms
exists in samples taken from a batch of food may fail to take
account of the skew in the distribution (Kilsby and Pugh, 1981).
At least 20 - 30 replicates/sample must be examined in order to
define this skew. If the variance of the log counts obtained
from such preliminary studies is reduced , however, the increase
in precision will allow a reduction in the number of replicates.
There is evidence also which suggests that micro-organisms in

231
general, and low numbers of Salmonella in particular, are distri
buted contagiously on whole pieces of meat (Kilsby and Pugh, 1981).
Thus thorough comminution is needed in order to achieve a random
distribution of the contaminants. Moreover as the variance of a
series of counts decreases, the apparent mean (average) of those
counts increases. In practice therefore examination of a sample of
whole meat may not yield isolates of Salmonella whereas they are
isolated from a comminuted sample. Similarly it has to be
recognised that a random but modest scheme of sampling of the
carcass may fail to detect one pig carcass which may have been
infected at slaughter with thousands of Salmonella. As butchering
and especially bowl chopping of sausage ingredients tend to random
ize the distribution of Salmonella, the probability of detection
of these organisms in samples of the finished product is increased
(Kilsby and Pugh, 1981). In the case of sausages, problems
associated with the sampling of ingredients such as meat can be
overcome by the use of multiple sub-sampling (Brown, 1977). If
the sample of finished product examined is considered in terms
of the contribution by individual ingredients, then it is evident
that very small samples of ingredients contribute to the bulk
sample (Table 35). This situation would be expected to favour
the isolation of Salmonella from (1) minced ingredients and
(2) ingredients which comprise a small proportion of the total
mix (e.g. rinds). Thus, fewer sub samples of these ingredients
were chosen from meats awaiting use in the factory and whole
pieces of meat (e.g. lean, fat) were minced (Kenwood A901)
in the laboratory to achieve a particle size similar to that
in factory-processed ingredients, (e.g. head, belly, rinds).

232.
2ffi2
73GC•H§gXCG•H73G4
•HXO
G0•HXUO
&klffi
G0
XXEG
S'w
cn
XO
G.3XXw0
1u73
§GNXG
GrHffi
X0GüG
giHXG
LD00
E-i
73
5S
GEGffiU G •G ffi en
G 00 e n (Ti cr» O2 X O O q CN CN inffi VÛ2 un ^ o o O o O>1ffi
73GG
X
XG
XGGX73 G ^G ffi enU G un 00 r~ r- Offi) X O —1 iH in un OG CNX O <—• O o O O iHXdP
CN o O O o*
en en VÛ rH H CNffi r~ VÛ CN CN »H
XX O q q oVÛ CN 'cr
dP CN CN
2S 2
X ffiG ffi kl ffiG kl 0 kl 2X 0 ffi 073 ffi S ffiG >1 Gkl G «H 73 73en G X fH G GG G G G G XH ffi ffi CQ 2 Pi
eoCN
OCN
Oro
SX0GGüGXffi
0§
îmffi
8ro
X0
XXs■gX
sG
S 'klG
SS
X
1GG<C
&SGX
G
73GUGX2

233
Even with all these precautions, experience showed that the level
of contamination was too low to permit enumeration by direct plating
and the MPN system had to be used.
Most Probable Number method
The most probable number (MPN) technique, a dilution method,
was developed to estimate the density of micro-organisms in a
liquid. Sub-samples of varying sizes of the liquid with several
replicates of each are analysed for the presence or absence of
particular micro-organisms using an appropriate medium for the
organisms sought. Three assumptions have to be made: (1) that
the micro-organisms being sought are distributed randomly
throughout the liquid, (2) that they do not attract or repel
each other, and (3) that growth in the medium inoculated with
a sub-sample of the bulk liquid will be initiated by one or
more micro-organisms. The probability of a negative result
(no growth) in a particular subsample replicate is related to
the density of micro-organisms in the original liquid thus:
where a subsample (v ml) is taken from the sample (V ml) in
which there are x organisms, the probability of any organism
being in the sample = v/V and of it not being in the sample
= (1 - v/V). The probability (p) that none of the organisms
are in the sample is p = (1 - v/V)* and when v/V is small,
it is approximately the same as p = e as x/V is the—vpdensity (p) of organisms per ml, p = e . If n samples,
each of volume v are taken, and if s of these are found to
be negative, the proportion s/n of samples is an estimate of p.

234
Therefore an estimate of the density (p) can be obtained by —vps/n = e which gives:
d = (- 1/v) £n (s/n) = -(2.303/v) log^^(s/n)
where d = the "most probable number" or organisms/ml.,
It follows that the precision of the MPN method is extremely
poor when all or none of the samples are negative since when
all are positive, the estimated density is infinite, and when
all are negative, it is zero, according to the confines or
threshold limits of the particular experimental design. So v
must be chosen so that some of the sub-samples are positive
and some negative. In practice, several levels of dilution are
chosen to achieve this end. Cochran (1950) showed that if a
fixed number of sub-samples is used, then the average precision
obtained is nearly the same for any dilution ratio between 2 and
10 although a low dilution ratio gives a more constant precision
over the range of densities being covered. Thus for a fixed
total number of samples, the choice of dilution ratio is a
compromise between the extra work involved with using several
levels of low dilution ratio and the fluctuation in precision
associated with a higher dilution ratio.
The number of replicates at each dilution level is important
also (Cochran, 1950). The standard error attached to an estimated
density (d) is usually computed from log^^d as this is closer
to a normal distribution than 'd' which is skewed. For a
dilution ratio of 10, the standard error of log^^d is:

235
S.E. (log^^d) = 0.58 / /n
where n is the number of replicate samples per dilution. The
precision of the MPN method can be estimated from plotting the
number of replicates/dilution level against the standard error
( l o g i o d ) .
The estimate of the MPN was obtained by reference to tables
(McCrady, 1915; de Man, 1977; 1983; Parnow, 1972) or computer
program. A program was written by Dr. A. Robinson (School of
Mathematics, University of Bath) for the present study such that
the MPN/g original material for any combination of replicates
or dilution levels could be calculated. At 3 levels of
dilution and a dilution ratio of 10,5 replicates/dilution were
done. Thus the detection level is theoretically 2 Salmonella/q.
Twenty complete batches were analysed.
Materials and Methods
Sampling and isolation of Salmonella ( 5 tube MPN scheme )
Samples of pork sausage, pork and beef sausage and all the
ingredients used in their manufacture were obtained from a large
factory during normal production in the period 2.3.80 to 5.4.81.
One kg of pork meat was taken from material awaiting processing.
The meat came from pigs slaughtered at the factory ; butchered
meat was stored at 4°C overnight. Cattle were neither slaughtered
nor butchered on the site and 1 kg samples were taken from
blocks of frozen,de-boned beef. Samples of rusks, seasoning,
polyphosphate (Fibrisol VlO), rinds and linked sausages were

236.
examined also. All samples were stored at 4°C and examined within
3 h of collection. To minimize sampling errors, all the meats
were comminuted with a sterile mincer (Kenwood A901). Sixty g
of samples and 540 ml of pre-enrichment broth (Appendix p. 330) ,
usually buffered peptone water (BPW) - were blended in a Colworth
Stomacher 400 (Seward, London) for 60 s. Five samples (each of
100 ml) of the homogenate were distributed in bottles. Five
samples (each of 10 ml) were added to 90 ml of sterile pre-enrich
ment broth in bottles and shaken for 10 s, and 5 samples (each of
1 ml) of homogenate were added to 99 ml of pre-enrichment broth
and shaken for 10 s.
Incubation was at 37* C for 24 h and after vigorous shaking
of the bottles, 1 ml samples were transferred to enrichment broth
(Appendix p. 330 )- usually tetrathionate (Difco) broth - (9 ml
amounts), and these subcultures incubated at 43°C for 48 h.
At 24 and 48 h, loopfuls of enrichment cultures were streaked
out on dried surfaces of Brilliant Green agar (BGA; Oxoid CM 329),
bismuth sulphite agar (BSA; Oxoid) and Desoxycholate citrate
agar (DCA; Difco) according to the methods of Edel and Kampelmacher
(1969). Inoculated selective agar in Petri dishes was incubated
at 37°C for 48 h; Colonies of presumptive Salmonella spp. were
transferred to Plate Count agar (PCA; Oxoid) after 24 and 48 h.
Characterization of Salmonella
Pure cultures on PCA were inoculated into Kohns I and II media
(Oxoid) and the API 20 E scheme (API, Basingstoke, England).

237
Isolates presumptively identified with Salmonella were characterized
further by the biochemical tests of Edwards and Ewing (1972) and
analysis of somatic and flagellar antigens using slide and tube
agglutinations with antisera from Burroughs Wellcome (Beckenham,
Kent). The MPN of Salmonella/q sample was established with
McCrady's (1915) tables or by reference to the computer program
of Dr . A. Robinson only after complete confirmation of isolates.
Determination of antibiotic resistance of Salmonella
Strains of Salmonella were incubated in Tryptone Soya broth
(TSB; Oxoid) at 37°C for 18 h and a sample (0.2 ml) spread over
the dried (37°c, 1 h) surface of Diagnostic sensitivity test
agar (Oxoid CM 261). Antibiotic-seeded "multodisks" (Oxoid) were
placed aseptically on the surface of the agar. Incubation was
at 37°C for 18 h.
Isolation of Enterobacteriaceae
Twenty g samples of minced meat, other ingredients and sausages
were homogenized (Colworth Stomacher) in 180 ml of quarter-strength
Ringers solution and decimal dilutions made in this solution. One
ml of appropriate dilutions was mixed with 15 ml of Violet Red Bile
Agar (VRB; Oxoid) cooled to 45°C. When set, the surface of the agar
was overlaid with 10 ml of VRB, incubated at 37°C for 24 h and
colonies of coliform organisms counted. One ml samples were
added also to 15 ml of Violet Red Bile Glucose Agar (VRBG; Oxoid)
cooled to 45°C. When the agar had set, its surface was overlaid

238
with 10 ml of VRBG, incubated at 30°C for 24 h, and the colonies
of Enterobacteriaceae counted. All counts were done in triplicate.
Randomly selected colonies from Petri dishes containing the
lowest, countable dilution were purified by plating on PCA. Gram-
negative, oxidase-negative, fermentative bacilli were tentatively
identified with Enterobacteriaceae. They were characterized
further by the methods of Edwards and Ewing (1972) and the API
20E and API 50CHE systems (API, Basingstoke).
Statistical analysis
A Hewlett-Packard 97 calculator and appropriate programes
were used to do simple linear regressions and Spearman's rank
correlation.
Results
The MPN scheme outlined in the Materials and Methods section
was adopted following extensive preliminary work in which comparison
was made with 0.1% (w/v) peptone water, lactose broth and buffered
peptone water for pre-enrichment (Table 36); selenite-cystine,
lysine iron cystine neutral red, and tetrathionate broths for
enrichment with incubation at 37° and 43°C (Table 37). A
combination of buffered peptone water, tetrathionate broth and
desoxycholate citrate or Brilliant green agar allowed best
recovery of Salmonella from samples of British fresh sausage
(Tables 36, 37). More isolations were made after 48 h compared
with 24 h enrichment and from enrichment at 43°C compared with

239.
uQ CN
E0UX
G Z 03M u POM HQ) ffiq G0 Xs 73M GG 603
O CX m ro G0 X
X>1 Gkl 1-4G ffi>0 XU G GG X u vr CN vo Gkl 73 û E
G ffiG E U0 X
en G kG G X G•H X G G
73 X GG G 0 XE X X 03 un «d’ uo 0ffi g OÛCT> X GG 73 G O•H G k XX G X XG G G1-4 X E-* ffiffi G p
G UG Ê U vO ro vO G> .5 ® X
X UX X ffio kG G COf-4 WGG k
073 U CN CN TfG ÛG CNX kG GG X
XG
U XX X 03 o o oy G CQ GG G XG X X
G G03 G
X 0G E2 -H- XG G
en Ü -k 03ü G m ro ro ^X G b ik G GG G XG G G1 XG rC Gkl G Xffi G G
kl o k 0X X 0 G U0 Xg ro G enX G !5 Gü X X 1—1G X G G ffiX X GX kl X 0a CÛ G k rG X en
G G X ffiX 0 G XG k ffi 0ü ^ ffiVO X 73 kro k G G G G
G G U) kG G 0 0 G1—4 1 X X Xa G ffi ü X 2G k G G GH ffi ffi ffi CQ ♦
73GXenGX
73en GG XX enffi G
X
w X0o 2
X
+ 4-

240.
GXXGG0GXGCOX0 0
ro>4kG> f f i0U COG TTk
G 00 X
row Gffi XX G0 Gk 0ffi X
f f iX XG G 0G k roX ^
GO 64X ffikGG CN
X0 0XG ro0X GX XG 73ffi GG EUG XX G 0
G roX S0 ffi
üG X f f ik kG G COX WGkG 0ffi +H- XE G roG tnX G
w73 G GG G XG tn X
GG ffi G0 en X 0X G G roX k CO 'vTG XkG ffi73 tn ffiXX X '(J'0 X CN
kX CQ 0U XG E roX 0X kW X
Xro enG EG X GrH X Xf f i G 73G X G64 eu E
VOrH
cnI—t
VO
CO CO
cn
CNX
cn
CO
♦00 VO
GXXGG
XGCOtnG
XG
XGXG0CJ
tn 73G GrH Xeu tn
GX
tntnX G
0 X&k E
G G■i tn
G CN2 ro
* -H -

241
37°c (Table 37).
Although Salmonella spp. were isolated frequently (Table 38)
from all the meat ingredients other than back-fact of pork sausages
as well as the finished product, the actual levels of contamination
were low (3 - 40 salmonellae/g ingredient). Thirty percent of
the cooked and comminuted rinds contained Salmonella spp. but
they were not isolated from the non-meat ingredients (rusk,
spices, polyphosphates) of pork sausage. Salmonella spp. were
isolated from all the meat ingredients other than pork back-fat
of pork and beef sausages (Table 39) but in low numbers only,
viz 8 - 40/g of beef flank and pigs' head meat.In contrast, 95%
of the mechanically recovered meat (MRM) contained 50 - 378
salmonellae/g. One sample of dried rinds but no samples of the
non-meat ingredients of pork and beef sausages yielded Salmonella
spp.
Of the 8 serotypes of Salmonella isolated from ingredients
and sausages, derby and dublin (Tables 40 and 41) occurred most
frequently. Salmonella agona was isolated on 4 occasions only
and S. infantis was the only serotype recovered from the cooked
rinds. The incidence of serotypes (7 out of the 8 isolated)
was highest with the mechanically recovered meat.
On no occasion was a serotype isolated from sausages without
it being recovered also from one or more of the ingredients used
to produce that particular batch. On several occasions, however,
serotypes were isolated from ingredients but not from sausages

242
m73c O VD O H CN•p CN tn M 1« kO
4J(Ü03Scto r~03 O 03 o m1—1 CN CN 11 r-4•H i-HE0303
M•P<D (0c 0)0 E 1—4E O CN o 0" <-1M 73 CN rH 1(Q to ro03 03Œx:-p•H
0) ■Pcn tofd 03u> E3 m«3 >1 O r' cn rH CNcn 1—1 CN m CN 1rH03p CQ0ftTlCn5 XPto 0 O-P ftC 03 VÛ o 103 c rH CN iH•H to iH03d> PIUftC•HU-t 0)0 ftto Oc U) O 03 o ^0 0 CN H CD CN 1•H to r~-V 03tl3C•H-P wC 03 030 iHu 73 -H p ft03 4J 0 E
pi -H tP tOW W 03 f t W03 0 > \ 0300 ••4-1 ft -H Z ft 03ro W 4J ft C >03 P P -H S to H03 ■H 03 03 to p 4-11-4 ft 43 ^ 0 C -HE E S ft to 2 to(T) to 3 3 03 ft QEh 03 d G dP S 2 ft
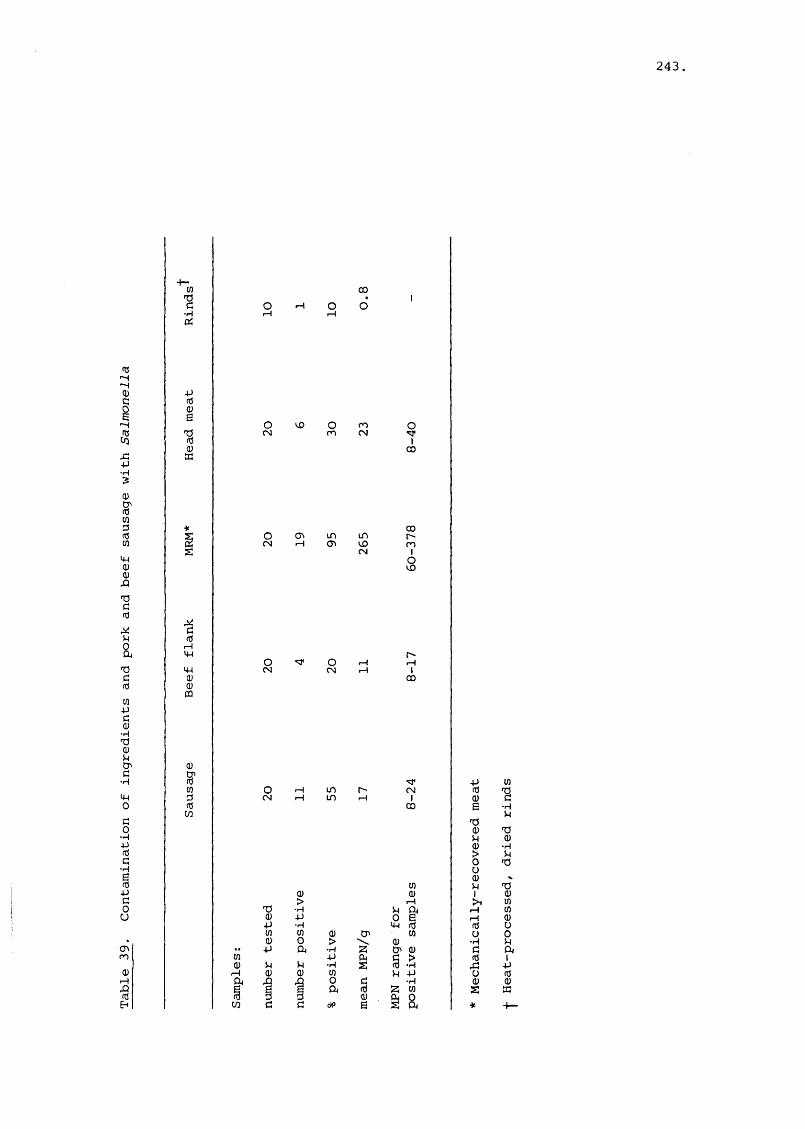
243
4—Ü) 0073 • 1G o <H o o•H r-4 1—1ft
<0MM03 PG G0 03e eM O CD O m otq 73 CM n CN ^03 G 103 00X ft-p•H303ft(0mD * 00(0 o ft CD ld r~-w CN P ft CD m2 CN 1PI O0) CD03A73GG
GP Gg 1-4ft p r-'O O P P73 p CN CN p 1G 03 00tO 03CQw■pG03•H7303Pft GG ft•H G P Wto O P LD r- CN G t)ip G CN p m p 1 03 G0 G 00 6 P03 PG 730 03 73•P P 0)P 03 PG > PG 0 73•H U03 "•ü) P 73p 03 03 1 0)G t> p >1 W0 73 p p ft p ü)U 03 P 0 e p 03P -H P G G UW U) Q) ft W U 00) 0 > \ 03 P Pft •• P f t p Z ft 03 G ftm W P ft G > G 103 P P P 2 G P Xi P03 P 03 0) W P P U Gp ft A A 0 G P 03 03-Q S S S ft G Z w 2 mG G 3 3 <U ft 0Eh 03 G G o¥> e 2 ft * 4—

244
(Uf tGm
Sco
f t in
co73CP05
03f tGW
3Oî
XP
f t
73§WP
5p
%p
p
iaG
PP03
IPccp0w
§p
Gr-4
8p
G
3N
6W
§
EH
PG03eGG03PIP§03
Ge'B03m
‘iQ)E> 1
XP
f t
00
Gf t>1P0P f t0) Pm 03 (3)
P 4 f t pG G P P 03 pp f t P 0 03 P Gp 4 P f t 73 G GG P rG P G <PG 03 3 03 G P Gi 73 73 G •G (3) PpG03
IP
p
IP IG

245.
03
WG
SP0303A
G•af t
73CGœPcG• H
Uf t.5epp
GPPG
igpow
§• ppG
P0tn•HGPG
S
g-en
p
E-t
G
S ’tn
Sen
w
p05
4«5GGP
P
PGGm
GGe
'BGm
+
I
f t CD C D 'd* COm
C N C D
00 m Om
ap0pGtn
Gpp
1pén
f t
73
P
IG
E P 733 G
P f t >1 toP P 1— 1 to3 G to 1— 1 G
f t E •Q P G UG G 1 P P U 0P • H p G G P PG P -G 73 G G f tG 4 f t P ip G 1P 3 f t G G Xi P(0 7 3 P •G P U G
G G2 X
G
e73G
g
ëu
co73GPP
%PP73

246.
containing the ingredients. In such instances, the level of con
tamination was low or the ingredient was used in small amounts only.
As the level of Salmonella-contamination of ingredients and
the letter's contribution to the sausage were known, it was possible
to compare the predicted and the observed levels of contamination
of pork sausages providing there were at least 2 of these organisms/g
of ingredient. With this type of sausage, the predicted contribution
by all ingredients and the observed level of contamination were in
accord (Fig. 71a) a linear regression line described the data with2a coefficient of determination, r - 0.95. In addition there was
2a good correlation (r = 0.92) of the predicted contribution by
lean pork alone and the observed levels of Saimoneila-contamination2of sausages. As all the other meat ingredients gave low r values
(footnote to Fig. 71) it was concluded that their contribution to
5aImoneiIa-contamination of pork sausages were subordinate to that
of lean pork, the principal meat ingredient. An acceptable corre- 2lation (r =0.79) of the predicted and observed contamination of
ingredients and pork and beef sausages respectively was also noted
(Fig. 71b). In this instance, the threshold level of contamination
of an ingredient was 5 salmonellae/g (cf. 2/g for pork sausages).
The observations summarized in Fig. 71 direct attention at
the role of dilution in diminishing the level of contamination of
a product by an ingredient that is relatively heavily contaminated.
This was particularly notable with MRM, the ingredient of pork
and beef sausages with the highest level of Saimoneiia-contamination
(Table 39)and the greatest variety of serotypes (Table 41). In

247.
practice, the extent of dilution of this material was such that2there was a poor correlation (r = 0.49) of the predicted and
observed levels of contamination of a product.
Enterobacteriaceae and coliform organisms were present in all
the samples of meat and sausages but none of the non-meat
ingredients of pork sausages (Table 42). There was little variation
in the levels of contamination of sausages with these organisms
but a pronounced variation with ingredients, especially whole pieces
of meat (Fig. 72). It was concluded that the former reflected
random contamination resulting from the vigorous mixing of
ingredients in sausage production and the latter contagious con
tamination resulting from contact of meat with dirty equipment
etc. In addition, dilution of contaminants was again noted. Thus
MRM, the ingredient containing the largest number of coliform
organisms, was used only in pork and beef sausages which contained
fewer of these organisms than pork sausages. Only one sample of
a dry ingredient, rinds, of pork and beef sausages contained coli-
forms and this was the sample from which S. typhimurium was iso
lated (Table 43).
In general, the Enterobacteriaceae count (VRBG medium) appeared
to be a more reliable index of possible Salmonelia-contaraination
of sausages and ingredients than was the coliform count (VRB
medium). This is evident in the results obtained with the lean,
semi-lean pork and belly meat used in pork sausages (Fig. 72b).
There was no apparent association between the level of contamination
of head meat, cooked rinds and pork and beef sausages with

248
M
gg•HP0 u
'SG
GGGUG•HPGPüG
ggP
a5•H
Gg-W3G01
•g
â73
§01P
g•H73GP
-p
p0G0•H
GG-P
PG0UPO
P
GP
â
CM
GrH
•§Eh
f t
G
P
ü
f tO
I Xa•PrHou
ft\
G
P
U
ft
G•PPGP
S镧PG
S
%G
P p
I X
WG
f-Hft
O r o f t CN VD CNrH M CN m en e n
O O 6 O O 6 6
CN CD CD 1—1 p f tf t f t P en co 'ey f t
CN CN en en CN CN p
o p CD r - b ' in coI—i en en en CN 'ey en
o d d d d d d
CD p CN •ey CN 00CN o co CD n* p LO
en 'ey en en en en CN
s LOP
OCM OCN
OCN OCN
OCN
f t
en
PGGS
P,x G P Gp G G G
G 0 G G ef t f t P SG 1 >1 en01 G P 73 1—1 73G G S G P GG G G G G pen kG en K CQ D5 G
f t
01GPGUPP
»PGG
PPO
G
e
I X
01G
UPPf t
2GG
PP0G0 P P G P
G73
73P
§P01
11b

249
wg0pp1—40u?G
GPUGP
%P0PG
W
XP
■SGftGWGGWPGG
P
73§
yft73g01pGGP%ë .GPP0§PPGGPE5G0üP0P
G
W
ro
GE-
ft
P CN m Lfl cy> 1—1• G 1—4 CN ro en
U o O O O OO
gr—I
01
g0PP I Xf—I 0 u
G
P
S’
GPPGPU
A l X
PGP
S
%p01GP p
001
P G G P
1 IZ 01
«cr'y
CM
ro
OCN
en CD P ft *00
ro CN 00 CD
ro p CN 1—1 6
ft UO CN 00I—1 CN UO ■G*
d d d d
f t O r~ f t■teO
00 00 CN f t CD
ro ro ro 1—1 d
OCN
OCN
OCN OCN O 1—I
PG
>1 G1—1 E1—1 AG 73 P GU G G G
G P P G r—iG f t G G S P
1—4 G G > 01f t 01 A 0 A P 73
G Ü U G G P GG G G G G G Pen en 2 P A A ft f t
d)•SPP0ft
g
üft
01G
üP1—4ftGP
GG
APPO
gE
I X
01GPGUP
P
S'P
GG
5p0
gPP■S1
73P
5

250,
Enterobacteriaceae (Fig. 72b). As would be expected, Entero
bacteriaceae in a heat-processed, dry material such as the rinds
used in pork and beef sausages provides evidence not only of post
processing contamination but, judging frc*n these observations
(Table 43), a warning that food poisoning organisms such as Salmonella
may have gained access to the product. Salmonella spp. and
members of the ’Arizona* group isolated from sausages and mechanically
recovered meat during the preliminary and main experiments were
screened for their sensitivity to a variety of antibiotics. All
of the strains tested were resistant to at least one of the
antibiotics used in the challenge test (Table 44), many were
multiply-resistant and one strain of Salmonella dublin grew in
the presence of 5 antibiotics. The majority of strains (88%)
were resistant to benzylpenicillin (Penicillin G) and chloramphenicol
(50%) and appreciable proportions (43% and 33%) to oxytetracycline
and sulphafurazole respectively. All strains were sensitive to
erythromycin, streptomycin, furozolidine, nalidixin and cyclo-
heximide. Although insufficient numbers of organisms were tested
to indicate clear patterns of resistance, some trends were evident;
Arizona organisms were all described by the profile CH^,P^,OT^ and
Salmonella infantis, P^.
In order to achieve an overall perspective of Salmonella
contamination of a commodity such as British fresh sausage, the
results of surveys of the finished product must be considered
together with observations on the precision of quantitative methods,
variations due to season and causes of contamination of ingredients.
Finally, of course, the behaviour of the food poisoning organism

251
GftGWGGm73GG
m-pGG
•H73GU
•H
G
S'w
Seneuiw
73GAGrH0en
•H
GMMGG
5m0GU
g4->en
•HGGP
O•H
•HA•H
<c
Eh
U
EhO
G W 73 0 Uü
■H
■SA•HAG<
ft
M
U
GM
GG
M
+ +
+ +4- 4- 4-4-4- -f-E 4--f--+- 4--E 4-
4- 4- 4- 4 4 4- 4 4 4 4
4 4
iH r-4 tH I—I CN
4 4 4 4 4 4
r-4 1-4 t—I t—I CN CN
4 4
CN CN CN
4 4
CNA 73 vO CN m 1—t CN f t ro in vo O CN f t 'ey ID CN r-' œ 1-4 C" CN rorH 0 vD o 'ey VD ro CN CN 1—1 kO in O O '—1 CN CN O O O OG U 03 03 03 03 03 03 03 03 03 03 03 03 03 03 03 03 03 03 03 03 03
G
IG
G•H
A
%
A
y&g
GiG
MGGAtn

252
u
g
ft
u
A
%GG
•HAG0U
■G*
«Eh
GftGCO
e G0 G rH Gp (0 0 G GA u iH AA •H G 0 U G G A73 G G G •H N G G üG ft G G O G G •H •H >iA G A f t >1 U •H rH 73 G UG CO ft e G r—1 O •H •H Gr—j G 73 g 0 A 1—1 >1 1—1 X A0 G G G u G •H U 0 •H Aw CO G kl A A U G N 73 GH 0 A f t •H A G •H A
f t ft A >1 1—1 G A U A >1U U f t U G G G G G Xp P U G CO ft A A G 0
>1 ft ft SG K f t f t Ehft A CN ro U W 03 f t Eh f t Z O
G73•H
•g
Ir4üS’u
G•H
IA(0
W
+ + + + + + + + + +
+ +
+ + + + 4 + 4 4 4 4 4 4 4 4 4 4 4 4 4
U 4 4 4 4 4 4 4 4 4 4 4 O
G
A r— CN (N r—1 A A A ro ro AA 73 O ro A CO C73 m ro CN ro r - r- r - A A o mt l Ps O CN 'T ro O CN ro f t Ü Ü <C f t u Q w o ffi 7)S "
03 03 03 03 03 03 03 03 03 03 03 03 03 03 03 03 03 03 03
U)G
G f t •HA A P G AA k G CO G A A A GG 3 A A G 73 f t f t f t f t ÇJ U Q Q A GG S A A 0 G W A
A G G N f t f t . COA 73 G A >1 G A AA f t A ft| k A 0 0 WG f t G G <5 G A G03 A A A D Ü dP A
ro4*
COCO
roro

253.
in a product post-manufacture has to be considered also. As the
MPN method had to be used to enumerate Salmonella, the number of
replicates per dilution and the precision of the methods have to
be considered in the context of the work involved. Indeed the
number of samples that had to be examined dictated the use of
5 tubes per dilution in the work discussed. As the standard error
of the logarithm of the MPN is inversely proportional to the
square root of the number of tubes at a single dilution (Cochran,
1950), it might be anticipated that a higher incidence, and maybe
a higher level, of Salmonella contamination of sausages would be
demonstrated by doubling the number of tubes per dilution. In
practice, there was no appreciable difference in the results
presented above and those obtained in a limited survey (Pain, 1981)
using 10 tubes/dilution. Indeed, as the latter survey was done in
the late autumn/winter, it showed a diminishing incidence of con
tamination of ingredients and sausages with Salmonella and thus
confirmed the many observations (e.g. Roberts et al., 1975) that
ambient temperature influences Salmonella contamination of meat
and meat products. Additional confidence in the MPN method was
given by the observations that a serotype recovered from sausages
was invariably isolated also from one or more of the ingredients.
Apart from dissemination of Salmonella spp. of gut origin through
a pigs tissues at slaughter (Kampelmacher et al., 1961), puncturing
of the gut at evisceration can be an important cause of contamin
ation of a particular carcass (Kampelmacher et al., 1961) and a
source for cross contamination in subsequent processing
(Kampelmacher et al., 1961; Chau et al., 1977). Indeed, it has
been shown that a 'comet tail' type of spread of Salmonella follows

254,
the processing of a contaminated carcass until the organisms are
eventually diluted out from the system. In this instance,
contagious (Elliott, 1977) rather than random distribution will
occur. In other instances, a process, such as the washing and
de-hairing of pigs' carcasses or the mincing of material obtained
therefrom, can provide not only a longer term source of infection
but also opportunities for the contaminants to be randomly rather
than contagiously distributed in or on meat or meat products
(Elliott, 1977). The influence of mincing or some other form of
comminution on the distribution of contaminants has been demon
strated by Kilsby and Pugh (1981). Moreover, the variation in
the viable counts of organisms, be they Salmonella or coliforms,
decreases as the contaminants become more randomly distributed.
Conversely the apparent mean of the counts will increase with a
more random spread of organisms. These influences were noted
in this work also and account for the differences between the ob
served levels of contamination of the major sources of Salmonella
and the predicted level in the final product (Fig. 71). The present
work has shown also that the influence of dilution must be taken
into account when considering the relative importance of ingredients
as causes of contamination of a product. Thus although MRM
contained the largest numbers and greatest variety of Salmonella
spp., the use of small amounts (e.g. 1.7% of sausages) meant that
its contribution to the contamination of the product was negligible
when compared with belly meat (cf. Table 39). Indeed, the latter
would appear to be the ingredient that ought to be monitored
routinely in sausage manufacture when the objective was to
minimize contamination of the product with Salmonella. The viable

255.
counts of Enterobacteriaceae appeared to be poor indices of
Salmonella contamination of the majority of uncooked ingredients
included in this study (head meat and backfat in pork sausage
and MRM, head meat, beef flank and backfat in pork and beef
sausage). From the viewpoint of management in a factory, they
are of particular value as would be expected in the routine
examination of processed ingredients. Thus there was a good
correlation of Enterobacteriaceae contamination of rinds and
dried rinds with the recovery of Salmonella. Moreover, the
isolation of Salmonella infantis from one ingredient only, the
cooked rinds, may indicate that a human carrier rather than pigs
was the source of this serotype.
Although the survey of Roberts et ai. (1975) included both
sulphited and unsulphited sausages, the small number of samples
(312) of the latter did not permit an assessment of the possible
influence of the preservative on Salmonella in the commodity mainly
because the survey was qualitative in nature. Our observations
that the actual was invariably greater than the predicted level
of contamination of sausages with Salmonella can be taken in part
as evidence that sulphite had little if any bactericidal action on
these organisms during the manufacturing stage. Moreover, the low
and unpredictable levels of contamination of sausages with
Salmonella means that samples taken from a factory cannot be
used in studies of the fate of these organisms during storage.

256.Figure 71. Linear regression of predicted number of Salmonella from
ingredients with number of Salmonella in a, pork sausage and b; pork and beef sausage.
a Regression coefficients (r ) in pork sausage. All ingredients, 0.95; Lean meat, 0.92; Semi-lean meat, 0.44; Rinds, 0.07;Head meat, 0.03; Belly meat, 0.15 (data points not shown)
All ingredients Lean meat
# #••
Q j
Salmonella / g sausage
b Regression coefficients (r ) in pork and beef sausage. All ingredients, 0.79; M.R.M. 0.49; Beef flank, 0.20; Head meat, 0.42 (data points not shown)
czn- Î 2
(Uc:o
aC o
' b
" b
tt D
Q j
qj
All ingredients
6’512
3-256
L #L2 4
Salmonella ! g sausage

257Figure 72. Relationship between numbers of Enterobacteriaceae and
presence of Salmonella in ingredients of a,pork and beef and b, pork sausage
OO00: 0
OqOqp OO ) OO
FAT RINDS SAUSAGEBEEFHEADM R M
I II
«5
5
4
od3OO
OoOOOp2
LEAN SEMI-LEAN BELLY HEAD RINDS FAT SAUSAGE50 g sample negative for Salmonella
60 g sample positive for Salmonella

258,
PSEUDOMONAS
The importance of members of the genus Pseudomonas in the spoilage
of proteinaceous foods stored at chill temperatures was recognised
slowly, a situation due in part to the inadequate descriptions of
the genus (e.g. Bergey et al., 1939; Breed et al., 1948). The
major taxonomic study of the aerobic pseudomonads by Stanier et al.
(1966) marked the beginning of a new approach to the classification
of these organisms even though initially the wealth of data produced
by such studies were difficult to analyse routinely. Indeed food
microbiologists continued to rely on simple keys (e.g. Shewan et
ai., 1960a) to identify their strains. Computer-aided numerical
taxonomy has negated the difficulties associated with the analysis
of large amounts of data, affirmed the significant contribution
made by the pseudomonads to the microbial populations on meat and
in meat products and enabled the identification of the principal
species.
Historical perspective
The first description of the genus Pseudomonas (Migula, 1894)
gave emphasis to the presence and location of flagella as well
as to general morphology. As the former criteria were not accepted
generally by bacteriologists, misidentification of micro-organisms
led to considerable confusion and contradiction in the literature
relating to foods.

259.
Early studies with meat isolates
Glage (1901) was perhaps the first person to note that motile
aerobic bacilli capable of liquefying gelatin occurred in the
slime developing on the surface of chilled meat. In the 1st
edition of Bergeys Manual the Gram-negative saprophytes were
sub-divided on the basis of pigment production without any regard
to the presence or location of flagella. Thus,organisms producing
yellow pigments were assigned to flavobacteria, green pigments to
pseudomonads and no pigments to achromobacters. These traits
were reflected in subsequent studies of the aerobic organisms
recovered from meat. Thus,Haines (1933), who studied the bacterial
flora which developed on chilled lean meat, identified the majority
of isolates with Achromohacter because they failed to produce pigment.
Haines and Smith (1933) referred to the principal component of the
"microbial association" on spoiled meat as the Pseudomonas/Achromohacter
complex, a term that was still in use in 1957 (Rogers and McCleskey).
The implicit weakness of basing classification on pigmentation alone
has been highlighted repeatedly. Thus Ingram (1934) reported that
all his isolates from pork mince were pseudomonads whereas Empey
and Vickery (1933) and Empey and Scott (1939) identified the
majority of their meat isolates with Achromohacter. Kluyver
and van Neil (1936) can be considered to have been instrumental
in causing a fundamental change in the classification of the
Pseudomonadaceae; they emphasised the taxonomic importance of
polar flagella (Bergey et al., 1939). Even so, reorganization of
the 5th edition of the Manual included 19 such organisms in
the genus Achromohacter. These organisms were later transferred
to the genus Pseudomonas (Breed et al., 1948).

260,
The failure of food microbiologists to accord taxonomic
significance to the presence and location of flagella led to per
sistent misidentification. Thus, although Brown and Weidemann
(1958) examined the isolates of Empey and his collaborators, and
re-identified the majority with Pseudomonas, the view that Achromo
hacter spp. were important spoilage organisms persisted. For
example, despite the implication of members of the genus Pseudomonas
in the production of off odours and slime on flesh meat (Jensen,
1944), some contemporary reports (Jepsen, 1947) suggested that
Achromohacter spp. were the predominant organisms in the slime.
Although the 6th edition of the Manual added to the number of
species in the Pseudomonadaceae, it provided only vague clues for
the identification of pseudomonads from food. Thus, Sulzbacher
(1950) was unable to identify organisms from frozen lamb/pork
with the species definitions in this edition of the Manual. Indeed,
of the 541 isolates of Pseudomonas from refrigerated beefburger.
Kirsch and his co-workers (1952) could identify a minority only,
147 with Pseudomonas geniculata, 88 with Ps. fragi, 62 with
Ps. rugosa and 9 with Ps. aeruginosa. The lack of suitable
criteria in the Manual for the identification of the unpigmented
species of Pseudomonas also resulted in a tendency to over-estimate
the importance of the pigmented types. For example, Ingram and
Hobbs (1954) identified all the pseudomonads from cured and un
cured hams with Ps. fluorescens and Kitchell and Ingram (1956)
the dominant species on slimy pork with Ps. multistriata. Like
wise, Wolin and his collaborators (1957) identified 95% of these
isolates with Pseudomonas geniculata. The need for further

261
systematic study of species within the Pseudomonadaceae was stressed
by Ayres (1960a) who, from a review of the literature in which
identification had been based on definitions given in the 6th
edition of the Bergey Manual (Breed et al., 1948), noted that at
least seven species (Ps. fluorescens, fragi, putida, ambigua, con-
vexa, taetrolens arid incognita) had been associated with the
spoilage of proteinaceous foods. The need for a simple, utili
tarian scheme for the identification of pseudomonads led Shewan
and his co-workers (1960 a) to devise a key which proved to have
most value in the classification of aerobic Gram-negative micro-organisms iso
lated from fish(Shewan et ai.,1960b).The inadequacies of the descriptions of Pseudomonas in Bergey's Manual can be considered as the major
cause of the dubious identifications noted above as well as the
reason for many microbiologists adopting routinely the scheme
of Shewan and his collaborators (1960 a).
A major, but in some ways, a traditional study of the taxonomy
of Pseudomonas was done by Stanier and his co-workers (1966). They
based their definitions of species and biotypes on the ability of
an organism to use a specific pattern of compounds as carbon and
energy sources, an approach inspired by the work of den Dooren
de Jong (1926). It was notable, however, that Stanier did not
favour the use of computer analysis; subsequent studies with this
method (Sneath et al., 1981) were in accord with the major trends
proposed by the original workers. Davidson and his co-workers
(1973), who adopted the methods of Stanier et ai. (1966) to study
231 pseudomonads isolated from meat, reported a very low incidence
of identification of their isolates - 16 were identified with Ps.

262
fluorescens, 19 with Ps. putida - with the descriptions given by
Stanier et al. (1965). The majority of isolates were non-
fluorescent and non-pigmented - the Group II of Shewan et al. (1960a).
Similar difficulties were encountered in studies of isolates from
milk (Juffs, 1973) and fish (Gillespie and Macrae, 1975; Gray and
Stewart, 1980; Gillespie, 1981). It is notable,however, that
Gyllenberg et ai. (1963), who applied computer analysis to isolates
characterized by traditional methods, recognized Pseudomonas fragi
as an important spoilage organism of milk. This organism, first
described by Eichholz (1902) and GrUber (1902), had long been
regarded as an important dairy organism (Hussong et ai., 1937) and
is now recognised as the dominant pseudomonad on chilled meats.
Thus it accounted for 67% of the isolates which Molin (1981)
studied, an observation which has been confirmed by others (Martin
and Patterson, 1982; Shaw and Latty, 1982; Molin and Ternstrom,
1982).
Sources of Pseudomonas
Early reports suggested that hides and wash water were the
most important sources of carcass contamination with psychrotrophic
micro-organisms (Haines, 1933; Empey and Scott, 1939), and the
isolation of Pseudomonas from dressed carcasses has led to suggest
ions that the hide is important in the transfer of organisms to
meat (Nottingham et al., 1974). As psychrotrophic strains of
Pseudomonas are common in soil, water and on vegetation, many
opportunities exist also for their transfer to meat via the factory
workers, their cutting knives or machinery. The walls and floors

263.
in chill rooms and cutting boards in boning rooms also harbour
substantial populations of pseudomonads (West et al., 1972; Newton
et al., 1978). Although structural and work surfaces might be
expected to spread organisms onto meat, the actual level of
psychrotroph contamination on meat has been shown to decrease
during the butchering stage (Newton et al., 1978). The seasonal
fluctuations in the level of psychrotroph contamination of meat
have been attributed mainly to changes in ambient temperature
(Empey and Scott, 1939), and rainfall (Newton et ai., 1978). The
low temperature and high relative humidity in chill rooms are
factors which are considered to favour pseudomonad growth and the
ratio of the psychrotroph count to that of the total count on
porcine carcasses increases with chilled storage (Scholfield et al.,
1981). The present study has associated high levels of contami
nation of sausages with prolonged chill-room storage of meat
(p. 165). Although it is generally accepted that Pseudo
monas spp. are the dominant micro-organisms on whole pieces of
chilled carcass meat, the behaviour of this group in meat products
has been seldom reported. As the pseudomonads generally comprise
a fraction only of the microbial associations which develop in
meat products, a major cause of the failure to monitor the fate
of this group of micro-organisms has been the lack of a suitable
selective medium. Indeed, the cetrimide-fusidic acid-cephalodrine
medium of Mead and Adams (1977) appears to be the only one that
has been recommended (Gardner, 1980) for use in meat microbiology.
Thus the early reports by Dyett and Shelley (1962, 1966) and
Dowdell and Board (1967, 1968) did not comment on the behaviour
of pseudomonads in British fresh sausage although the last
mentioned workers demonstrated that this group of organisms formed

264
a significant proportion of the microbial association in heavily
contaminated sausages made by local butchers (Dowdell and Board,
1971). They noted also that pseudomonads multiplied in sulphited
sausage which had been stored at 4°C or room temperature. Plate
count agar, as used by Dowdell and Board (1971), was supplemented
with hexadecyltriammonium bromide by Ashworth et al, (1974). They
found that pseudomonads grew well in unsulphited but not sulphited
sausage at refrigeration temperatures. Brown (1977), who used the
medium of Masurovsky et al. (1963), confirmed these findings, and
noted also that the pseudomonads in sulphited sausage did not
die during storage at 22° or 4°C.
This section of the thesis is concerned with a detailed
characterization of Pseudomonas spp.; the behaviour of these
organisms in sausages is considered on pp. 125 - 132 and their
tolerance of sulphite on pp. 151 et seg.
The classification of pseudomonads and other aerobic Gram-negative
bacteria from sausage
The above review has shown that the classification of Pseudomonas
spp. isolated from chilled proteinaceous foods has tended to lag
behind developments in the taxonomy of the genus. There has been
a trend also for food microbiologists to isolate from the "blooms"
which develop during the storage of such foods. As this can be
considered to be a form of enrichment culture, the study of Shaw
and Latty (1982), for example, may have been inadvertently biased
by the dominance of enriched organisms. This possibility was

265,
avoided in the present study simply by using isolates taken from
meat stored in a factory at chill temperatures for a short while
only, or from stored sausage.
Materials and Methods
Sampling and Isolation
Samples (1 kg) of pork were obtained from meats awaiting use
in the factory - the pigs were slaughtered on site and the butchered
meat stored at ca. 4°C for 24 h. Samples of sulphited and un
sulphited British fresh sausage were taken also. All samples were
stored at ca. 4°C and examined within 3 h. To achieve random
rather than contagious distribution of the micro-organisms
(Kilsby and Pugh, 1981), whole pieces of meat were minced
(Kenwood A901, England). Samples (20 g) of minced meat or sausage
were added to h strength Ringers diluent (180 ml) and homogenized
in a Colworth Stomacher 400 (Seward, London) for 60 s. Serial
dilutions were made in h strength Ringers (90 ml) and 0.2 ml of
an appropriate dilution spread over the dried surface of Cetrimide
Fusidin-Cephalodrine (CFG; Mead and Adams, 1977) medium. Incubation
was at 15°C for 48 h.
One hundred and sixty-five strains of Gram-negative aerobic
bacteria were isolated from 20 samples of sausage ingredients and
the corresponding freshly-prepared sausage and those stored at 4°C,
10°, 15° or 20°C for up to 8 days. Colonies were picked at
random from the lowest countable dilution on CFG medium, purified
by restreaking on Plate Count agar (PGA) and maintained on slopes

266
of PCA at ca. 4°c with sub-culture every 4 weeks.
Preliminary characterization
Isolates on CFG agar were examined for colony form and pigment
ation; their Gram reaction, (Kopeloff and Beerman, 1922), cell
morphology, flagella arrangement (Mayfield and Inniss, 1977) and
oxidase reaction, (Kovacs, 1956) were determined also. Production
of acid from glucose was tested in the 0-F medium (incubation,
15°G for 10 days) of Hugh and Leifson (1953).
Culture characteristics
Pigment production; Cultures were streaked onto the medium of
King et al. (1954) and incubated at 15°G for 7 days.
Accumulation of poly B hydroxybutyrate: The method of Stanier et
al. (1966) was used.
Growth temperatures: Ability to grow at 41°G in 48 h, 30°G in 7 d,
15°G in 10 d and at 4°G in 14 d was tested in the YE medium of
Stanier et al. (1966); 0.05 ml of culture grown at 15°G for 48 h
in YE medium was used as inoculum.
Growth factor requirement: The ability to grow on the YEA basal
medium of Stanier et al. (1966) containing acetate (0.1% w/v) was

267
tested at 30°C for 7 d. Failure to grow was assumed to be due to
a deficiency in growth factors.
Denitrification: The method of Stanier et al. (1966) was used.
Biochemical tests
Catalase: Hydrogen peroxide (20 vol.) was added to cultures grown
on Nutrient agar containing glucose (1% w/v) at 15°C for 48 h.
Tween 80 hydrolysis: The method of Sierra (1957) was used (in
cubation, 30°C for 7 days).
Starch hydrolysis: The method of Stanier et ai. (1966) was used
(incubation, 30°C for 7 days).
Gelatin hydrolysis: The. method of Frazier (1926) was used (in
cubation, 30°C for 7 days).
Ornithine and lysine decarboxylation: The method of Miller (1955)
was used (incubation, 15°C for 7 d).
Egg yolk reaction: The method described by Anon. (1957) was used.
Carbon sources
The ability to grow on 126 compounds as the sole source of

268,
carbon was tested. The methods and media of Stanier at ai. (1966)
were used with the exceptions that (1) all compounds were filter-
sterilised, and (2) isolates were suspended in sterile h strength
Ringers and ca. 1 yl was placed on the surface of dried solid medium
containing a specific substrate by multipoint inoculation (Denley,
England). Incubation was at 30°C and growth was scored (at 1, 3, 5
and 7 days), as strongly positive (2+) or weakly positive (1+)
if growth was visibly greater than on the control medium. If growth
was equal to or less than that on the control medium, it was scored
as negative (0-). All the tests were repeated on 10% of the
isolates and the average probability of error (p) was calculated
(Sneath and Johnston, 1972).
Computer analysis
Multistate characters were analysed using the Taxpak program
(M.J. Sackin, Department of Microbiology, University of Leicester,
England). The Gower (S ), Simple Matching (S ) and Jaccard (S )G SM Jcoefficients together with unweighted pair-group average (UPGMA),
and single linkage clustering were calculated on the ICL 1900
computer at the University of Nottingham, England. Data were
analysed by the Ovclust program also (Sneath, 1979). A program,
DICHAR (Sneath, 1980 ) was used to list the most diagnostic
properties of the phenons and these results form the basis of
the key to the identification of clusters (Fig. 74 ) .
Results
The majority of the isolates were Gram-negative, non-fermentative,

269
oxidase-positive, motile bacilli with polar flagella and were
identified with Pseudomonas, A small number of Gram-negative,
oxidase-positive, non-motile bacteria were identified with Moraxella.
Both groups of organisms were included in the computer analysis
as were type strains of Pseudomonas, pseudomonads implicated in
the spoilage of pork and organisms from the genera Alteromonas,
Acinetohacter and Aeromonas (Table 45). All the isolates were
tested for their ability to use a range of substrates as carbon
sources. Ten percent of the isolates were re-tested and the
reproducibility calculated (Sneath and Johnston, 1972). As the
average probability of error (p) was ca. 3%, no appreciable dis
tortion in the computer analysis of the results was to be expected.
The Sg^ coefficient (matrix not shown) and clustering by UPGMA
linkage sorted 138 operational taxonomic units (OTU's) into six
clusters, A-F (Fig. 73 at the 77% level of similarity (77% S).
Ten of the 13 stock cultures were loosely clustered; one (Ps.
fluorescens NCIB 9046) clustered with A, another (Ps. fragi
NCIB 8542) with C, and the third {Alteromonas putrefaciens NCIB
10471) with group II (Fig. 73). The remaining OTU's (25) were either
scattered among the stock cultures or contained in groups I
(OTU's 15-18) or III (16 isolates).
Cophenetic discrepancies, the contributions made by each
strain to the distortion between (1) the similarities in the
similarity matrix and (2) the corresponding values implied by the
phenogram (dendrogram), were measured by:
%
(s... - c, , . ) ’ ^

2 7 0 ,
'g30)SU-l0coQ)r—I 0 ü) -H
oa•Hîm•HUO
m
•SEh
O(U ^H j â a
(-4 CM m m1-4 o LD C' 50 oO o m en m CMI— I CM 1— 1 Tf 1-4 m
CD U Q U U U m54 0 u k y y u wP C Eh * Eh Eh u■P 50 50 < < < < ut-4 a (X 03 lT> co \ mP 0 en CM iH 1 1 in m m en 50 m CM m C'' 00 roU •H 1 1 1 rH r4 en co 'Ü' co Q CM co en
4-1 r-1 1-4 1-4 CNJ O en O O O m O co O coc U + 1 co LO en en en f-4 co en en rH 00•H (D pe CÜ Pi CO coP rH m u CQ CQ m y m m m CQ pa54 r-4 H Eh M H H P H H H H H4J 0 y y y u u y y y y y yCO u z z z z z z z z z z z
U-(0u
1
Ë•Hc(13enO
CDenCD (den co(T3 Pco (dp coIdco T)
co CDV d 4-1c 4J 54 CD •H•H (d 0 4-> b54 44 eu •H eu
rHX X c eu p54 54 (cs rH co0 0 (D p ceu eu i-a co p
CTi (N CN LD kOco in
IDin
CM iH tM CN
0)tn
§wsil
co cofC3 +Ji sg -HO T3 t) CD 3 54 Q)
CDen(C3en
sco
V444
&co
iss
(V54PLI—IPUiü
i13 CD 3 r4 CD "-4 CO O eu U
CO co fd P(d CD p •H 0[Q P eu M 540 m u •H -H CDP en fd co -P 54 4.)co CO -H •H •H CD t u fd CD COc (d en M ü 54 -H 0 13 N 00 p 3 fd fd 0 en 4J •H +J •U•H 0 54 ü tau 3 fd M 4.) 3 CO4-) 5 CD M CD M ,54 £3 3 •U CDu 0 id fd CJ C54 <44 eu CO 4.)

271
(Dg•HC
m
•§EH
440) -P U (d k r45 o o enw H
(Dup-p1-4pu'c•H<d 1-4 p f-4 4J Oœ u
U4O en P54 -H(D (d b 54 E -P p en Z
I•HCpCnpO
X XP P P P PO O 0 O Oeu eu eu eu eu
œ u oen CM CM mH o O OQ en en en k Z Z Z * * * *
I—I f-4 CM
P•Hb(U4Jfd•H enO P0 •Hen fden pP +JenenA P fd CDÛ4 (D eu eu en uen (D P en en 0 pen en P CDen p eu en en •H Pfd iH P fd fd tn CDP •H 0 P P 3 440 3 Q P CDeu M e CD P0 en 44 0 0 fd13 13 13 P3 V. 3 3 CD(D 0 CD CD ben 0 en en 4->eu 44 eu eu O
U y
s4J
poeu
t" oTf in O CM rH 00 CQ CMCQ PQ rH H H CM Z 4e
iH rH iH
en3, ü en "HP 4J<u (ü en •H ü P ü fd Q)« 8'3 iï '3 -2■U Ü 0) 3 3P, H (n Q) -Hen -u rH fd ü P fd ene o Pp 44 Q I
g< f<

2 7 2 .
where S and C are the similarity between OTU's j and K in jK jKthe similarity matrix and in the phenogram respectively (Sneath
and Sokal, 1973; Sneath et al., 1981).
The cophenetic correlation coefficient, r = 0.8899, (Sneath,
1978) demonstrated that the results given in Fig. 7 3 did not
differ markedly from the data contained in the similarity matrix.
The mean inter- and intra-group similarities with associated values
for the variance and standard deviation (Table 46) provided further
evidence of the validity of the separation of the organisms into
clusters A-F and groups I-III.
The Ovclust program of Sneath (1979) demonstrated, however,
that although clusters A, B and F were distinct entities, C, D
and E were not (p " 10%). Indeed the considerable overlap
between clusters C, D and E suggests that they ought to be
considered as one cluster (Table 47). Groups II and III were
separated significantly from all the other clusters but group
I could not be distinguished readily from clusters A-E (Fig.
73). These findings are supported by other results (vide infra).
Distinctness of clusters/groups and their relationships in hyperspace
The overlap statistics of phenons ( clusters of groups ) were
calculated by a program which tests for the significance of over
lap of 2 clusters in Euclidean space. This program used the number
of operational taxonomic units (OTU's) in each sub-cluster/group
and the inter- and intra-centroid distances. d based on the

273.
VO CO 003 88 :
in Q VO O Q COCOO O po O O
2 8S S 8 S
CJl rH in min Q in Tf Q m ■vrQ M" m4CO o O vo O O r~ O O TT
CO CM O CM 0\ O M- OCO Q VO O CO CM Ô VO CM QCO O o O O O O r- O O
TT -4 CO (Tv CM o -H CM ro Or- Q tM V Q in o ifi ro g VO r~ O O r- O O vo 0 O VO O O
CO rH CM CO rH TT ro roCM rH r~COO g ro rH Q COg ro (Tv g m CO g ro o g COCO O o COO O CO O o o o VO 0 O vOo o in 0 O
(Tv Ç O rH «f CO c5 (TV i-H (TV rH Tf O «T 00m c (Tv O CM vo I o (TV g CM m g VO o O -qr r--(TV c5 O r- O O r- c5 O o o vo o O vo o O voO O m 8S
(Tv VO rH CO in rH CO ro rH r-1-4 in CMCM rH Q CM CO g CM m g CM O m g r- o ino CO 5 o r- O O r- 0 O o o voO O VOo OTf
88
HO
r—1 EÊ 10 >H (1)to ^ 'O0 O nsM C T3Di (TJ C•H toc u u

274
Table 47. The distinctness of clusters as measured by their
overlap
Cluster/Group
*
A — >99** >99 >90 >99 >99 <90 >99 >99
B - - >99 >99 >99 >99 <90 >99 >99
C - - - < 90 < 90 < 90 <90 >99 >95
D - - - - >99 >99 < 90 >99 >99
E - - - - - >95 < 90 >99 >99
F - - - - - - >95 >99 >99I - - - - - - - > 90 >90II - - - - - - - - >99
III - - - - - - - - -
A B C D E F I II II]
* Ovclust output estimated with the V(O) = 0.025 overlap setting
(Sneath, 1977, 1979)
** Probability that overlap is less than 2.5%

275
relationship, d = (1 - ^ to calculate an index of overlap
(V^). The level of overlap expected for a rectangular distribution
(Sneath, 1977) was used as a critical level of overlap to compare
all pairs of subgroups/clusters.
Thus from Table 47 the distinctness of each cluster/group
from every other cluster/group shown on the dendrogram (Fig. 73)
is measured by the "w" statistic (Sneath, 1977; 1979; 1980).
Cluster A (16 OTU's; 85% S). This group included 11 isolates
from sausage and 2 from ingredients. More than 80% of cluster
A strains used 48 out of 126 of the carbon sources tested (Table
48), and all produced fluorescent pigments. Organisms of this
cluster were identified with Pseudomonas fluorescens biotype A
(Stanier et al., 1966).
Cluster B (8 OTU's; 92% S). This group contained 7 isolates
from sausage and 1 from an ingredient. More than 8C% of these
closely-related isolates used sixty out of the 126 carbon sources
tested (Table 48) and 63% produced fluorescent pigments and
lipases although one strain only gave a typical egg-yolk reaction.
Organisms of this cluster were identified with Pseudomonas
fluorescens biotype C, Stanier et al. (1966).
Cluster C (88 OTU's; 85% S). This group contained 70 isolates
from sausage and 17 from ingredients. All the organisms of this
major cluster (54% of pseudomonad isolates), which contained
Pseudomonas fragi NCIB 8542 also, were non-fluorescent and did
not produce extracellular lipases or proteases. More than 80%

276.
E0P44MEm•HCroenU04-10W•HW>11—4rOCrOf—1(0U•H54Q)EDCC•Hb0)W3014-»01014-)04-1010)4JrorH001•HQ*D -P0 rO54 0)en E\54 b(D 014-) 0)01 013 01rH Uü 0
p44 a0 bC G0 (0•H4J bü 01rO 0101 pCd 44
00
01rHbroEh
M
Q
U
m
50f—I
50
'f
00CO
00
LD m rH (Tl o r- CM m 50
o o o o
8 °
8 ° 8
O O m m OO CN f'-
LO m o O O r' CM o
O O O Q O 50 ^ O
g o o 8 o
rp- PO CTi rH oTf LD (T)
00 CM co CM o00 rH (O r4
C'- PO t'- PO O 00 rH Tf UO
u §G •Hd 01 0 m -PG G b -p ü0 "d E 0) G roP fO P 4-) 01 01O p 0 b G s P> 4-1 44 4-> b 01 tT»P CO 0 d E •H E01 >1 0 G en Û4 rO•p 44 G Ë 0 •H G P01 0 0 co k k D CJG rHrH 0 0u Z u
8 °
o o
8 °
8 °
+J
iI:fdr—I « S'rHk
0)IU•HU4-5•ri•PfH3S
in O OCM
o o
O O O
o o o
o o o
o o o
o o o
00 o o 00
o o r"O
O O O 0o o o
8 8 8 ° i—l i-H iH
8 8 8 ° f—I rH rH
8 8 °
8 8 8 ° rH I— I rH
8 8 8 °
8 8 8 ° rH I—Il—I
8 8 8 °
8 8 8 ° rH r— 1 I— I
4-)G01 01ü G 001 •H P01 N m 4JP rO \ rO0 GG 0) G brH b •H -Pk k k Sô
o o o o2 3
S°88
8 ° 8 °
O O O Oo
o o o o o o
8 ° 8 °
8 ° 8 °
o o o o
_ o 0 ^ 00 in o rHTf rH PO
01010üGrHCJ1 UQ •p 01u b 01e •H 0 rO0 b p t-Hp 0 01 rO44 P to 400) G rO2 < <C Uo<
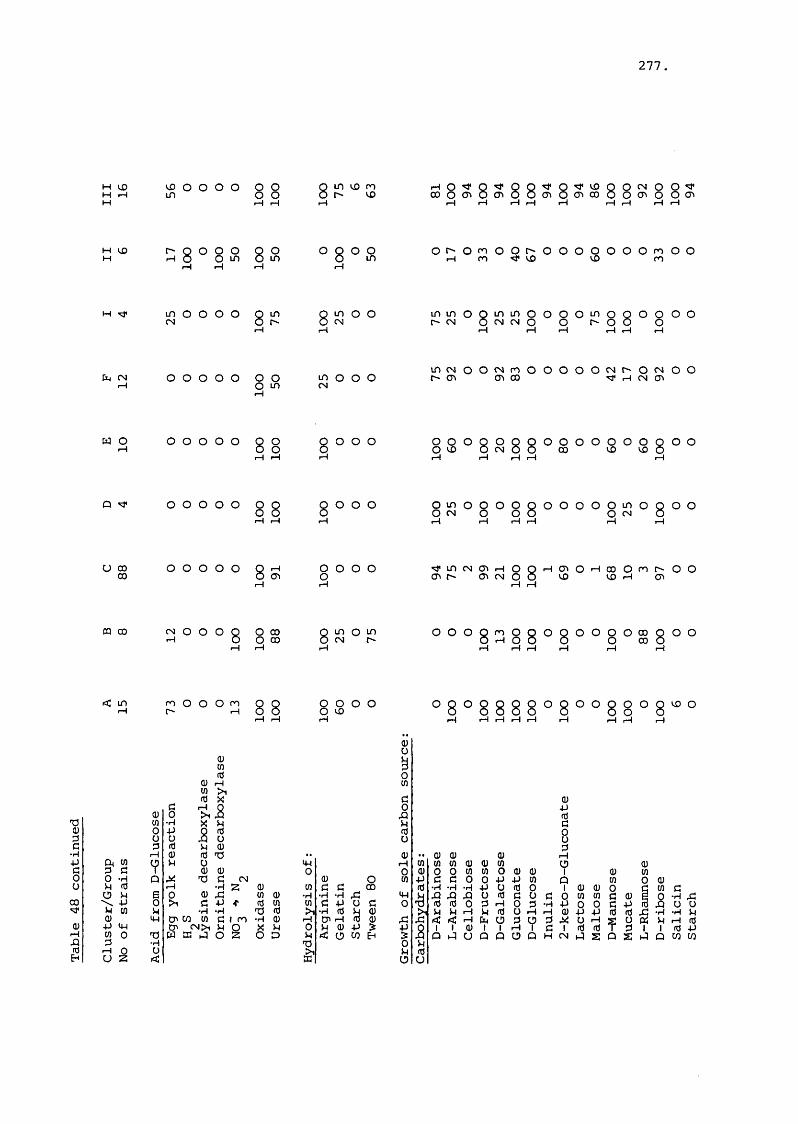
277
H 50 50 O O O O o o L D o o m 50 ro r- 50 r P O ' ^ O ' ^ O O ^ Q ^ 5 0 O O C N O O ' : 3 'o o o o i o m o o o i o o i o o o o o i o o m
r—I I— I
H 50 M r Q o o o Q orH o o in o 50 ° 8 ° S O r ' - O r o O O r - ' O O O O O O O r o O O r H f O S f 5 0 5 0 ro
50 o o o o O 50 CM o r- lO O OCM
l O i n O Q i O i O O O O O i o O Q O O O O [ ' - C M O ( N CM O o C ^ o o O
r H t - 4 r H r H r H i H
k CM O O O O O o oo 50 5 0 o o oCM5 O C M O O C M C 0 O O O O O C M t ' ' O C M O O r- m (71 co TT rH CM
w o o o o o o g o C O Q O O O Q O Q O O O O O O O O O O O O 50 o CM o o 00 50 50 o
Q o o o o o o8 8 8 O O O 0 L 0 0 0 0 0 0 0 0 0 0 0 5 n 0 0 0 0 o CM o o o o CM o
u 00 00 o o o o o <71 G O G OG
' v d ' i o c M C T i r H G O ' - i o i G r H o o G c o r ' - G G C T i r ^ (71 CM O G 5 0 5 0 r H (71f—I 1-4
DQ 0 0 CM o G G G G 00r H G G 0 0
8 50 o 50 ( M t ' -
G G G g c o O g G g G G O G O O g G G
c o G G G f O C ' r H 8 8 G G G G
G 5 0 ° 8 8 8 8 ° 8 ° ° 8 8 ° 8 5 0 G
(U(fl(TJ( U r 4w >1G XG «H Qd) 0 >1 bb (0 •H X p0 0 -P Q (dG u U b uG G (d P (U•H rH 0) (d 43-P k co G p o 44G G G T (D 01 00 0 •H Q b T3 G CM 0)U P (d iH •H Z (U CO G GG p e 0 (U b (0 Q) •H •H •H00 > -p 0 îP G +j f td (0 CO G b'd* P co p •H •H 43 (d >1 •H (dQ) 44 Di co co G 1 co •H 01 iH en rH01 -P (H Di CM>i P G X p 0 p 01rH CO 0 b W m b G Z G D p < Gb G •H 43G r-H 0 ü >1Eh U 2 < K
13-P
g§
I
en en
w (Q(13 K3 p p
k G
w co

278
H O Q OH O O OM r— I I— I I— I
O O O ' ^ r m œ o o c n O oi 01 m O rH cooO'^f'^roo'd'oooocoiOTrcorooioiœoicocDCDr'-oi ^ n CM o 00 01 Q 01 o 00
H o O O c o O O O O O O O OPI LD IT) O O O O O r - O O O O c n m vo 00 O O O O en ^ in PI
8 8 8 O O O O c o i n m m i nm o co , CM CM CM o o i o o m o m m o o o
CM o O' r'- C" mO O 50 o 50
C'- 50 CM
k 00 O O r '-OOojOOOLDOO vo 01 50 CM O O C M P i O O O O O O o c M01 00 oi o o o 00 o
w o o oO 00 O O O Q O O O O OO O Oi CMI—I I—I
O O O Q O O O O O O OO O o o o o o oo o 50
Q lO 50r~ r- 8 8°°8°S°gS o o g g o o o o o o o o o o o 50o o t"rH r—H
u PI r-4 r~ O I O O O O O O O O O '00 00 01 01 o o 00 00 O O O r H M O O O O O O^J'OOC'o o o CM o o vor H I— I r H i H i H
CQ o o o O O O O O O O O O O o o o 00 8°88°8°S888r H r H r H
° 8 8 8 SI—I '—I I—I
iH rH OP I c~ 8 ° ° 8 8 s ° s : °
I— I iH rH rH rH O vo PI o o 50 01 o o CM
43<Dg•Hg0U00
gEh
§■OPGInObwGrHu
01(fi 01 W 0101 0 Ü) 43 01 bm rH 0 •H b G0 td rH ü G pp b >1 (d b >1ü 01 X 01 bG p 1 >1 u Gw Eh Q bbGk
< PQ
S(d
GPQ
IOH
td >1 P P A A k (d fd 0) u U K
(Dbtd Q)S ^ o td•H p C5, 01 o rH
01 b td p 01 I—Itd>ÔH
W43•H
gü•HrHr
SO•HQ
01btdo
•H43 01 01 (fi b01 01 b 01 01 01 01 b 43 GG b G 01 b 01 b b b G •H rHG G P b G b G G G G O fH(fi P G G G G rH U p •H G 00 G b 01 0 1—1 01 G 01 U >1 uü G rH rH G E b b U X >1•H iH G G X •H 01 3 G 0 rHW k G 2 2 O k CO co CO p G43
>1
01gPIP
ü >1 b b >1 K u tdI td

279
rH o CO I'> LD <T> PI O 'd'O cn 03 O iH O
O O 'd' o O m oI— I I— I
8 8 8S 'd' 'd' rH Tf Tf Tf03 03 C7( <N <7i 03 (Ti
O O O O 00 O O O O P3 o p) in o o o o o o o o o O P3 o o o o oP3
O O O vn LD in LD LO o O 50 Q inCN O CM o O'I—I I—I
O O i n o o i n i n o o t'- O in CN CM o i n i n o O O O OCN p' m
k CO O O O'00 rH O O rH C'- O CO <73 rH O CO OOOOrHr'-r-com in 03 vo VD CM O O O O O in O
w o o o oCM vo q o o o Q orH o 03 o O o o o o o o o o orH rH o rH ^ rH o o o o o o o
O O m O O in O O O O O Oo o o o o o o m o o o m oCM O in CM O O O O O 0 o
o in vo CN 00 P) vo vo O CO vo COC73 00 03 O c M O O O m m i n oO rH m O O 00 O C" rH
rH m rH
CQ o o o O m O ro o o CM O rH O O ° ° 8 8 8 8 ° ° rH rH I— I rH
O O O -H o CM CM
43GG G G GG b b b•H G G Gb k P P PG G b b b0 0 G P p pO P b G G GG G Eh Eh Eh00 \ rH 1P G 1 T 0G s toG b 1 1 1 GrH to b Q b Bb GG rHEh u
S ° 8S 8 8rH rH rH
G bb GG G G rHb G b 03 GG 0 G G bb U b G 0 G•H G G 0 b >G P P 0 G G0 b b G X Pü •H •H b 1 >1< G U H 0 k
(/)rH0&engG
I
12
O 03 O vo o O in rH m o m O vo m vo m o m o H o<N P) CO
>1 rHrH 1—i 003 p rH uG Ü 0 >1G >1 b rH.G rH rH •H enfH O' 0 to GrH >1 G b r-H 0 rH G 1—1q b G •H 0 G 0 G 0b G G p p H b 1—1 b•H CQ rH b G 1 •H •HG >1 b U 0 G a b0 P3 b >i >t to G 0 p'9 b p rH G G p 0< CM w w O B 2 k w
I—I
B pG k
B rHG 0
G G b I&PCQ b p b o kI b CD G CO IG W G 2 -H G
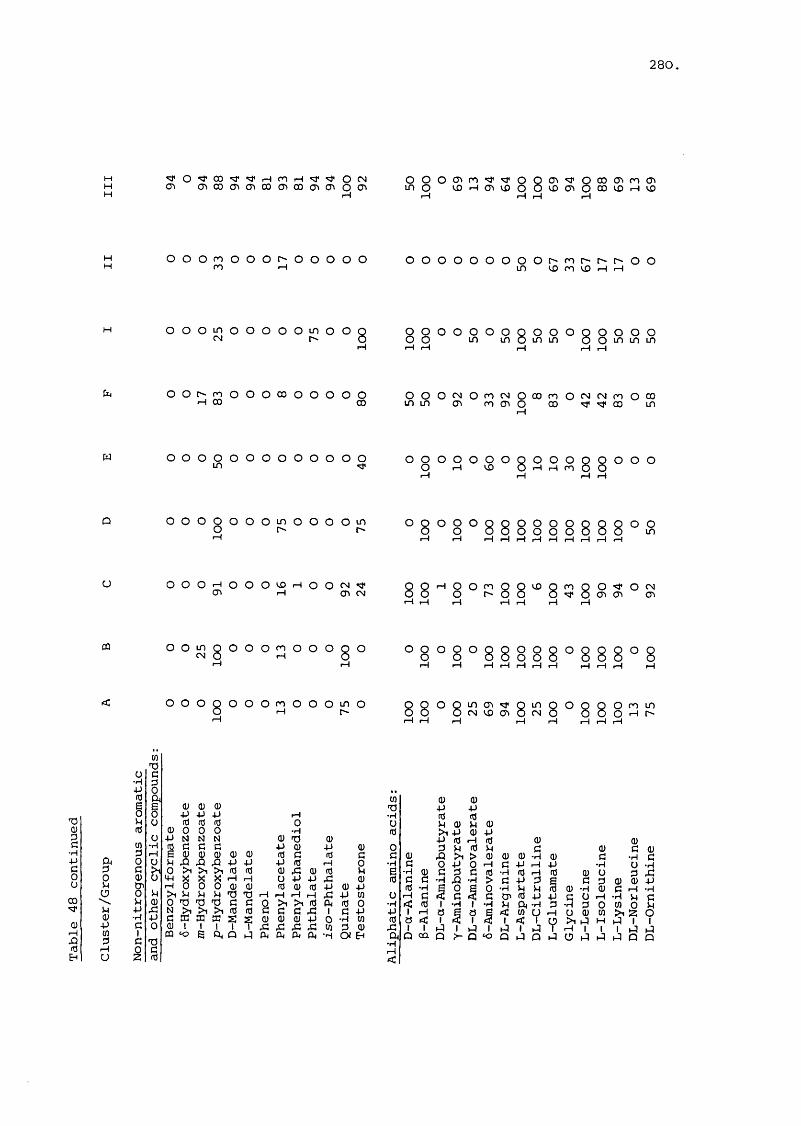
280.
^ O * d * 0 0 ' ! 3 ' ^ r H P ) i H ' d ‘ ' d ' O C N 03 03C003030003000303003 O O O O 3 m < f ' ; f O O O 3 T f Q 0 0 O 3 m O 3 m o mrH03voo05D030oovOrHm
O O Q m O Q O P ' O G O O G m iH O O O O O O O O O P ' m p ' p ' p ' G O m m P 3 v o i H i H
G G G L O G O G G O m G OCM p- Q O O O O O G Q O O O Q O O O O G G m m O m m o O m m m
k G O P ' P i G O G C O G G O O O I—I 00 co O G O C N G m c M G o o r o G C N P H r o G C O m m 03 P3 03 G 00 oo m
H O G O O G O G G O G G O Gm O Q O O G O O Q O O G Q O G O O G rH vo G rH rH m G O
O G O O G G G m G O O O m G P' p~ °8°8°888888888°SU O O O r H G O G V O r H G G C N ^ 03 rH 03 CM Q Q r H G G m O Q v O O r o O O ' d ' G C M G O G P ' G O G 'd' G 03 03 03rH r-H r—1
CQ O O ^ Q O O G P O O G O Q O ° 8 ° 8 ° 8 8 8 8 8 ° 8 8 8 ° 8 rH rH rH rH rH r-H rH r—I rH ,—| i—|
O G G O O G r o G O G m GI—I p- Q Q O Q m 0 3 - v } < Q m G O O O O G O G CM vo 03 G (N G G O GrH rH rH rH i—I i-Hr-HrHr o mrH P-
43I•HbCOuco
bGEh
§-2U
*bwGrHU
2G§PGMG§CDS'g•HGICg
W43GI0U•prH
p10
1
01G(D 0 b N
I IrH Sg’-O0 1CQ <o
Q) bSN G 0 0b bS S "p p "O43 43 G
G Ô
0 •H 0 13
>1 >1 G G G b 0 0 b b b b
0bGrH5bgIOV)C4 C4 k C4 -H G< Eh
W43•HüGB•H§U•Hbsa•H
BG G G *H 'd 53 <C G I rH? “FQ cû
0bGP 0 tP b b G G P
0bGP 0 0 bbS• " g
i lQ ‘O
tn G P euG P b •Hw u
0 G -P 0 ü
U 0 M :P Z îP b H I I
II0 0 bS 3W o
H
. . IL b bG b b b Q Q
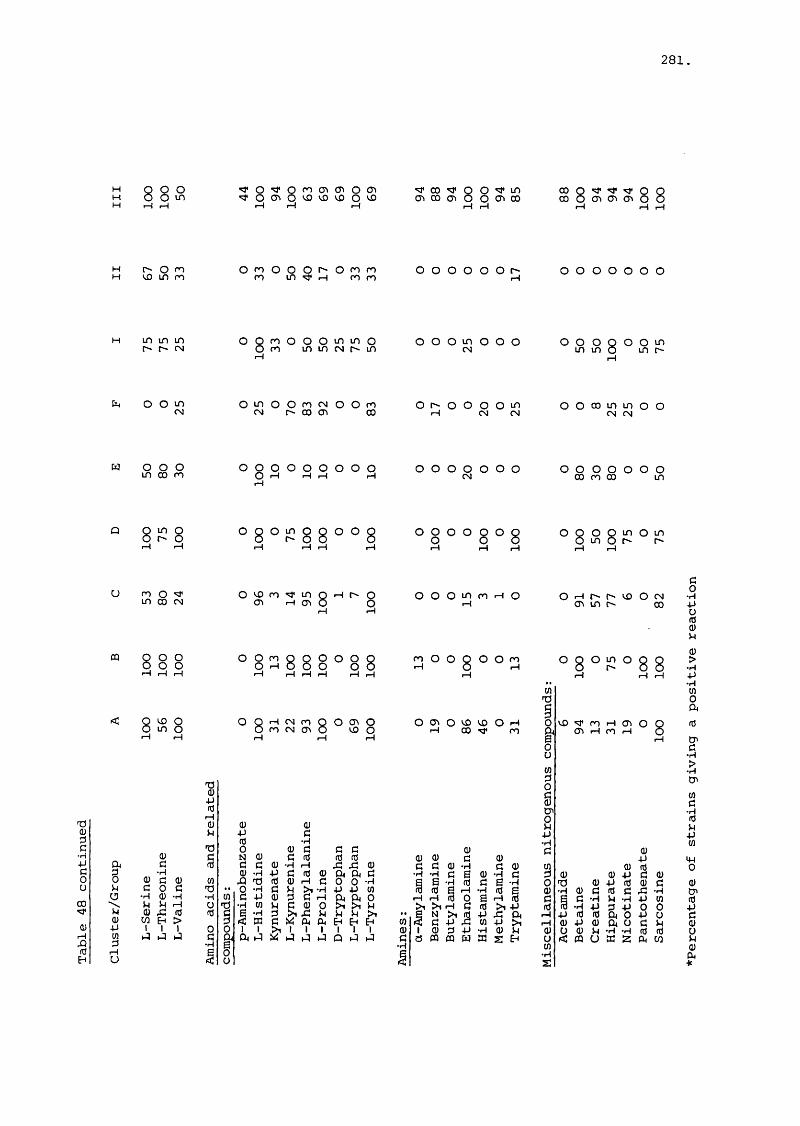
2 8 1 .
8 8 gT— I r H
'3*O'^O<n03a3OC33'd'OCTiOvOVDVOOvOI—I I—I I—I
'd' CO ^ O O ^ m03 co 03 o o 03 co co Q '=J* 'd' ^ O co o 03 03 03 orH iH
r - O m vû m P3 O co O O O t" ro lD ^ iH O co ro ro ro O O O O O O t" O O O O O O O
LD LD LD p- p- CN O O r o O O O m i o O O ro m m CN p- m O O O m O O OCM O O O
in uoO O in
m p-
k O O m CN O m O O r o c N O O r o CN p- CD <73 co
O t" O O O O m iH CN CN o o co in uo o oCN CN
w o o oLO CD co o o o o o o o o oO rH rH I—I rH O O O O O O O CN o o o o o o oco ro CD LO
8 : 8 r-H rH ° 8 ° : 8 8 ° ° 8 O O O O O Os s .48 ° 8 g 8 G lO p-
U
OQ
ro GLO CD CN
8
8
8 8
LD G lO G
430GG•H 0b CD GG G •P0 0 0 GU P G 0CJ •H 0co \ P PP 0 b0 CD Eh0 b 1 1rH en b bb GG rHEh u
GLDro^ioO'bP'G 03 rH C73 G G
O O r o Q Q O G O O rH G O O G
O O r H ( N r o 0 0 0 3 G O ro CN C73 O LO G
0b§N 0
b 43B b
s G GG G G-H b b 0G 0 CD (k G ■ ^ 0 0b bG< m CD b
G 0g gg 2 ........X b b b Q b b
U U
O O O LO ro rH GrH
co G G O O G rorH G rH
G C33 G LO VD G «H I— I co ^ co
G 0 -Hb | §G S rH>1 rH Bg g ' . s0 G bPQ CQ k S3
0G 0HrH f3
S Eh
G rH P~ p. LO G CN 03 LO p- 00
LO G p- 8 8
en431uenG0Sen2 b •H G
1 i
LO «d* ro rH C73 G(73 rH ro r4
0b0 G
0 0 b G 043 0 b G 0 G•P 0 G G G b •P
G •P P •P b en•H b G b 0 0
b G G CD 0 b u0 b 0 CD ü G pU 0 P •P •p G G< PQ U X 2 PD CD
Go•HbüG0U
>•Hb•Hen0CD
I.503enGuben
MHo0
fB0(D
*

282
of the strains utilized 45 of the 126 carbon substrates tested
(Table 48). Organisms of this cluster were identified with Pseuo-
monas fragi.
Cluster D (4 OTU's; 86.5% S) contained 4 isolates from sausage
and was related closely (83.5% S ) to cluster C. The isolates of
this cluster did not produce fluorescent pigments, proteases or
lipases. Forty-five out of the 126 carbon sources tested were used
by at least 80% of strains (Table 48). Isolates of cluster D,
which were separated from those of cluster C on the pattern of
substrate utilization (Fig. 74) were identified with Pseudomonas
fragi .
Cluster E (10 OTU's; 83% S) comprised 10 isolates from
sausage. Organisms of this cluster used 28 out of the 126
substrates tested (Table 48) and were identified with Pseudomonas
fragi. All the strains in this cluster failed to grow on quinate,/benzylamine, histamine, tryptamine and D-xylose for example
(Fig. 74).
Cluster F (12 OTU's; 82.5%S) included 11 isolates from sausages
and comprised Gram-negative, non-motile bacilli capable of utilizing
23 out of the 126 carbon sources tested (Table 48). Members of
this cluster, which were similar to the group A isolates from meat
(Davidson et al., 1973), were identified with Moraxella.
Group I (4 OTU's; 73% S) comprised 3 isolates from sausage
and 1 from an ingredient. These strains, which used 23 out of the

283
126 substrates tested (Table 48) have not been assigned to a species
The pattern of substrate utilization suggests that 3 OTU's resemble
Pseudomonas fluorescens whereas one resembles Pseudomonas fragi,
(Table 48).
Group II (6 OTU’s; 77% S) included 4 isolates from sausage.
These strains produced pink/brown pigments on the medium A of
King et al, (1954), H^S and DNAase. Few carbon sources were
utilized (Table 48). Members of this group were identified with
Alteromonas. They did not have an obligate requirement for salt
(their tolerance of salt was not tested).
Group III (16 OTU's; 00% S) comprised 16 isolates from
sausage. These strains used 98 out of the 126 substrates tested
(more than 80% of strains, positive) and resembled Pseudomonas
fluorescens biotype G of Stanier et al. (1966), who stated
"Biotype G is frankly a provisional group, which lacks the uni
formity characteristic of the other six biotypes". It is note
worthy that 3 of Stanier's strains (1, 99, 124), which clustered
together,in the numerical analysis of the original data (Sneath
et ai., 1981), shared many properties with organisms of group III
of this study.
Discussion
The results presented above have confirmed the observations of
several workers (e.g. Davidson et ai.,1973; Shaw and Latty, 1982;
Molin and Ternstrom, 1982) that the majority of pseudomonads isolated

284
from meat differed from those studied by Stanier at al. (1956).
Indeed only ca. 19% of our strains of Pseudomonas from British
fresh sausage and ingredients could be identified with the species
described by Stanier et al. (1966).
Despite some claims that fluorescent pseudomonads predominate
on chilled meat (e.g. Kirsch et al., 1952; Wolin et al., 1957,
and Ayres, 1960), this report supports the findings of other workers
(e.g. Davidson at ai., 1973; Shaw and Latty, 1982, and Molin and
TernstrOm, 1982) that non-fluorescent types dominate. This study
indicated also why the over-simplistic scheme of Shewan at ai.
(1960 a) has been used extensively by food microbiologists and
has reinforced the findings of others (e.g. Gardner, 1965; Shewan,
1974; Barnes, 1976; Law at ai., 1979; Gillespie, 1981) that the
dominant pseudomonads of chilled proteinaceous foods form a
distinct group (Group II of Shewan at al., 1960 a). The pseudo
monads included in this study were not isolated from"blooms"
and therefore the conclusion that Pseudomonas fragi is the dominant
Organism both of fresh, spoiled and processed meats appears to be
justified (e.g. Blickstad at ai., 1981; Shaw and Latty, 1982;
Molin and Ternstrom, 1982; Martin and Patterson, 1982).
In the past, unsatisfactory media have been used for the
isolation and enumeration of Pseudomonas from meat and meat products
in which these organisms were not numerically dominant. The
contention of Gardner (1980) that CFG agar (Mead and Adams, 1977)
is a useful medium for the recovery of Pseudomonas is supported
by the results presented above. Although Pseudomonas spp. are

285
considered to be minor contaminants of sausage (Dowdell and Board,
1971), and are not implicated in the spoilage of the product,
the biochemical changes brought about by these metabolically
versatile organisms may be out of all proportion to the size of
their climax populations in sausage and their potential to
contribute to spoilage of the product should be considered in
future studies.
The presence of sulphite did not appear to exert any selective
pressure on the species of Pseudomonas which were recovered from
meat and sausage. Indeed isolates from sulphited and unsulphited
samples were represented in all the major clusters or groups shown
in Fig. 73
Addendum
Numerical taxonomy of Pseudomonas spp. isolated in previous studies
of sausages and meat
The data upon which Davidson,Dowdell and Board (1973) based
their discussions of the properties of Gram-negative aerobes
isolated from meats, including British fresh sausage, was available
for numerical analysis. The Clustan package of Wishart (1978)
was used with the objectives of (1) establishing whether or not
Pseudomonas fragi was a common isolate in their work and (2)
assessing whether or not the Gram-negative aerobic flora of
sausage had changed over a 15 year period.
Of the 231 polar-flagellate Gram-negative rods studied by

286.
Davidson et ai. (1973), the properties of 218 isolates were available
for study, 104 strains had been isolated from beef (Davidson, 1970)
and 114 from British fresh sausage.
The majority (206 strains) of isolates were contained in 6
major clusters which fused at 65.8%S (Fig. 75). Twelve unclustered
strains and those of clusters 1, 2 and 3 (161 OTU's) were closely
related (82.3%S). There was little relationship between clusters
4 (20 OTU's), 5 (4 OTU's) and 6 (21 OTU's), however, with fusion at 69.65%S
and 65.8% S respectively (Fig. 75). A separate analysis of
the 104 beef strains assigned 93 to 8 clusters. Although the
majority (55 OTU's) were contained in cluster 1, the extent of
overlap between the 78 OTU's in clusters 1 - 5 , which were separated
from the remaining 15 OTU's of clusters 6 - 8 at 72.5%S (Fig. 76) ,
suggests that they comprise one group only.
One hundred and four of the 114 strains from British fresh
sausage were contained in 5 clusters (Fig. 77); the majority (62
OTU's) of strains being in cluster 1 (87.2%5). This cluster was
closely related to cluster 2 (84 S) and 3 (78.4 S). Cluster 4
and 5 (15 OTU's) formed minor fractions of the isolates and
were not closely related to the cluster 1 - 3 complex, linking
at 75.9%S and 62.6%S respectively (Fig. 77).
The conclusion that the majority (79%) of the 218 strains
depicted in Fig. 75 were Pseudomonas fragi (clusters 1, 2, 3 and 12
unclusteréd strains) is supported by a detailed analysis of utili
zation of specific substrates and other features (data not presented).
It was notable that these organisms were easily distinguished from

287
the fluorescent ones contained in clusters 4 - 6 (Elg. 75). The
proportion (75%) of isolates from beef which were identified with
Pseudomonas fragi was of the same order as that (83%) obtained with
isolates from British fresh sausage (Fig. 77) and in the study (73%)
described on p . 258 - 285 . Thus it was concluded that changes in
composition due to manufacturing methods had not influenced
appreciably the pseudomonad flora of sausages over a 15 year period.

LOL O0 3
G- Hu0buGb
0>• HbGo>0GI
Uo
u•rHbo%G
>.rH0g03 • H I—IbOH
.gb0
■gGto
'B
1scoa4H0
U030?043430• H4H•rHfH1CD
0
to
SM.Xëabto0vti
Gtob
0•rH4305hB’• H
0B'0S0
U4-1
■S
giHB•rH
gD
g ’
aG
• H5h0btoG
bG0
•rHu
• H44440OU
CD
0bb
b
S
1-4GU
i-HGü
>1b•rH54G
a• H
• §CO
0I0B0k
&GM
0%
Bêk
L OL O03
MG• U05h
0•rH
bCD
ab0•rHb
to
COë3
to
G
•§3£<
to aG 0 G0 G
•H 0 bU u 0G 0
0 U0 «4 05h 0 -Hb 3 G3 a Ga 44 b
CDto 0
43 G G0 G G 0G 0 Qa e E 0
•rH 0 O ato 5h 43 R0 0 3 bG •u 0 oG a 0 •HD k b
HH H
H H H
U0ab0•rHb
toSto
ë3
to
I0k
CQ
aGAG
G
■ §
0k
U
OinCMco
PQHCJz03 mÜ 03 m
a co rob co 03 000 (33 oü CQ m QG H CQ (N ap y H U O ro 00Ü z U E4 a o 00a Z b Z a aG a z zCJ G 0 CQ
p 0 G a G Hk, 54 G 0 0 0 CJ0 0 0 0 0 zb b a G 0 GCJ 0 a a G a G2 0 a a G a 43b b G 3 0 3 a0 0 CJ 54 E 54 bb 0 a 0 0 0 30 b G G 43 G aG 3
a 0CJ 0 0 0 0 0 0< k k k k k k00 (33 O r4 CN rof“H rH fH a 1—1
r HOOCNQk+a00Oa
m mCQ a OH CN 0 CN roy 00 a a U 00z G CN a
CQ 0 CQ O OG H 3 H a r Ha y G u z
a a CJ z 0 z DQa a G HG G a G a CJ54 54 0 G X a 0 Z44 44 ü 0 0 a
0 a a0 0 0 G 0 b 0 54G G G a G a G 0G G G a G 0 G N
0 3 b 0 bE M a E 30 0 0 0 0 0 b43 43 43 G 43 43 03 3 3 3 30 0 0 0 00 0 0 0 0 0 0 0k k k k k k k k
Q H a CN r o i n L O
r oC"
0UGo>•rHk
0bsbO
k

289
50 60% Similarity70 80 90 100
< 15-18 >
<19-24 >

291
Figure 75 Simplified dendrogam of 218 strains of Pseudomonas spp. isolated from fresh beef and fresh pork sausage by Davidson et al. (1973)
Footnote; Percentage similarity calculated with Sg^ coefficient; clustering by UPGMA
^Pseudomonas fragi
—Fluorescent pseudomonads
5
100 90 80 70 60Vo Similarity

292
100 90 80% Similarity
70Figure 76. Simplified dendrogram of 104 strains of Pseudomonas
spp. isolated from fresh beef by Davidson et ai. (1973)
Footnote : Percentage similarity calculated with S^^ coefficient,
clustering by UPGMA.
Clusters 1 - 8 unassigned.

293
Figure 77. Simplified dendrogram of 114 strains of Pseudomonas
spp. isolated from fresh pork sausage by Davidson et al (1973).
i100 90 80
% Similarity70
Footnote : Percentage similarity calculated with coefficient;
clustering by UPGMA.
Clusters 1 - 5 unassigned.

294
DISCUSSION
When this project began there was circumstantial evidence
(Brown, 1977; Abbiss, 1978) that sausages made on a small scale
in a test kitchen or laboratory rarely raimiced the commercial product
especially with respect to the size and composition of the micro
flora. As a consequence the majority of the present study was done
with material taken at the point of bowl-chopping in the factory
of a large sausage manufacturer. Thereafter the material was
treated in such a manner that it simulated commercial production.
During studies of changes in the growth and composition of the
microflora, particular attention was given to the previously
ignored Gram-negative organisms. Pseudomonas spp. and Entero-
bacteriaceae. The development of an analytical technique that
distinguished between bound and free sulphite was an essential
stage in this study also. Indeed the technique proved to be
particularly valuable in studies of sulphite-inhibition of
Salmonella.
Analysis of the ingredients of fresh sausage showed that all
the major members of the microbial association were present, the
size of the populations being influenced by the ingredient and
methods of preparation. Thus mincing of ingredients and then
storage at chill temperatures was associated with a moderate
increase in the numbers of micro-organisms. This was attributed
in part to the re-distribution of micro-organisms on the meat
but mainly to the growth of psychrotrophs. As mincing of meat
releases enzymes which cause a rise in the concentration of glucose
(Newbold and Scopes, 1971), microbial growth was probably aided

295 .
also by a fresh supply of nutrients. Indeed Ayres (1955) contended
that comminuted meat can support a 10-fold greater population than
jointed meat because of the increased surface area and an enhanced
supply of nutrients. Backfat harboured fewer organisms than minced
meat. This could be due to the low nutrient status of such
material as well as the limited contamination to which fat is ex
posed because of the protection afforded by the skin until butchering.
Oxygen-catalysed toxic substances e g. hydroxyls (OH ), singlet1 “ oxygen ( Og), peroxides (O^ ) and hydrogen peroxide (H2O2) also
be involved in restricting microbial growth on fat. The rinds
invariably contained large populations of micro-organisms. As this
was a cooked ingredient, post-cooking contamination before, during
and after mincing,together with opportunity for microbial growth
all contributed to the large populations. Thus it was the practice
of the factory to cook the rinds and then allow them to cool slowly
at ambient temperature before mincing and storage overnight at
chill temperatures. Further evidence of factory practices increasing
the level of contamination of ingredients came from studies of
sausages made in the course of a working week. The chill storage
of butchered meat over the weekend was shown to influence significantly
the extent of the initial contamination of sausages (Fig.20). Thus
on Mondays and Tuesdays sausage contained > 20 fold more micro
organisms than those produced on Wednesdays, Thursdays or Fridays.
This situation is likely to be due to the growth of micro-organisms
on the meat rather than to build up of contaminants over the weekend
in or on the processing equipment which was thoroughly cleaned at
the end of each shift.
It was notable that the size of the initial microbial population

296,
in fresh sausage was related inversely to that of the final (climax)
population (Fig. 21). This was noted by Dowdell and Board (1971)
and Brown (1977). It is possible that in a sausage having a high
initial load (usually lactobacilli) the ratio of one of the co
dominant groups of micro-organisms (e.g. yeasts or Brochotbrix thermosphacta) to another may influence the capability of the microbial
association as a whole to achieve high climax numbers. Although the
reasons for this have not been elucidated in the present work, it is
possible that preferential utilization of a particular carbon or
energy source or production of a selective growth-stimulating/
inhibiting compound may be involved. Of practical significance,
however, is the fact that the presence of a low initial microbial
load, as compared with a high initial load, enables the microbial
association to develop at a faster rate. Thus caution should be
exercised by the "trade" should they aim for a product having a low
initial contamination. By doing so, the selection of a microbial
association capable of faster growth might accelerate spoilage.
The hypothesis that the contribution of a particular group
of micro-organisms to the microbial association in sausage might
be determined by the concentration of sulphite which inhibited
their growth was tested in broth culture with strains isolated
from sausage (pp. 151 - 157 ). In general the strains of yeasts and
Brochothrix thermosphacta tested were tolerant of concentrations of
free sulphite in excess of those which would occur in fresh sausage.
The lactobacilli and streptococci were inhibited by intermediate
concentrations, especially with incubation at chill temperatures,
of free preservative (Fig. 47) whereas the Gram-negative bacteria,
pseudomonads and Enterobacteriaceae, were prevented from growth by

297
a lower range of sulphite (Fig. 55). Thus the microbial association
can be defined in terms of sulphite-tolerance. There is evidence
(p.168 - 187) that the efficacy of sulphite is enhanced by chill
temperatures. The more rapid binding and oxidative loss of active
sulphite at higher temperatures are factors which are likely to
diminish the effectiveness of the preservative. Indeed the evidence
presented on pp. 160 - 163 (Figs. 64 - 66) and pp. 15F- 157.
(Fig. 58) suggests that the decline in the concentration of free
sulphite in sausage or broth is associated with the subsequent
growth of Pseudomonadaceae and Enterobacteriaceae.
In an exceptional case, a batch of sausage was found to con
tain unusually high concentrations of sulphite. Although the rate
of growth of the yeasts was suppressed slightly, those of Brocho
thrix thermosphacta, pseudomonads and Enterobacteriaceae were
drastically affected (Figs. 39 b, 41 a, b). Thus yeasts dominated
during storage. The substantial growth of lactobacilli and
streptococci may have been associated with an
increased availability of growth factors of yeast origin. It is
notable that Brown (1977) suggested that a'yeast-dominated'
sausage was a factory-specific phenomenon associated with a
concomitant low initial bacterial (i.e. Brochothrix thermosphacta) count. Although he did not speculate upon the reasons, analysis
of his data reveals that the sulphite content of such sausages
was significantly higher than in those dominated by Brochothrix thermosphacta. He also noted that the loss of free sulphite
was faster in the yeast-dominated sausages and attributed this to
increased binding by agents of yeast origin. Although sulphite-
binding compounds were produced by pure cultures of yeast in broth

298
systems (Fig. 59), the evidence obtained in the present study did
not link unequivocally the growth of yeasts in sausages with free
sulphite depletion.
As the results presented above suggested that the Gram-negative
portion of the microbial association was the most sensitive to
sulphite, the composition of their populations in unsulphited and
sulphited sausage was assessed with the view to detecting whether
or not subtle selection of the organisms among the initial contamin
ants occurred during storage.
There was no such selection in the Pseudomonadaceae, pp. 258-285.
Isolates from the ingredients, sulphited or unsulphited sausage
were identified with the major clusters shown in Fig. 73.
Analysis of the composition of the populations of Enterobacteriaceae,
on the other hand, highlighted differences between the species
dominant in preserved or unpreserved sausage (Table 22). Thus
Escherichia coli, Enterobacter cloacae, Serratia liquefaciens and
Yersinia spp. were isolated frequently from the former, whereas
Enterobacter agglomerans and a biotype of Hafnia alvei from the
latter. The hypothesis that this selection was due to sulphite-
sensitivity was tested in batch broth culture. Enterobacter agglomerans (minimum inhibitory concentration, MIC = 50 - 100 yg
free sulphite/ml) was particularly sensitive whereas some strains
of Hafnia alvei (MIC = 210 - 264 yg/ml) and Serratia liquefaciens (MIC = 200 - 250 yg/ml) were relatively tolerant of the preservative.
Although such results tend to support this hypothesis, the range of
sensitivity of strains of Escherichia coli to sulphite was such
that the relative abundance of this species in the sulphited, but

299
not the unsulphited product was not due to the concentration of
sulphite alone. It should be stressed that sausage contains a
wealth of carbon and energy sources in the form of starch (Abbiss,
1978). Thus rusk (ca. 840 mg insoluble carbohydrate/g) and lean
pork (ca. 2.2 mg/g) are likely to contribute as substrates for
microbial growth. Because of the occurrence of two a-amylases
and a raaltase of meat origin, glucose and maltose are available
to the microbial association throughout storage of sausage (Abbiss,
1978). Although lipids are probably used also (c.f. accumulation
of valerate, Abbiss, 1978) there is probably a "sparing effect"
(Mossel and Ingram, 1955) on protein. Indeed it has been shown
recently (Dainty and Hibbard, 1983) that iso-valeric, iso-butyric
and 2-methylbutyric acids may be formed by Brochothrix thermosphacta from leucine, valine and iso leucine respectively and contribute to
spoilage odours (Dainty and Hibbard, 1980; Stanley et ai., 1981).
A 6-amylase has been detected in rusk also and probably contributes
to changes in the concentration of carbohydrate in sausage. As
amylases contain relatively few disulphide bridges they are resistant
to dénaturation by sulphite, but it is possible that hydroxy-
sulphonate derivatives are formed, removing sugars from sausage
as a consequence. Abbiss (1978) concluded, however, that the
concentrations of sugars (glucose, maltose, maltotriose and malto-
tetrose) found in fresh sausage were substantially greater than
those required theoretically to bind all the sulphite at point of
manufacture. As the presence of the preservative is always re
flected in the composition of the microbial association, by inference,
free sulphite must always be present. Moreover as lactate and
acetate accumulate in the unsulphited but diacetyl and acetoin
predominate in the sulphited sausage (Abbiss, 1978), the preservative

300,
appears, as suggested by Brown (1977) to influence the fermentation
pathways of members of the association. It is notable, moreover
that the addition of sulphite (ca. 450 pg total sulphite/g) to
minced beef or pork results in selection akin to that observed
in fresh sausage (Nychas, pers. comm), the growth of Pseudomonas fragi and Enterobacteriaceae being severely curtailed. Sulphite
results also in a sparing effect on the low concentrations (ca.
0.1 - 0.2%) of glucose, and eventual spoilage is manifested by
souring in sulphited mince cf. putrefaction in the control. The
evidence presented in the above discussion suggests that sulphite
elects for a particular type of microbial association in fresh
sausage, selectively inhibiting certain groups, e.g. the
Pseudomonadaceae and Enterobacteriaceae, whilst allowing others
e.g. the yeasts and Brochothrix thermosphacta to grow. This situation
directs attention to possible mechanisms of action of the preservative.
Gould and his co-workers (1983), for example, surmised that
sulphite probably reacts with enzymes, particularly those associated
with the tricarboxylic acid (TCA) cycle whereas others (e.g. Freese
et al., 1973; Warth, 1977) inferred a reaction as a "proton
ionophore". The oxidative function of the TCA cycle, mediated
by dehydrogenases, produces the reduced nucleotide, nicotinamide
adenine dinucleotide (NADH), subsequent reoxidation to NAD^ being
coupled to ATP production. Thus NADH may be considered not only
as an end-product, but as a repressor of citrate synthase (Fig. 85).
Weitzmann (1966) noted this phenomenon in Escherichia coli. The
citrate synthases of Gram-positive organisms, on the other hand,
are rarely affected by NADH (Weitzmann and Jones, 1968; Table 49).

301
Table 49. Regulatory patterns among bacterial citrate synthases'
NADH inhibition
AMP reactivation No AMP reactivation
Acinetobacter spp. Arizona arizonae
Moraxella spp. Aeromonas formicans
Pseudomonas spp. Erwinia uredovora
Escherichia coli
Hafnia alvei
Klebsiella spp.
Proteus spp.
Salmonella spp.
Serratia marcescens
No NADH inhibition
Arthrobacter spp.
Bacillus spp. Streptomyces spp.
Clostridium spp. Microbacterium thermo-sphactum
Micrococcus spp.Staphylococcus aureus
Nocardia spp.Kurthia zopfii
Adapted from Weitzmann (1980)

3 02 .
The repressed enzymes of strictly-aerobic Gram-negative bacteria
were reactivated by adenosine monophosphate (AMP); AMP had no
demonstrable action on those of facultatively-anaerobic Gram-
negative bacteria, (Table 49). It is noteworthy that these patterns
and those of sulphite-sensitivity appear to be common. Thus those
micro-organisms - the yeasts, Brochothrix thermosphacta, micrococci
and Kurthia zopfii (Dowdell and Board, 1971) - which are not in
fluenced by sulphite possess citrate synthases which are not inhi
bited by NADH. In contrast, the Enterobacteriaceae, markedly
sulphite-sensitive organisms, have citrate synthases which are
inhibited irreversibly (i.e. no AMP reactivation) by NADH. Those
organisms in which NADH inhibition is relieved by AMP, e.g.
Moraxella spp. and Pseudomonas spp. are ones which are inhibited
only by moderate concentrations of sulphite in broth and which
grow to a limited extent in sulphited sausage (Table 49).
Sulphite reaction with NAD^ that prevents energy-yielding oxidative
reactions is well known (Meyerhof et ai., 1938; Dupuy, 1959).
Indeed this reaction is most marked (Dupuy, 1959) at the pH values
found in sausages. Rehm (1964) and Wallnofer and Rehm (1965)
tentatively identified the NAD-dependent step from malate to
oxaloacetate in Escherichia coli as being the most probable target
for sulphite.
As well as citrate synthase, malate dehydrogenase and pyruvate
dehydrogenase are also subject to NADH inhibition and are thus
possible targets for sulphite (Fig. 85). As citrate synthase
activity is inhibited by a-oxoglutarate (Gottschalk, 1979), it is

303,
possible that feedback repression by high concentrations of a-oxo
glutarate may occur with sulphite blocking of the NAD-dependent
a-oxoglutarate succinyl-CoA step in the TCA cycle (Fig. 85) .
Such a lesion could result from an indirect reaction of sulphite
with NAD, or by direct inhibition of the a-oxoglutarate dehydrogenase
complex (Fig. 85). It is notable that the above reaction occurs only
in Enterobacteriaceae (Sanwal, 1970) - the most sulphite-sensitive
micro-organisms. During anaerobic growth, which might be expected
to occur in the centre of sausage, energy is generated by ferment
ation.
Thus in Enterobacteriaceae, a-oxoglutarate dehydrogenase
is absent and the TCA cycle is modified (Fig. 86). Although the
branched non-cyclic pathway satisfies the biosynthetic demands for
both a-oxoglutarate and succinyl-CoA (Amarasingham and Davies,
1965), sulphite may impede further utilization of a-oxoglutarate,
thereby inhibiting citrate synthase by feedback repression.
Although it is evident from the above discussion that sulphite
may inhibit aerobic metabolism, the Enterobacteriaceae are
capable also of anaerobic mixed acid and butanediol fermentations.
Organisms comprising the genera Escherichia, Salmonella and
Shigella ferment sugars to lactic, succinic and formic acids (Fig.
87) with the concomitant production of carbon dioxide, hydrogen
and ethanol. Species of the genera Enterobacter, Serratia and
Erwinia produce less acid but more carbon dioxide, ethanol and
2,3-butanediol (Fig. 88).
It is theoretically possible that sulphite will interfere with

304
dehydrogenase enzymes (Figs 87, 88) and may even combine with
acetaldehyde and remove the latter from subsequent reactions,
e.g. regeneration of oxidised NAD by ethanol dehydrogenase.
Shortage of NAD would prevent activity of glyceraldehyde-3-
phosphate dehydrogenase and therefore prevent flow of intermediates
via glycolysis and further decarboxylation of pyruvate. In this
respect it is noteworthy that sulphite prevents two features from
occurring in fresh sausage, (1) the rapid and extensive proliferation
of the Enterobacteriaceae and (2 ) a dramatic decrease in the pH
(pp. 129 - 130, Figs. 29 and 34). It follows that sulphite- pertur
bation of the metabolism of the Enterobacteriaceae may account for
the non-occurrence of both features noted above.
It needs to be stressed, however, that the above hypothesis
and that discussed later (pp. 305 - 307 ) do not explain fully the
anomolous results obtained with rifampicin-resistant Salmonella serotypes in sulphited broth stored at 4* and 12° C (pp. 152 - 154) .
Pseudomonadaceae use the Entner-Doudoroff pathway for sugar
degradation. As in the pentose phosphate cycle,glucose-6-phosphate,
is first dehydrogenated to yield 6-phosphogluconate (Fig. 89)
which in turn is converted by a dehydratase and an aldolase to
one molecule of 3-phosphoglyceraldehyde (3-PGA)and 1 molecule
of pyruvate. The 3-PGA can then be oxidised to pyruvate by the
enzymes of the Embden-Meyerhof pathway. Some pseudomonads, e.g.
Pseudomonas aeruginosa, Ps. fluorescens and Ps. putida, contain a
glucose dehydrogenase which converts glucose to gluconate. The
latter can be alternatively phosphorylated to 6-phosphogluconate
or further oxidised to 2-ketogluconate which is subsequently
phosphorylated and reduced to yield 6-phosphogluconate (Fig. 90).

305
Thus the membrane-bound glucose and gluconate dehydrogenases
facilitate the extracellular conversion of glucose into compounds
which are less preferred substrates for ccmpeting micro-organisms.
The pseudomonads are then capable of transporting and degrading
glucose, gluconate and 2-ketogluconate, As sulphite inhibits the
growth of pseudomonads in sausage and mince, whilst conserving
the level of glucose in these products, it is feasible that it
reversibly inhibits the dehydrogenase step from glucose-6-phosphate
to 6-P-gluconate (Entner-Doudoroff pathway) as well as limiting
the activities of glucose and gluconate dehydrogenases in extra
cellular glucose oxidation. One additional hypothesis which cam
be discussed in order to explain both the growth inhibition of
the Enterobacteriaceae and the successful colonization by Brocho
thrix thermosphacta of sausage needs comment.
It has been suggested (p. 52) that sulphite may
cause inhibition of growth by entering the cell, lowering the pH
and releasing protons. In an attempt to expel protons and re
establish the membrane proton gradient, the cell might expel ATP.
As internal acidification must be avoided, removal of protons
against a steep concentration gradient, is mandatory.
Such "proton export" is energy demanding and diminishes the amount
available for cell synthesis. As the growth rate is reduced, the
fraction of a cells' energy utilized for maintenance rises so
that energy available for additional proton-extrusion becomes
even less (Gould et al., 1983).
Michels et ai. (1979), on the other hand, have surmised that

3 06 ,
lactate excretion causes proton efflux thereby generating an appreciable
electrochemical proton gradient. Under conditions of homolactic
fermentation of glucose this gradient, if coupled to energy
transduction, would increase the theoretical ATP yield by ca. 30%.
Brochothrix thermosphacta grows well at 4 - 9°C (Rogers et al., 1980)
and 25°C (Kitchener et al., 1979) with L-lactate being the major
product of glucose breakdown. Thus it is proposed that during
metabolism of glucose to lactate,protons, derived from diffusion
of sulphite into the cell, are continually expelled thus sparing
ATP and permitting growth viz.:
SO. 2-
H s o r ^
Proton extrusion inBroctiothrix ttiermosptiacta
Protons
31 kal ine — acidic
Glucose
Protons Lactate ^ ^ If
iT 'w VLactate Protons
Such symport of protons by homofermentative micro-organisms in
the presence of high concentrations of sulphite would be of con
siderable advantage and may explain in part the significant
contribution by Brochothrix thermosphacta - and perhaps of

307
homofermentative lactobacilli (pp. 141 -145) also - to the microbial
association of sausage.
From the above discussion it is evident that the inclusion
of sulphite in sausage is largely ineffective against the spoilage
association. Thus although future work may elucidate the modus operand! of the preservative, there remains little prospect of
extending the shelf-life of the product. With this situation in
mind an attempt (pp. 308 - 322 ) was made to identify systems
which could be used as adjuncts to, or alternatives to sulphite.

3 08 .
COMMERCIAL IMPLICATIONS
It is evident from the results presented above that, in terms
of its influence on the major members of the microbial association,
sulphite ought not to be regarded as a preservative when used at
the legally accepted levels. Even with the minor members, sulphite
at these levels does not act only as a preservative but as an
elective agent also. In the light of this evidence, attempts were
made to identify^ tentatively^ systems that could be considered for
commercial use with the objective of retarding microbial growth in
sausages containing reduced amounts of sulphite.
The efficiency of two preservative systems were examined:
sulphiteiCOg and sulphite:potassium sorbate, these combinations
being included in 6 batches of sausage produced on a pilot scale
(Table 50). It needs to be stressed that such small scale production
of sausage seldom mimics, particularly from a microbiological stand
point, the commercial product. In spite of this reservation,
several important trends were evident in the results which may be
of significance to "the trade".
Sources and extent of contamination of ingredients of pilot scale
sausage
All of the meat ingredients were contaminated heavily with most
of the members of the microbial association, although streptococci
were present in low numbers only (Table 51). The relatively high
initial level of infection of the final sausage mix (ca. 10^ c.f.u./g)

309
•îü<Dü-HW5O+JWCO•H-P•HS(0W0)§■w3dü)luowQ)•g■PrOd
fduU)p)or4•H04M-4OCO-HP)•PWOtU
oLD
gH
0}pl(dAp0W (Ji
\S tn0 3-•H û)co o en 1 1 1 1 + +w O (d(d O wpi p0 CN fdÛ4 w
CN ^O >U \__ >
•H o'P'P O 1 1 + + 1 10 CNW _
d)Oc >•H \O 50) dPpo W fH•H 'p tJ* ••W 0) d <T» 1 + 1 + 1 +(d pi -H cr»
h -H c LnÆ 0
G Oc M §p) H <d S
P G ttîW W (/) '- 'c0
•Hpl•p
T3 —-< ^
dP
T3G œ iHp) tm " + 1 + 1 + 1•P d cr>Pî -p <j>Oc d IG
I—1 0p en 2en fd Hd G et:D w —
0 GPJ pl(d fda Ap P
CN CN G 0o o œ enU U+ + + +\ \ \
(D CN CN CN CN CN CNd O O O O O O0 co co e/) co C/3 wu 1 + 1 + 1 +
pGA
dr4 CN m Tf m CD
Upl(dPÛ

310,
Q)
enPPen
(duen
-PO1-4•HÛ4IPOen-PGeu•H
ug•HU-l0g•H•P(dc•p
-pG0üip0
gg
inagEh
pltdIp rP m Q C" C'
1— 1 un Q 00 q q enü Z(d r~ I f in r4 CD mm V
enT3G•pp CN e^ O Q m CN
CN 00 Q CD q l" enZ
eu œ Hf co r-4 r - in0 V0u
pen 8cpl C7\ 'f C'- O C'-G 'Ü CN Q 00 C'- <T> LGeu fd Z•p eu en TT m r4 'd' CN
T3 m(UuenGH
p0eu 00 VO Q "d* •d*
O C' CD O O>11-4 in en fn 1-4 •d* CNr4euPQ
.XP0 ■JeDe œ 1—i Tf Q 00 VD
O en Q m O 00 kOS 2fd fn LD r-4 LDeuP)
pl euP fd Ôc De fdP p De De eu0 ü en en De üU td De fd
æ: en en en •peu Qc 3 3 prp en M 0 en (Ud M 0 fd pjId •p 0 G u•p P ü b fd> Q) fd 0 Aen æ ; 4 P) 0 0rp pl PI 0 De ■3 Pfd en PJ eu 3 eupl fd 0 p Qj pl0 eu P fd Pl en GEh >H cq ip (0 Ûc H
eur-4Deen
en\Pipü
tn0
Gg
Ig

311,
probably reflected the substantial populations in the components of
the product.
Loss of sulphite
The inclusion of CO^ (Fig. 78 b) or sorbate (Fig. 78 c) reduced
significantly both the rate and extent of loss of total sulphite.
B o t h of these additives also reduced significantly the initial extent
of binding of free sulphite, CO^ being more effective than sorbate
during storage for 7 days at 4° or 10°C (Table 52). Thus with storage
at 4°C in the former case, the concentration of active preservative
was maintained within 50 yg of the initial level, whereas in the
latter case, ca. 150 yg had been bound by day 7 (Fig 78 b, c).
Influence of sulphite concentration
As in batches of sulphited or unsulphited sausage examined
previously (pp. 125 - 132) , the temperature of storage influenced
directly and the inclusion of sulphite inversely, the rate and
extent of growth of the total viable count (Fig. 79a). The yeasts
grew in sulphited sausage at 10°C, but not at 4°C; they did not
grow at either temperature in unsulphited sausage (Fig. 80a).
Although sulphite delayed the onset of growth of Lactobacillus spp. at 4°C for 24 h,the rate and extent of growth thereafter
was not influenced by sulphite concentration and large populations
were formed by day 7 (Fig. 81 a). The pseudomonads did not grow
in either the preserved or unpreserved sausage during the course
of the experiment although the populations in sulphited sausage

312
îgQJ 4— ^t" 00 TT r-4 O 00 C'-(0 »x> r" n iHU0+JW440G0•H-P ¥(tJ «ü* 1 ^ l o 13 ^ r- ^ 00a
Q)-P•iHA1—13MCDQJ QJU p4-1 3-P4-1 n)0 PQ)O' QaG B•H QJPG U O ^ O O•H QJ 0 I—1 1-4 1—1h 3G P0 0-PQ) W+JdUP0WP0CNOu
4-10 QJ4-)Q) X tOÜ U hG -p PQJ (TS CN 03 PQ O w1-4 U4-1 44 + +G 0H CN CN CNc O O O0 W CO CO•HCN P» 4 - 4 - 4 -LD •Hen0) 01—i Û4Er3 0Eh u
o r->1fO >1TJ (d+Jce) p(dQJ4-> eu•P Ph ■HC4 X1-4 A3 r4en 3enr-4(d 1-4-P (d0 P4-J 0P440 p
0QJen QJ(d en4-1 (dG PQJ Gü QJP UQJ PD4 QJA(d fden(d entdeu euP P•H •HXIeu A1-4 1—43 3en en
eu QJeu eup pCL| k

313
were invariably smaller than those in unsulphited ones (Fig. 82 a).
Sulphite reduced significantly the rate and extent of multiplication
of the Enterobacteriaceae, nevertheless a significant increase in
the numbers of these organisms in preserved sausage stored at 4°C
or 10°C occurred by the 7th day of storage, (Fig. 83 a). This
situation may be due in part to the rapid loss of free sulphite from
the product, (Fig. 78 a). Although temperature of storage influenced
directly the rate and extent of acid formation, higher temperatures
favouring a marked acid drift, sulphite prevented substantial pro
duction of acid even at 10°C, (Fig. 84 a).
Influence of CO concentration
Addition of CO^ to sulphited sausage delayed the onset of an
increase in the total viable count for up to 48 h at 4° or 10°C,
thereafter the rate of growth at 4°C was substantially reduced,
(Fig. 79 b). Inclusion of CO^ in unsulphited sausage did not in
fluence the rate or extent of increase of the total viable count
(Fig. 79 b). Although a combination of CO^ and sulphite reduced
significantly the size of the yeast populations at 4°C during
the 48 h post-manufacture, these organisms grew subsequently,
albeit poorly (Fig. 80 b). The presence of CO^ in the unsulphited
product stored at 4°c stimulated growth, a situation not evident
at the higher temperature of storage (Fig. 80 b). Neither CO^
in the unsulphited sausage stored at 4° or 10°C nor in the sulphited
sausage stored at 10°C was effective in preventing substantial
proliferation of lactobacilli, although the combination of CO^,
sulphite and storage at 4°C curtailed growth (Fig. 81 b). Carbon

3 14 .
dioxide alone retarded the growth rate of the pseudomonads such that
their populations were lower in the unsulphited product at the 5th
day of storage, (Fig. 82 b). As in sausage containing sulphite only,
the addition of CO^ prevented growth of pseudomonads for up to 7
days (Fig. 82 b). Carbon dioxide was effective in retarding the
rate of growth of the Enterobacteriaceae in unsulphited sausages and
prevented multiplication completely in sulphited ones (Fig. 83 b) .
The drifts in pH of preserved or unpreserved sausages containing
CO^ were not substantially different from those of 'uncarbonated'
batches (Fig. 84 b).
Influence of sorbate
The rate and extent of growth of micro-organisms was drastically
reduced in the presence of sorbate and, in unsulphited sausage
stored at 4°C for example, no significant increase in numbers was
evident during the period of storage (Fig. 79 c). The combination
of sulphite, sorbate and chilled storage reduced significantly
the size of the yeast population for upwards of 48 h post-manu-
facture, thereafter they began to grow (Fig. 80 c). Sorbate alone,
or in combination with sulphite, was not lethal at 4°C and 10°C
or 10°C respectively, but it did prevent the growth of the yeasts
for up to 7 d (Fig. 80 c). The rate of multiplication of members
of the genus Lactobacillus was retarded significantly by the
addition of sorbate, especially at chill temperatures (Fig. 81 c).
The numbers of pseudomonads were reduced to the greatest extent in
sausages containing both sulphite and sorbate although after 4 days
they resumed growth, albeit slowly, in all batches of sausage,
(Fig. 82 c). As with the addition of CO^, sorbate acted synergis-

3 15 .
tically with sulphite to maintain the Enterobacteriaceae in a
quiescent state at 4° or 10°C for up to 7 days; sorbate alone
was effective also in preventing growth, (Fig. 83 c). The pH
of sausages containing sorbate did not change significantly at o oeither 4 or 10 C over the period of storage, a situation which, in
the absence of sulphite, was probably due to the suppression of
growth of the acid-producing lactobacilli and Enterobacteriaceae,
(Fig. 84 c).

316
(UenfOcotM
(13UC/)I+J0r-4
■HA
C•rH0)-P•Hx:
(T3-P0p)?(T30303PtpipOWü)0c003p
5(üp03
03-PP003üC033t—\PC
CO03pDenpt,
£•H
'Ü03PS§1-4;w03
C3)Sen
>
dPOCN
03-Pg•H'Ü
§X3UXp•H
P5 g
r—4
6w03enfdUies(d(33
03P•H§:)(3)
(dP5
£
% (3) 0 r—iU
en
en3.o
03pfdpps§
•H(33(333
â
03P•HÛ3rHS
0303PP(33I—I
£
I
o" o"' ( f un
O <
O8 SO
aOesnes ^_6^0S 6n<
1o o o
Al
aBesnes ^OS 6n<
aBesnes ^_6^0S 6f^
03|

317
tuuMI-PO
•Hû
C•rH-PC00ü
Xitu•H>tu
+ J
5tu
x:V
4-1O
rC
0p
coU)
p
50)
4-H0
Q)PP-Ptup
10)-ptp0tuudtudrH4HCH
<y\[>
Q)Pg>
•H
ntu-pc
Ir H
I(Utuwdtuw
tu
dPOCM
(UTU• H
g• PT3
IPtuU
-P•H
■S
(UIrpâg(Uentutn3CO
a
(UentuCOdtuCO
enen3 .
q
CM
(U
A
UCO
•pCOCOtu-pâXIV•p5
(US ’CO
3CO
s•pAiHSCO
rH
5
Ko
(U-p
Bog
(UsCOsCO
%p)•pArHdCOdd
COiH5
D
0r HU
u
o <
- J o u |
OUafiesnes _6 nyu 5oi
OUa6esnes^_6 n j ] 6o)
tu I
OLafiesnes ^_6 n ; ] 6o)

318
T30) <UCn ■P<d Gw Q)n(0m rHC4<u ArH P(Q 0)Utn Q)1 Cn+J ft0 w1— 1 P•H PA Mn) UC•H TJCw ft-PMIt 5(U \>1 3V4 dP0 OCN5
Q)0 'Op -HCr X0C •rl0 730) Ccn 0 Cnft A \u P pi0 P :a4-1 uw OQ4-1 p O0 ■H CM0)p 73ID (P+J P Pft G Pp 0) A(U Pa 0S i-H cn(U A-P AP4-1 w •p0 0)(U œQJ 5^ ft piÜ ft P •HC W 0 k3 P A0 ft GrH CO •H4H PC •H WH rQ <
4 <
01a5esnes ^_5 6 o i
L J
01 ,a6esnes ^_6 n ; o 6o)
tn
u I
(t3 I
aBesnes , 6 ny ] 6o|

3 1 9 .
œ>4p
-H
T30)+JgI0) rHr—1 Afd Au 3
U) to■p tu0 crnH fd•H toa DfdtofdC u
•HTJc
Ù, (d0410to3M"MÜ af>fd 04 OJ04J0 tufd A•p
X(4-1 00 •HT)A
-P C3 00 AU PCn fduC0 A
+Jeu •Pen enfd \P 'D en0 tu p.-P Ato Ctutp e0 turH CN(U au Û43 d tu4J to Afd fdp tu A(U en P& fd 0e to totu dp fdto enip •P0 to
A to(U (d enu A •pc tu 0 tl(U en3 fd crH to A •pfp d Ac fd •P to
H to 5 <
01.a 6 e s n e s ^_6 n j 3 6 o |
LA
Al
01,a G e s n e s ^^_G n ^ 3 G o ]
0001,
aGesnes _G n j 3 Go|

320,
mCP(C3m3(CScn +J•H0)1—1(T3 T3U <Ucn ■P1 d+J OJ0 Er4 (U-r4 rHA 0404
dn3 cnC (D•H CPtocn04 d04 tocn cncn VCQ ÜC0 T36 d0 (0%30) 5cn \Û4m àP0 oCN
(U0 T5u •HCP X
0c •H0 T3(U dCP 0fC3 X)U P0 to■0 ucn CP\
-p CP0 •H 3.Q) ou T> QD 0) q■P -P(CJ d CN(U(U E04 <u CDE rH -PÛJ 04 to4J 04
d P CP4-1 cn 0 r-0 cn
CDm S' E CPu to d •Hd cn •H0) d cn0 (0 cn drH cn to •H4-4 -Pc 0 cnH A 04 <CN00CDU3CP•H
a6esnes _6 n ; ] ®'’6o]
i<o'
u|
ffl I
01,aGesnes |_6 n ; ] 5oi

3 2 1
Td0)•Pa(d
(U sI-H <d(0 I-HÜ AU) ft1 0
■p w0r-H (d
•H cncu fd
COP
fd fdCO
c•rH
oQJfd V
au fdfd
•rHu ?mP ?Üfd <)P
A o0 CMP(U•p 0)c Tdw •H
Xf+H 00 •p
x;c
0 Sp pCr> fd
uC0
•POJ •pcn 5 cnfdP T i cn0 (d 3.P •Pm c Q
fd O<+H s O0 (d
r— 1 CM(U ftp ft0 P (dP CO •Pfd fdp (d 13(d S' PA fd 0£ CO CO0) 3•P fd
CO cnM-4 •p c^0 CO
A CO(d fd cnU P •pC (d 0 k(d cn ft3 fd a1—1 CO Æ •pM-1 P Pa fd •p COH CO 3 <
mCO
<V
<6 <*
• o
un m
Ul
OLa6esnes n; ] 6oj
q<
< -
at
OLa G e s n e s n | D Go)
(d 1
OLaGesnes _G n ; ] 6o|

322
-P• H
T30)rO +JC(Ü0) eCP <Ur— !W a0 Û4(ti 3m m0) CDI— 1 u<ftj n3o cnin p1 n3
cn0r4•H uo , T ?CfO IT)C•H4J ?<4-1•H dP
o'Ü C Ntc& 0)' OG • H0 X0<U • HCP T 3fOu0 s4J Aw CP\<4-1 u CP0 p.0) -pX4 •HG4J(Q C Nd) -P& c (US cu -pQ) s Id4J Q) Ar H p 0 ^<4-4 A 00 A MG<u cn CPÜ • Ha Q) •r4 ko C P cna Id cn G1— 4 w Id •H<4-4 0 PC Id 0 WH cn A <
C OQ)
■H
U|
( a B e s n e s ; o a ^ e u a B o w o q ^ j ] i )
At
•O d 4
OJ I
(aBesnes a4eua6oujoq^_oi ) ^d

323
■KC0■H-PUfd
XIouA
(U
uo(/)
Oj
8to Ll>< .•501 Lj
I\oAO)
/QJ
w
(U+j•H1—40m0U)Q)
4-»•HW
■S«ueu
Q)4JOC4JOs
Q)Q) 4->4J fd<d 1-4u njfd E-P3 Of—4 1—1en0X d)0 en1 fdS pfd«X Ee>4-4d)ü) enfdC(Uen d)0 cnp fdT) C>1 dJÆ enQ) 0T) pd) >14J x:fd d)xj•H d)ü 4-)0 fdm c•H •HüLD u0cnt »,œ(Ucnfd4-> d)fd cnp fdT3 G>1 •Hx: 0d) •H4-) Xfd 4->4J•H (UC 4J0 fdP Cfd •H1 Uen u en•p G r~ü en enI—1CM XXX fdCD Xd) r—1 ucn eu cnfd E VX 0 X4-> U 0C O>1 dJ dJcn cn en Efd fd 0dJ G G P4-) d) d) (pfdp S' S'V4-> p p CD•H 'D XU >1 >1 euX X fdd) dJ T)f-H T) 'e) c

324,
acetyl - Co A
oxaloacetate/
malate
fumarate
s cinate
citrate \ISO-citrate
\ __________cc-oxoglutarate
succinyl-CoA
Figure 8 6 . Modified TCA cycle operative in Escherichia coli
growing anaerobically.

325
Figure 87. The mixed acid fermentation in Enterobacteriaceae and probable
sites of sulphite action*
* Adapted from Gottschalk (1979) _________glucose
H
10
lactateH2.
Y ^ D P. . S a t p
i^PEP1 r"A D P
<Urppyruvate
3 ^CoA
acetaldehydeo formate
ethanol
acetyl-CoA
acetateFootnote ; O Probable sites of sulphite action
1 Embden-Meyerhof pathway enzymes
2 lactate dehydrogenase
3 pyruvate-formate lyase
4 formate-hydrogen lyase
5 acetaldehyde dehydrogenase
6 alcohol dehydrogenase
7 phosphotransacetylase
8 acetate kinase
9 PEP carboxylase
1 0 malate dehydrogenase , fumarase, fumarase reductase

326
Footnote
Figure 8 8 . The butanediol fermentation in Enterobacteriaceae
and probable sites of sulphite action*
O Probable sites of sulphite action
I - lO as in Fig. 87
I I a -acetolactate synthase
1 2 a -acetolactate decarboxylase
13 2,3-butanediol dehydrogenase glucose
lactateH
WyfADP^ATP
■pyruvate
Hacetaldehyde*o^acetyl - CoA formate
ettianolCO, H,
<y: acetolactate
12 C0„
acetoin
2,3 butanediol
Adapted from Gottschalk (1979)

327
M<D4-)•HenQ)r—(
IàT)C03Q)03QJÜ
"Q03q0033QJ
QJ
+J•S§oS03QJ
ÆenSa
rC4-J03a4-14-1034aDO?I4QJC4-JgQJÆEhon00QJ343en•Ht,
*g•H-PU03
•H
"âr 4aen
440
OOLJm
— L-j —
enen
Icn4J4-J
gi3444
4-J
§•T3<
wIC? s - iLJ— LJ ,^ rn
8 ? LJ— LJ□C^ o— ILJ — LJ□C
©oI zc LJ — LJni
C3
ca
m m o oS - u L ^ ? LJ — LJ
I m o
g-
mo '^CN I 00 LJ — LJm
oIILJJZ
mo mo ©LJ LJm I o m
LJmI mLJ LJ
'CNJ
13CL-Q
I
I\o
X)Qj
§ QJ4J
oo
QJen03I—IoT3
G0
•r4 O4J QJ 04U en03 03
G euQJ QJ en4-J en 03 QJ•H 0 4-J 4J
34 03 03A 03 34 G
rH >1 03 0P rG >1 Uen QJ ,G P
03 QJ r—144 03 en0 QJ 1
4-J eu 04en 03 4J 1QJ .G 03 304-J A G 1•H en 0 >1en 0 U X
Æ P 0eu 04 nH QJ
1—1 1 (n 03£>' eO Q 103 1 X i rot l QJ 04 10 en en 034 0 0 4-JA ü f : q
0 04
O r—4o 13D 1OJ
r—1 eN m

328,
0)in0uar—Icn
4-1OCO•H■p
•HX0>4to
(1)u(04J
0)5c•rH
o•rH-PUto0)HJ•rH
Ar—IDin
tw0in(U4-)•Hin
AIpa,
I§<uVh•rH
o•p■S
toc0BCnS(1)1
CN
?to(Üpis0ÜCirHcn
in8Pr— Icn
tpO§•Hinp(U>coÜ
nH0)utoa•H
Tlctoin
tOÎp(UinOh
'C
IZ)
c\|
II
Q.C3
CJ
I
I
ICSl
slO
-aIII
CL
¥S IfUCL
O' T DZ3OQc_cuc§•H•p
•H
gPin
tp0in0)•P•Hin
Q)•PPPO0 prH
10Æ (U Û4 in1 to VO •P8 I0) (1) PI
C Nin
Sipc•H0)c0upr Hcn
88I
C N
Q)in30)S'p
"373Q)•P
tIVOI0)inoPi—Hcn
Xi Q)0 inp P111 P•Ho Q) X
in (UtO pP to•H pQ) X 0•P 0 u0 X pC <u rH•P x: cn00k pH CN
cncn
rX1—4uin80OPip%•PI<

329
X•HII
•gS
Q)enPenPPtnP0ip <P
Q)tn 0C CQ0•p 'd+J CP pu•HtP p•H 0ü Çk0AtnT3ppDGP-PM
CDgœPfOœ
tnTP
■P p(0 ■p •H
■p 0 (d p tu(0 e (U -Pip e TP p
-P >1 tu x;C .ic: P rH TP N en Û4eu p 0 (ü i-H (d •H p tn•H 0 fO -P tu tu P •p 0np A X) g x> X: P p x:tu 3 tl) o QhP g \ Xi x; Xi •P tn >,en (0 P 0) tn p p tn (d rHC 0) 0 U P 0 0 (d O 0M PI Pk H (ü Ck w PU
Q)S'sM
TT O ro m o O OVÛ CN CN 'd' 1— 1CN CN rH I— 1
mO 8
"vT vo m Md 1—1 tn CN 1—1CN rH in
•H'O
-p 73+j fd prd tu •HUH E P
-P XiP p Xi TP 73 tu entu p u p tu u p
•H 1-4 fd tu tn •H •HT3 ip XI X tn \ ptu tu s p p 0P M H Xi Xi u p Xi (U tnen tu p p 0 CÜ* 0 tn -p Pp tu 0 0 p rH P p tuH m cu cu s k Pi 2 w
8
IenIrHO

330,
Appendix 2
Isolation, identification and maintenance media
Salmonella
Buffered peptone water (BPW), g/&: Peptone (Oxoid L37), 10; sodium
chloride, 5; disodium hydrogen phosphate (Na2 HP0 ^.1211 0, 9;
potassium dihydrogen phosphate, 1.5; distilled water to 1 I.
pH 7.0 at 25°C after sterilization at 121°C for 15 rain.
Lactose broth, g/£: 'Lab Lemco' beef extract, 5; peptone (Oxoid
L37), 5; lactose, 5; distilled water to 1 &. pH 6.7 at 25°C
after sterilization at 121°C for 15 rain.
Peptone water, g/£ : Peptone (Oxoid L37), 1; sodium chloride, 5;
distilled water to 1 £. pH 7.2 at 25°C after sterilization at
121°C for 15 rain.
Tetrathionate broth, Tetrathionate broth base (Difco B104), 46 g;
distilled water to 1 £. Heated to 100°C, cooled to 25°C. Iodine
solution (iodine, 6 g; potassium iodide, 5 g; distilled water
to 2 0 ml), 2 0 ml added to broth base.
Selenite broth, g/£: selenite broth (Oxoid CM 39), 23; distilled
water to 1 £. Heated to 100°C for 10 rain.
Lysine iron cystine neutral red (LICNR) broth, g/£ ; L-lysine, 10;
tryptone (Oxoid L42), 5; yeast extract (Oxoid L21), 3; mannitol,
5; D-glucose, 1; salicin, 1; ferric ammonium citrate, 0.5;
sodium thiosulphate, 0.1; L-cystine, 0.1; neutral red, 0.025;

331
distilled water to 1 £. pH adjusted to 6.2, medium heated to 100°C,
cooled to 25°C.
Brilliant green agar (EGA), EGA (Oxoid CM 329), 52 g; distilled
water to 1£, heated to 100°C, cooled to 45°C and poured (20 ml)
into Petri-dishes.
Bismuth sulphite agar (ESA), ESA (Oxoid CM 201), 52 g; distilled
water to 1£, heated to 100°C, cooled to 45°C, mixed and poured
(25 ml) into Petri-dishes. Medium stored at 4-6°C for 3 d before
use.
Desoxycholate citrate agar (DCA), DCA (Difco B274), 70 g; distilled
water to 1£, heated to lOO^C, cooled to 45°C, and poured (20 ml)
into Petri-dishes.
Kohns medium, No. I. (Oxoid CM 179); No. II, (Oxoid CM 181) made
up and used according to manufacturers recommendations.
Lactic acid bacteria
Dowdell and Board's (1968) modification of Keddies (1951) medium (g/£):
Peptone (Difco B118), 5; Lab Lemco (OXoid L29), 5; yeast extract
(Oxoid L21), 5; D-glucose, 10; manganous sulphate (MnS0 ^.4 H2 0 ),
0.1; Tween 80, 0.5 ml; potassium citrate, 1; agar No. 2 (Lab M),
15; cycloheximide, 50 mg ; distilled water to 1£. pH adjusted to
5.4 before sterilization at 121°C for 15 min. Medium cooled to 50°C,
added (9 vols.) to 2 M acetate buffer (pH 5.4; 1 vol.), mixed

332.
and poured (20 ml) into Petri-dishes.
Whittenbury's (1963) medium. Lab Lemco (Oxoid L29), 5 g; peptone
(Difco B 118)/ 5 g; yeast extract (Oxoid L21), 5g; Tween 80,
0.5 ml; D-glucose, 0.5% w/v; agar No. 2 (Lab M), 1.5% w/v;
manganous sulphate (MnS0 ^.4 H 2 0 ), 0 .0 1 % w/v; tap water to 1 £.
Bromocresol purple (14 ml., 1.6% w/v in ethanol) was added, the
pH adjusted to 6.9 and sterilization was at 121°C for 15 min.
Whittenbury*s medium (for carbohydrate utilization tests). Above
medium excluding D-glucose. Filter-sterilized carbohydrate (1%
w/v) added aseptically to sterilized basal medium at 55°C.
Streptococcus medium (SM), g/£: Tryptone soya broth (Oxoid CM
129), 12; Lab Lemco (Oxoid L29), 2.5; yeast extract (Oxoid 21),
5; TRIS buffer, 3; hemin, 0.01; cystine, 0.4; distilled water
to 1 £. pH adjusted to 7.6, sterilization at 121°C for 15 min.
Kneteman's (1947) hydrogen peroxide medium. The medium of Whittenbury
(1963) vide supra was overlaid to a depth of ca. 2 mm with the
same containing manganese dioxide (4% w/v).
Arginine medium (g/£): Tryptone (Oxoid L42), 5; yeast extract
(Oxoid L21), 5; dipotassium hydrogen phosphate, 2; L-arginine
dihydrochloride, 3; D-glucose, 0.5; agar No. 2 (Lab M), 15;
distilled water to 1 £. pH adjusted to 7.0 and sterilized at 115°C
for 1 0 min.

333
Bile agar (g/£): Tryptone (Oxoid L42), 5; yeast extract (Oxoid
L21), 5; dipotassium hydrogen phosphate, 2; bile (40% w/v);
D-glucose, 0.5; agar No. 2 (Lab M), 15; distilled water to
1£. pH adjusted to 7.0 and sterilized at 115°C for 10 min.
Todd-Hewitt agar. Todd-Hewitt broth (Oxoid CM 189), 36.4 g ;
agar No. 3 (Oxoid L13), 15 g; distilled water to 1£; sterilized
at 115°C for 1 0 min.
Robertson's cooked meat medium. Fresh pork (500 g) was minced
(Kenwood A901) and simmered (1 h) in distilled water (500 ml)
containing IN NaOH (2 ml). The slurry was filtered through cheese
cloth and samples (ca. 5 g) of dried meat, distributed into 1 oz
screw-capped bottles. Ten ml of Lab Lemco (Oxoid L29) broth was
added to each bottle. Sterilization was at 115°C for 10 min.
Yeasts
Malt extract agar. Malt extract (Difco B 186) , 30 g ; agar No. 3
(Oxoid L13), 15 g ; distilled water to 1£. Sterilization at 121°C
for 15 min.

334,
Appendix 3.
Media used in sulphite tolerance work
All micro-organisms: Tryptone soya broth (Oxoid CM 129) or
Nutrient broth (Oxoid CM 1).
Lactic acid bacteria: Buffered Whittenbury's broth. Lab Lemco
(Oxoid L 29), 5 g; yeast extract (Oxoid L 21), 5 g; peptone (Oxoid
L 37), 5 g; manganous sulphate (MnSO^.411 0) , 0.01% w/v; Tween 80,
0.5 ml; tri-sodium citrate, 2.5 g; sodium dihydrogen phosphate
195 ml; disodium hydrogen phosphate, 305 ml; distilled water to 1 £,
Sterilization was at 121°C for 15 min. Filter-sterilized D-glucose
(1% w/v) was added to the basal medium at 25°C.
Yeasts ; Yeast nitrogen base-glucose broth. Yeast nitrogen base
(Difco B 392), 6.7 g; D-glucose, 10 g; sodium dihydrogen phosphate
+ disodium hydrogen phosphate buffer (1 0 % v/v); distilled water
to 1£. pH adjusted to 7.0, medium filter-sterilized.
Enterobacteriaceae: Modified glucose-glutamate medium (MGGM). (g/£);
sodium glutamate, 12.7; sodium formate, 0.5; L-cystine, 0.04;
L-aspartate, 0.048; L+ arginine, 0.04; thiamine, 0.002; nicotinic
acid, 0 .0 0 2 ; pantothenic acid, 0 .0 0 2 ; magnesium sulphate, 0 .2 ,
ferric ammonium citrate, 0 .0 2 ; calcium chloride, 0 .0 2 ; ammonium
chloride, 2.5; D-glucose, 20. pH buffered at 7.0 with sodium
dihydrogen phosphate (0.2 M) and disodium hydrogen phosphate
(0.2 M) at a final concentration of 20% (v/v). Medium was filter-
sterilized.

335.
Pseudomonas: Palleroni and Doudoroff's (1972) medium. Ammonium
chloride, 0.1%; magnesium sulphate, 0.05%; ferric ammonium citrate
0.0005%; D-glucose, 1%; phosphate buffer (M/30) to pH 7.0.
Medium was filter-sterilized.
Brochothrix theimosphacta. Grau's (1980) medium.
Appendix 4
Nilox scrubbers
In order to remove the last traces of oxygen from high purity
white spot dinitrogen (Air Products, Bristol), 3 Dreschel bottles
containing brown Nilox (2 bottles) and blue Nilox (1 bottle)
scrubbers were arranged in series. Composition of brown Nilox
(g): potassium hydroxide, 200; 1,2-naphthoquinone 4 sulphonic
acid, (sodium salt), 10; lead wool, 250; distilled water, 900.
Composition of blue Nilox (g): chromic sulphate, 300; sulphuric
acid (10 M), 45; zinc granules, 250; distilled water, 900.

336
REFERENCES
Abbiss, J.S. (1978). Enzyme-mediated changes in carbohydrate of
British fresh sausage. PhD Thesis, Univ. of Bath, England.
Abel, E. (1913). Inaktivierung eines nicht-Kolloiden, anorganischen
katalysators durch temperatursteigerung. Monatsh. 1361-1391.
Acton, J.C. and Saffle, R.L. (1972). Film observations at an oil-
water interface. J. Food Soi. 37, 795-796.VWAdam, W. (1957). Preparation and pasteurization of fishmeal in
relation to the content of Salmonella bacteria. Berl, Miinch.
tierërztl. Wschr. 49.
Amarasingham C.R. and Davies, B.D. (1965). Regulation of a-ketogluta-
rate dehydrogenase formation in Escherichia coli. J. hiol.
Chem. 240, 3664-3667.
Anderson, E.S., Galbraith, N.S. and Taylor, C.E.D. (1961).
An outbreak of human infection due to Salmonella typhimurium
phage type 20a associated with infection in calves. Lancet
£, 854.
Anderton, J.I. and Locke, D.J. (1956). Studies in the manufacture
and keeping qualities of sausages. B.F.M.I.R.A. Res. Rpt.
No. 72.
Andrews, W.H. and Presnell, M.W. (1972). Rapid recovery of
Escherichia coli from estuarine water. Appl. Microbiol.
23, 521-523.
Anon. (1947). Spec. Rep. Ser. med. Res. Coun. London , No. 260.
Anon (1950). Food poisoning in England and Wales, 1941-1948.
Mon. Bull. Minist. Hlth. 9, 148-158.
Anon (1951). Editorial Board, Judicial Commission. Status of the

337
generic name Bacterium, the specific name Bacterium coli
and the family name Bacteriaceae. Preliminary statement,
file No. 5. Int. Bull. Bacteriol. Nomenclat. Taxon 1,vv^
64-79.
Anon (1955). Food poisoning in England and Wales, 1953. Mon. Bull.
Minist. Hlth 14, 34-42.
Anon (1957). In 'Manual of Microbiological Methods' eds. Soc.
American Bacteriologists McGraw-Hill, Co., Inc., New York
and London.
Anon (1958). Judicial Commission. Opinion 15. Conservation of
the family name Enterobacteriaceae, of the name of the type
genus and designation of the type species. Int. Bull.
Bacteriol. Nomencl. Taxon 8, 73-74.
Anon (1959). Food poisoning in England and Wales,1958. Mon. Bull.
Minist. Hlth 18, 169-178.r-S A .
Anon (1961). Food poisoning in England and Wales, 1960. Mon.
Bull. Minist. Hlth 20, 160-170.
Anon (1964). Salmonellae in abattoirs, butcher's shops and home-
produced meat, and their relation to human infection. Rpt.
of P.H.L.S. Working Group . J. Hyg. Camb., 62, 283.
Anon (1976). In "Recommended procedures for the examination of
water and waste-water" 14th edition. Am. Publ. Hlth. Assoc.
New York.
Arnold, L. (1928). The passage of living bacteria through the wall
of the intestine and the influence of diet and climate upon
intestinal auto-infection. Am. J. Hyg. 8, 604-663.
Asakawa,Y ., Akahane, S., Shiozawa, K. and Honma, T. (1979).
Investigations of source and route of Yersinia enterocolitica

338
infection. Contr. Microbiol. Immunol. 5, 115-121.
Ashworth, J., Jarvis, B., Rawlinson, A.P. and Wood, J.M. (1974).
The replacement of sulphur dioxide as the microbial preservative
in British fresh sausage. B.F.M.I.R.A. Res. Rpt. No. 207.
Aulisio, C.C.G., Mehlman, I.J. and Sanders, A.C. (1980). Alkali
method for rapid recovery of Yersinia enterocolitica and
Yersinia pseudotuberculosis from foods. Appl. and Environ.
Microbiol. 39, 135-140.
Ayres, J.C. (1955). Microbial implications in the handling,
slaughtering and dressing of meat animals. Adv. Fd. Res.
£, 109-161.
Ayres, J.C. (1960). Temperature relationships and some other
characteristics of the microbial flora developing on refrig
erated beef. Food Research 25, 1.
Ayres, J.C. (1960a). The relationships of organisms of the genus
Pseudomonas to the spoilage of meat, poultry and eggs.
Journal of appl. Bact. 23, (3), 471-486.
Babich, H. and Stotzky, G . (1974). Air pollution and microbial
ecology. Crit. Rev. Environ. Contr. 4, 353-421.
Babich, H. and Stotzky, G. (1978). Influence of pH on inhibition
of bacteria, fungi and coliphages by bisulfite and sulfite.
Environ. Res. 15, 405-417.
Bailey, P.L. and Riley, M. (1975). Performance characteristics
of gas-sensing membrane probes. Analyst 100, 145.
Baird-Parker, A.C. (1980). In "Microbial ecology of foods" Vol. I,
(eds. I.C.M.S.F. editorial board). Academic Press, London,
126-135.
Barker, S.B. and Sommerson, W.H. (1941). The colorimetric determination

339
of lactic acid in biological material. J. hiol, Chem. 138,
535-554.
Barksdale, L., Laneelle, M.A., Pollice, M.C., Asselineau, J.,
Welby, M., and Norgard, M.V. (1979). Biological and
chemical basis for the reclassification of Microbacterium
flavum Orla-Jensen as Corynebacterium flavescens nom. nov.
Int. J. Syst. Bacteriol. 29, 222-233.
Barlow, J. and Kitchell, A.G. (1966). A note on the spoilage
of pre-packed lamb chops by Microbacterium thermosphactum,
J. appl. Bact. 29, 185-188.
Barnes, E.H. (1976). Microbiological problems of poultry at
refrigerator temperatures - a review. J. Sci. Food
Agricult. 27, 777-782.
Barnes, E.M. and Shrimpton, D.H. (1968). The effect of processing
and marketing procedures on the bacteriological conditions
and shelf-life of eviscerated turkeys. Brit. Poult. Sci.
£, 243-251.
Barnes, E.M., Mead, G.C., Impey, C.S. and Adams, B.W. (1979).
Spoilage organisms of refrigerated poultry meat. In "Cold
tolerant microbes in spoilage and the environment" (eds.
Russell, A.D. and Fuller, R.) Academic Press, London,
102-116.
Barren, R.A.E. (1982) . Isolations of salmonellas from human,
food and environmental sources in the Manchester area:
1976-1980. J. Hyg. 88, 403-411.
Beech, F.W., Davenport, R.R., Goswell, R.W., and Burnett, J.K.
(1968). Two simplified schemes for identifying yeast
cultures. In "Identification methods for Microbiologists,

340.
Part B" (eds. Gibbs, B.M. and Shapton, D.A.) Academic Press,
London.
Bergey, D.H., Breed, R.S. and Murray, E.G.D. (1939). In 'Bergey's
Manual of Determinative Bacteriology' 5th edition. Williams
and Wilkins Co. Baltimore, U.S.A.
Bertolacini, R.J. and Barney II, J.E. (1957). Colorimetric
determination of sulfate with barium chloranilate. Anal.
Chem. 29, 281-283.
Bevan-Jones, H, Farkas, G ., Ghosh, A.C. and Hobbs, B.C. (1954).
Salmonella brandenburg: an epidemiological study. Mon. Bull.
Minist. Hlth. 23, 162.v\ANBiemuller, G.W., Carpenter, J.A. and Reynolds, A.E. (1973).
Reduction of bacteria on pork carcasses. J. Fd. Sci. 38,VVwi
261-263.
Bissonette, G.K., Jezeski, J.J., McFeters, G.A. and Stuart, D.G.
(1975). Influence of environmental stress on enumeration
of indicator bacteria from natural waters. Appl. Microbiol.
£9, 186-194.
Blickstad, E., Enfors, S-0 and Molin, G. (1981). Effect of
hyperbaric CO^ pressure on the microbial flora of pork o ostored at 4 C or 14 C. J. appl. Bact.50, 493-504.
Bogadi, H.O. and Sewell, M.M.H. (1974). A study of the route
of dissemination of orally administered spores of Clostridium
novyi type B in guinea-pigs and sheep. Res. in Vet. Sci.
17, 179-181.
Borchert, L.L., Greaser, M.L., Bard, J.C., Cassens, R.G. and
Briskey, E.J. (1967). Electron microscopy of a meat
emulsion. J. Fd. Sci. 32, 419.

341
Breed, R.S. Murray, E.G.D. and Smith, N.R. (1948). Bergey's
Manual of Determinative Bacteriology (5th edition).
Williams and Wilkins Co. Baltimore, U.S.A.
Breed, R.S., Murray, E.G.D. and Smith, N.R. (1957). In 'Bergey's
Manual of Determinative Bacteriology' 7th edition. Williams
and Wilkins Co. Baltimore U.S.A.
Brooks, F.T. and Hansford, C.G. (1923). Mould growth upon cold-
store meat. Trans, Brit, mycol. Soc. 8, 113.
Brown, M.H. (1977). The microbiology of the British fresh sausage
PhD Thesis, University of Bath, England.
Brown, A.D. and Weidemann, J.F. (1958). The taxonomy of psychro-
philic meat-spoilage bacteria; a reassessment. J. appl.
Bact. 21, 11.
Brownlie, L.E. (1966). Effect of some environmental factors on
psychrophilic microbacteria. J. appl. Bact. 29, 447-454.
Bryan, F.L. (1979). In "Food-borne infections and intoxications",
2nd edition, (ed. H.Riemann and F.L. Bryan) Academic Press,
New York, U.S.A.
Bryan, F.L. (1980). Food-borne diseases in the United States
associated with meat and poultry. J. Food Prot. 43, 140.
Buchanan, R.E. and Gibbons, N.E. (1974). (eds). In "Bergey's
Manual of Determinative Bacteriology" 8 th edition.
Williams and Wilkins Co., Baltimore, U.S.A.
Buchanan, R.E., St. John-Brooks, R. and Breed, R.S. (1948).
International bacteriological code of nomenclature.
J. Bact. 55, 287-306.
Buckley, J., Morrissey, P.A. and Daly, M. (1976). Aseptic technique
for obtaining sterile beef tissue. J. Fd. Technol. 11,
427-430.

342
Burroughs, L.F. and Sparks, A.H. (1963). The determination of
total SO- in cider. Analyst 88, 304.
Burroughs, L.F. and Sparks, A.H. (1964). The determination of free
SO- in cider. Analyst 89, 55.2
Burton, H.S., McWeeny, D.J. and Biltcliffe, D.O. (1963). Non-
enzymic browning; the role of unsaturated carbonyl compounds
as intermediates and of SO^ as an inhibitor of browning.
J. Sci. Food Agric. 14, 911-920.
Butler, J.L., Stewart, J.C., Vanderzant, C ., Carpenter, Z.L.
and Smith, G.C. (1979). Attachment of micro-organisms to
pork skin and surfaces of beef and lamb carcasses. J. Food
Prot. 42, 401-406.
Butler, J.L., Vanderzant, C ., Carpenter, Z.L., Smith, G.C.,
Lewis, R.E., and Dutson, T.R. (1980). Influence of certain
processing steps on attachment of micro-organisms to pork
skin. J. Food Prot. 43, 699-705.
Cary, W.E. (1916). The bacteriological examination of sausages
and its sanitary significance. Am. J. publ. Hlth. 6, 124.
Cecil, R. and McPhee, J.R. (1954). The estimation of thiols and
disulphides by potentiometric titration with silver nitrate.
Biochem. J. 59, 234-240.
Cesari, E.P. (1919). La maturation des saucissons. Compt. rend.
Acad, de Sci. 168, 802.
Cesari, E.P. and Guilliermond, A. (1920). Les levures des saucissons.
Ann. de 1'Institut Pasteur 34, 229.
Chau, P.Y., Shortridge, K.F. and Huang, C.T. (1977). Salmonella
in pig carcasses for human consumption in Hong Kong: a study
on the mode of contamination. J. Hyg. 78, 253-260.

343
Cherry, W.B., Scherago, M. and Weaver, R.H. (1943). The occurrence
of Salmonella in retail meat products. Am. J. Hyg. 37,
211-215.
Childers, A.B., Keahey, E.E. and Vincent, P.G. (1973). Sources of
salmonellae contamination of meat slaughtering procedures.
II. J. Milk Fd.Technol. 635-638.
Chipault, J.R., Mizuno, G.R.,Hawkins, J.M. and Lundberg, W.O.
(1952). The antioxidant properties of natural spices. Fd.
Res. 17, 46.
Chipault, J.R., Mizuno, G.R. and Lundberg, W.O. (1956). The
antioxidant properties of spices in foods. Fd. Technol.
10, 209.
Christensen, W.B. (1946). Urea decomposition as a means of
differentiating Proteus and paracolon cultures from each
other and from Salmonella and Shigella. J. Bact. 52, 461.
Christian, J.H.B. (1963). The preservation of minced beef with
sulphur dioxide. CSIRO Food Pres. Quart. 23, 30-36.
Clarke, H.T. (1932). The action of sulfite upon cystine. J. Biol.
Chem. 97, 235-248.
Clise, J.D. and Swecker, E.E. (1965). Salmonellae from animal by
products. Publ. Hlth.Rep. Wash. 80, 899.
Cochran, W.G. (1950). Estimation of bacterial densities by means
of the 'most probable number'. Biometrics 5, 105-116.
Collins, C.H. and Lyne, P.M. (1976). In "Microbiological Methods"
4th edition, Butterworths, London.
Collins-Thompson, D.L., Sorhaug, T., Witter, L.D. and Ordal, Z.J.
(1972). Taxonomic consideration of Microbacterium lacticum.
Microbacterium flavum and Microbacterium thermosphactum.
Int. J. Syst. Bacterid. 22, 65-72.

344
Cox, N.A., Mercuri, A.J., Carson, M.O., and Tanner, D.A. (1979).
Comparative study of Micro-ID, Minitek and conventional
methods with Enterobacteriaceae freshly isolated from foods.
J. Food Prot. 735-738.
Craven, J.A. and Hurst, D.B. (1982).The effect of time in lairage
on the frequency of salmonella infection in slaughtered pigs.
J. Hyg. Camb. SS, 107-111.
Dainty, R.H. and Hibbard, C.M. (1980). Aerobic metabolism of
Brochothrix thermosphacta growing on meat surfaces and in
laboratory media. J. appl. Bact. 45, 387-396.
Dainty, R.H. and Hibbard, C.M. (1983). Precursors of the major end
products of aerobic metabolism of Brochothrix thermosphacta.
J. appl. Bact. 55, 127-133.
Davenport, R.R. (1980). In "Biology and activities of yeasts"
(eds. Skinner, F.A., Passmore, S.M. and Davenport, R.R.)
Academic Press, London, 1-27, 215-230, 261-278.
Davidson, C.M. (1970). Some aspects of the microbiology of pre
packed beef. PhD Thesis, University of Bath, England.
Davidson, C.M. and Hartree, E.F. (1968). Cytochrome as a guide
to classifying bacteria: taxonomy of Microbacterium thermo
sphactum. Nature, Lond. 210, 502-504.
Davidson, C.M., Mobbs, P. and Stubbs, J.M. (1968). Some
morphological and physiological properties of Microbacterium
thermosphactum. J. appl. Bact. 31, 551-559.
Davidson, C.M., Dowdell, M.J. and Board, R.G. (1973). Properties
of Gram-negative aerobes isolated fron meats. J. Food Sci.
303-305.
Davies, G.H.G., Fomin, L., Wilson, E. and Newton, K.G. (1969).
Numerical taxonomy of Listeria, streptococci and possibly

345
related bacteria. J. gen. Microbiol. 333-348.
Dawkins, H.C. and Robertson, J. (1967). Salmonellas in animal feeding
stuffs. Mon. Bull. Minist. Hlth. 26, 215.
Deibel, R.H. and Evans, J.B. (1960). Modified benzidine test for
the detection of cytochrome containing respiratory systems in
micro-organisms. J. Bact. 79, 356-360.
den Dooren de Jong, L.E. (1926). Bijdrage tot de kennis van het
mineralisatieproces. Nijgh and Van Ditmar, Rotterdam cited
by Stanier et al. (1966).
de Man, J.C. (1977). MPN tables for more than one test. Europ. J.
appl. Microbiol. 4, 307-316.
de Man, J.C. (1983). Corrected tables for the most probable number.
Europ. J. appl. Microbiol. Biotech. ^ , 301-305.
de Man, J.C., Rogosa, M. and Sharpe, M.E. (1960). A medium for
the cultivation of lactobacilli. J. appl. Bact. 23, 130.
Dennis, C. (1978). Post-harvest spoilage of strawberries. A.R.C.
Res. Rev. 4, 38-42.
Dennis, C. and Bugahiar, R.W.M.(1980). Yeast spoilage of fresh and
processed fruits and vegetables. In "Biology and activities
of yeasts" (eds. Skinner, F.A., Passmore, S.M., Davenport,
R.R.). Academic Press, London, 123-133.
Dennis, C. and Harris, J.E. (1979). The involvement of fungi in
the breakdown of sulphited strawberries. J. Sci. Food
Agric. 30, 687-691.
Dijkmann, K.E., Koopmans, M. and Mossel, D.A.A. (1980). The
recovery and identification of psychrotrophic yeasts from
chilled and frozen, comminuted fresh meats. In "Biology and
activities of yeasts" (eds. Skinner, F.A., Passmore, S.M.,
Davenport, R.R.) Academic Press, London, p. 300.

346.
Dockerty, T.R., Ockerman, H.W., Cahill, V.R., Kunkle, L.E. and
Weiser, H.H. (1970). Microbial level of pork skin as affected
by the dressing process. J. Anim. Sci. 30, 884-890.
Douglas, J. (1966). The behaviour of Saccharomyces cerevisiae var. ellip-
soides in solutions containing sulphurous acid. PhD Thesis
University of London.
Dowdell, M.J. and Board, R.G. (1967). Changes in microflora
during storage of sausages. J. appl. Bact. 30 (3) ii.
Dowdell, M.J. and Board, R.G. (1968). A microbiological survey
of British fresh sausage. J. appl. Bact. 31, 378-396.
Dowdell, M.J. and Board, R.G. (1971). The microbial associations
in British fresh sausages. J. appl. Bact. 317-337.
Drake, S.D., Evans, J.B. and Niven, C.F. (1958). Microbial flora
of packaged frankfurters and their radiation resistance.
Fd. Res. 23, 291-296.(VA-Drake, S.D., Evans, J.B. and Niven, C.F. (1959). The identity
of yeasts in the surface flora of packaged frankfurters.
Fd. Res. 24, 243-246.
Duitschaever, C.L. and Buteau, C. (1979). A research note -
Incidence of Salmonella in pork and poultry products. J.
Food Prot. 42, 662-663.
Dupuy, P. (1959). L'inhibition par I'acide sulfureux de l'oxydation
de 1'ethanol par Acetobacter rancens. Ann. Technologie
agricole S, 233 and 337.
Dutka, B.J. (1973). Coliforms are an inadequate index of water
quality. J. Environ. Health 36, 39-46.
Dutka, B.J. and Kwan, K.K. (1978). Health indicator bacteria in
water-surface microlayers. Can. J. Microbiol. 24, 187-188.

347
Dyett, E.J. and Shelley, D. (1952). Microbiology of British fresh
sausage. 1st Int. Cong. Food Sci. and Tech. London p. 54.
Dyett, E.J. and Shelley, D. (1955). Microbiology of the British
fresh sausage. Proc. 1st Int. Congr. Fd. Sci. & Technol.
2 , 393.
Dyett, E.J. and Shelley, D. (1966) . Effects of sulphite preservation
in British fresh sausage. J. appl. Bact. 29, 439.
Eddy, B.P. and Kitchell, A.G. (1959). Cold-tolerant fermentative
Gram negative organisms from meat and other sources. J. appl.
Bact. 22, 57.
Edel, W. and Kampelmacher, E.H. (1969). Salmonella isolations in
9 European laboratories using a standardised technique .
Bull. W.H.O. 41, 297-306.
Edel, W. and Kampelmacher, E.H. (1970). Salmonella in mesenteric
and portal lymph nodes and faeces from normal slaughter pigs.
Zentrablatt fur Veterinmedizin Reihe B 17, 875-879.
Edel, W., van Schothorst, M., Guinee, P.A.M. and Kampelmacher
E.H. (1973). Mechanism and prevention of Salmonella infections
in animals. In "Microbiological Safety of Food" (eds. Hobbs,
B.C., Christian, J.H.B.), Academic Press, London, pp. 247-256.
Edwards, P.R. and Ewing, W.H. (1972). In "Identification of the
Enterobacteriaceae" 3rd edition. Burgess, Minneapolis, U.S.A.
Eenick, W.H. (1963). Consequence of a serious Salmonella contamin
ation in the meat preserving industry for the state and
municipality. Tijdschr. Diergeneesk. 88, 501.
Eichholz, W. (1902). Erdbeerbacillus (Bacterium fragi). Zentralblatt
fur Bakteriologie, Parasitehkunde, Infektionskrankheiten und
Hygiene Abteilung II, 9, 425-428.

348
Elliott, J.M. (1977). Some methods for statistical analysis of
samples of bethnic invertebrates. Freshwater Biol. Assoc.
Publication No. 25, Ambleside, England.
Empey, W.A. and Scott, W.J. (1939). Investigations of chilled beef.
I. Microbial contamination acquired in the meat works. Bull.
Coun. Sci. Ind. Res. Australia, 126.
Empey, W.A. and Vickery, J.R. (1933). The use of carbon dioxide
in the storage of chilled meat. J. Coun. Sci. Industr.
Res. Aust 6, 233.
Enfors, S.-O., Molin, G. and Ternstrdm, A. (1979). Effect of
packaging under carbon dioxide, nitrogen or air on the
microbial flora of pork stored at 4^C. J. appl. Bact. 47,
197-208.
Ewing, W.H., Farmer III, J.J., and Brenner, D.J. (1980). Proposal
of Enterobacteriaceae fam. nov. nom. rev. to replace
Enterobacteriaceae Rahn 1937, nom. fam. cons. (Opin 15,
Jud. Comm. 1958) which lost standing in nomenclature on
1 January 1980. Int. J. System Bacteriol. 30, 674-675.
Facklam, R.R. (1972). Recognition of group D streptococcal
species of human origin by biochemical and physiological
tests. Appl. Microbiol. 23, 1131-1139.
Faparusi, S.I. (1969). Effect of pH on the preservation of palm
wine by sulphite. Appl. Microbiol. 18,122-123.
Farmer III, J.J., Brenner, D.J. and Ewing, W.H. (1980). Opposition
to recent proposals which would reject the family name
Enterobacteriaceae and Escherichia as its type genus. Int.
J. System. Bacteriol. 660-673.
Farnsteiner, K.Z. (1904). Uber organisch gebundene schweflige
sMure in nahrungsmitteln. Z. Untersuch. Lebensm. 7, 449.

349
Felsenfeld, O., Young, V.M. and Yoshimura, T. (1950). A survey
of Salmonella organisms in market meats, eggs and milk.
J. Amer. vet. Med. Assoc. 116, 17.
Floyd, T.M., Baranski, J.R. and El-gannani, M. (1953). Recovery
of human enteric pathogens on meat from butchers shops in
Cairo, Egypt. J. infect, dis. 92, 224-227.
Fornachon, J.C.M. (1963). Inhibition of certain lactic acid
bacteria by free and bound sulphur dioxide. J. Sci. Food
Agric. 857-862.
Fournaud, J., Jacquet, B. and Poujardieu, B. (1971). Development
of microflora of sausage meat depending in preservatives
used and method of storage. Ind. Alimen. & Agric. 88,
15-26.
Frazier, W.C. (1926). A method for the detection of changes in
gelatin due to bacteria. J. Infect. Dis. 39, 302-309.
Freeman, G.G. and Donald, G.M.S. (1957). Fermentation process
leading to glycerol. I. The influence of certain variables
on glycerol formation in the presence of sulfites. Appl.
Microbiol. 5, 197-210.
Freese, E. , Sheu, C.W., Galliers, E. (1973). Function of lipophilic
acids as antimicrobial food additives. Nature, Lond. 241,
321-325.
Fridovitch, I. and Handler, P. (1961). Detection of free radicals
generated during enzymic oxidations by the initiation of
sulfite oxidation. J. biol. Chem. 236, 1836-1840.
Galbraith, N.S., Archer, J.F. and Tee, G.H. (1961). Salmonella
saint-paul infection in England and Wales in 1959. J. Hyg.
Camb. 59, 133.

350,
Galton, M.M., Lowery, W.P. and Hardy, A.V. (1954). Salmonella
in fresh and smoked pork sausage. J. infect. Dis. 95, 232.
Gancedo, C ., Gancedo, J.M. and Sols, A. (1968). Glycerol metabolism
in yeasts. Pathways of utilization and production. Europ.
J. Biochem 165-172.
Gardner, G.A. (1965). The aerobic flora of stored meat with particular
reference to the use of selective media. J. appl. Bact. 28,
252-264.
Gardner, G.A. (1966). A selective medium for the enumeration of
Microbacterium thermosphactum in meat and meat products. J.
appl. Bact. 29, 455-460.
Gardner, G.A. (1968). Effects of pasteurization or added sulphite
on the microbiology of stored vacuum packed baconburgers.
J. appl. Bact. 31, 462-478.
Gardner, G.A.(1980). Occurrence and significance of Pseudomonas
in Wiltshire Bacon Brines. J. appl. Bact. ^ , 69-74.
Gardner, G.A. (1981). Brochothrix thermosphacta (Microbacterium
thermosphactum) in spoilage of meats: A review. In "Psychro
trophic micro-organisms in spoilage and pathogenicity"
(eds. Roberts, T.A., Hobbs, G., Christian, J.H.B. and
Skovgaard, N.), Academic Press, London, p. 139-173.
Gardner, G.A. and Carson, A.W. (1967). Relationship between
carbon dioxide production and growth of pure strains of
bacteria on porcine muscle. J. appl. Bact. 30, 500.
Gardner, G.A. and Patton, J. (1969). The importance of Microbacterium
thermosphactum in the microbiology of meats. Proc. 15th
Europ. Meeting Meat Res. Workers, Helsinki, 176-184.
Gardner, G.A., Carson, A.W., and Patton, J. (1967). Bacteriology

351,
of prepacked pork with reference to the gas composition
within the pack. J. appl. Bact. 30, 321-333.
Garvie, E.I. (1967). The production of L(+) and D(-) lactic acid
in culture of some lactic acid bacteria with a special study
of Lactobacillus acidophilus NCD02. J. Dairy Res. 31-38.
Gehman, H. and Osman, E.M. (1954). The chemistry of the sugar-sulf-
ite reaction and its relationship to food problems. Adv. Food
Res. 5, 53-96.
Gerats, G.E., Snijders, J.M.A. and van Logtestijn, J.G. (1981).
Slaughter techniques and bacterial contamination of pig
carcasses. Proc. Europ. Meat Workers Assocn., Vienna (B4),
198-200.
Gerrard, F. (1976). "Sausage and small goods production" Northwood
Publications, London.
Gibson, T. and Abd-el-Malek, Y. (1945). The formation of CO by2
lactic acid bacteria and Bacillus licheniformis and a cultural
method of detecting the process. J. Dairy Res. 14, 35.
Gill, C.O. (1979). Intrinsic bacteria in meat. J. appl. Bact.
47, 367-378.
Gill, C.O. (1980). Total and intramuscular bacterial populations
of carcasses and cuts. Am. Meat Sci. Assoc. 33rd Reciprocal
Meat Conf. 47-53.
Gill, C.O. and Newton, K.G. (1977). The development of the aerobic
spoilage flora on meat stored at chill temperatures. J. appl.
Bact. 43, 189.
Gill, C.O. and Penney, N. (1977). Penetration of bacteria into
meat. Appl. & Environ. Microbiol 33, 1284-1286.
Gill, C.O., Penney, N. and Nottingham, P.M. (1976). Effect of

352.
delayed evisceration on the microbial quality of meat. Appl.
& Environ. Microbiol. 31, 465-458.
Gill, C.O., Penney, N., and Nottingham, P.M. (1978). Tissue sterility
in uneviscerated carcasses. Appl. & Environ. Microbiol. 36,
356-359.
Gill, O.N., Bartlett, C.L.R., Sockett, P.N., Vaile, M.S.B., Rowe,
B., Gilbert, R.J., Dulake, C., Murrell, H.C. and Salmaso, S.
(1983). Outbreak of Salmonella napoli infection caused by
contaminated chocolate bars. The Lancet i, 574-577.
Gillespie, N.C. (1981). A numerical taxonomic study of Pseudomonas
like bacteria isolated from fish in S . Eastern Queensland
and their association with spoilage. J. appl. Bact. 50,
29-44.
Gillespie, N.C. and Macrae, I.C. (1975). The bacterial flora of
some Queensland fish and its ability to cause spoilage.
J. appl. Bact. 39, 91-100.
Glage, F. (1901) . "Uber die Bedentung der Aromobacterien fiir
die Fleischhygiene". Z. Fleisch-und Milchhyg. 11, 131.
Goodfellow, M. and Trüper, H.G. (1982). Escherichiaceae
nom. nov., a name to replace Enterobacteriaceae. Request
for an opinion. Int. J. System. Bacteriol. 32, 383.
Gottschalk, G. (1979). In "Bacterial Metabolism" Springer-Verlag,
Berlin.
Gould, G.W., Brown, M.H. and Fletcher, B.C. (1983). Mechanisms
of action of food preservation procedures. In "Food
Microbiology, advances and prospects" Society for Applied
Bacteriology Symposium Series no. 11. Academic Press, London.
Grassetti, D.R. and Murray, J.F. Jr. (1967). Determination of
sulfhydryl groups with 2,2' or 4,4' dithiopyridine. Arch.

353
Biochem. Biophys. 119, 41-49.
Grau, F.H. (1980). Inhibition of the anaerobic growth of
Brochotrhix thermosphacta by lactic acid. Appl. Environ.
Microbiol. 40, 433-435.
Gray, P.A. and Stewart, D.J. (1980). Numerical taxonomy of some
marine Pseudomonads and Alteromonads. J. appl. Bact. 49,
375-383.
Greer, F.E. and Nyhan, F.V. (1928). The sanitary significance
of lactose-fermenting organisms not belonging to the B. coli
group III. Bacterial associations in cultures containing
lactose fermenting bacteria. J. infect. Dis. 525-536.
Grüber , T. (1902). Pseudomonas fragaride. Eine Erdbeergeruch
erzeugende Bakterie. Zentralblatt fiir Bakteriologie Abt.
2, 705-712.
Gyllenberg, H.G., Eklund, E., Antila, M. and Vartiovaara, U.
(1963). Contamination and deterioration of market milk
IV. Taxometric classification of pseudomonads. Acta Agr.
Scand 13, 157-176.
Haddock, R.L. (1970). Efficacy of examining rectal swabs to
detect swine Salmonella-carriers. Am. J. Vet. Res. 31,
1510.
Haines, R.B. (1933). The bacterial flora developing on stored
lean meat, especially with regard to "slimy" meat.
J. Hyg. Camb. 33, 175.
Haines, R.B. (1937). Microbiology in the preservation of animal
tissues. Spec. Rep. Fd. Invest. Bd., Lond. No. 45.
Haines, R.B. and Scott, W.J. (1940). An anaerobic organism
associated with bone taint in beef. J. Hyg. Camb. 40,
154-161.

3 54 .
Haines, R.B. and Smith, E.C. (1933). The storage of meat in
small refrigerators
Spec. Rep. Fd. Invest. Bd., Lond., No. 43.
Hammond, S.M. and Carr, J.G. (1976). The antimicrobial activity
of sulphur dioxide - with particular reference to fermented
and non-fermented fruit juices. In "Inhibition and in
activation of vegetative microbes (eds. Skinner, F.A.,
and Hugo, W.B.). Academic Press, London, 86-110.
Hag, A., Webb, N.B., Whitfield, J.K. and Ivey, F.J. (1973).
Effect of composition on the stability of sausage-type
emulsion. J. Fd. Sci. 271.
Harvey, R.W.S. and Price, T.H. (1967). The isolation of
salmonellas from animal feeding stuffs. J. Hyg. Camb.
65, 237-244.
Harvey, R.W.S., Price, T.H. and Morgan, J. (1977). Salmonella
surveillance with reference to pigs - Cardiff abattoir,
1968-1975. J. Hyg. Camb. 78, 439-448.
Hasegawa, T., Pearson, A.M., Price, J.F. and Lechowich, R.V. (1970).
Action of bacterial growth on the sarcoplasmic and urea-
soluble proteins from muscle. Appl. Microbiol. 20, 117-122.
Hibbert, H.R. (1970). White spot in sausages. B.F.M.I.R.A. Res.
Rept. No. 159.
Hitchener, B.J., Egan, A.F. and Rogers, P.J. (1979). Energetics
of Microbacterium thermosphactum in glucose-limited continuous
culture. Appl. Environ. Microbiol. 37, 1047-1052.
Hockley, S.J.E. (1980). A survey of the microbiology of the British
fresh skinless sausage. B.Sc. Project Report No. 443,
University of Bath, England.

355,
Hormaeche, E. and Salsamendi, R. (1936). Sobre la presencia de
Salraonelas en los ganglios mesentericos de cerdos normales
Arch. Urug. Med. Cir. y. Espec. 9, 665.
Hormaeche, E. and Salsamendi, R. (1939). El cerdo normal como
"portador" de Salmonelas. Arch. Urug. Med. Cir. y Espec.
375-387.
Hubener, cited by Cary, W.E. (1916). Am. J. publ. Hlth. 6, 124.
Hugh, R. and Leifson, E. (1953). The taxonomic significance of
fermentative vs.oxidative metabolism of carbohydrates by
various Gram-negative bacteria. J. Bact. 24-26.
Humphrey, R.E., Ward, M.H. and Hinze, W.L. (1970). Spectrophoto-
metric determination of sulfite with 4,4'dithiodipyridine
and 5,5'-dithiobis-(2-nitrobenzoic acid). Analyt.Chem. 42,
698-702.
Hurst, J. (1972). The selection of the microbial association of
British fresh sausage. M.Sc. thesis. University of Bath, England.
Hussong, D., Damare, J.M., Weiner, R.M. and Colwell, R.R. (1981).
Bacteria associated with false-positive most-probable-number
coliform test results for shellfish and estuaries. Appl.
& Environ. Microbiol. 41, 35-45.
Hussong, R.V., Long, H.F. and Hammer, B.W. (1937). Classification
of the organisms important in dairy products Agr. Exp.
Stat., Iowa State College Research Bulletin 225. 117-136.
Ingram, M. (1934). The microbiology of pork and bacon Rep.
Fd. Invest. Bd., London, No. 77.
Ingram, M. (1948). Germicidal effects of free and combined
sulphur dioxide. J. Soc. Chem. Ind., Lond. 67, 18-21.
Ingram, M. (1949). Science in the imported meat industry. Part II,
Hygiene and Storage. J. Royal Sanit. Inst it. 69, 39-47.

356,
Ingram, M. (1958). The yeasts in food spoilage. In "Chemistry
and biology of yeasts" (ed. A.H. Cook). Academic Press,
London, 603-633.
Ingram, M. (1962). Microbiological principles in prepacking meats.
J. appl. Bact. 25, 259.
Ingram, M. and Dainty, R.H. (1971). Changes caused by microbes in
spoilage of meats. J. appl. Bact. 21-39.
Ingram, M . and Hobbs, B.C. (1954). The bacteriology of pasteurized
canned hams . J.R. sanit. Inst. 74, 1151.
Ingram, M., Ottaway, F.J.H. and Oppock, J.B.M. (1956). The preserv
ative action of acid substances in food. Cherny Ind. 1154-1163.
Ingram, M. and Vas, K. (1950a). Combination of sulphur dioxide
with concentrated orange juice I. Equilibration states. J.
Food Sci. Agric. 1, 21-27.
Ingram, M. and Vas, K. (1950b). Combination of sulphur dioxide with
concentrated orange juice. II. The rate of the reaction. J.Sci.
Food Agric. 1, 63-66.
Jacobs, J., Guinee, P.A.M., Kampelmacher, E.H. and van Keulen, A.
(1963). Studies on the incidence of Salmonella in imported
fish meal. Zentralblatt fiir Veterinarmedizin Reihe B 10,
542-550.
Jensen, L.B. (1944). Microbiological problems in the preservation
of meats Bact. Rev. 8, 161.
Jensen, L.B. and Hess, W.R. (1941). A study of ham souring. Fd.
Res. 6, 273-333.
Jepsen, A. (1947). Bacteriology of pork. Microbial contamination
acquired in the meat works. Royal Veterinary and Agricultural
College, Copenhagen Yearbook, 40.

35 7 .
Johnson, R., Colwell, R.R., Sakazaki, R. and Tamura, K. (1975).
Numerical taxonomy study of the Enterobacteriaceae. Int. J.
Syst. Bacteriol. 25, 12-37.
Johnston, R.W., Green, S.S., Chiu, J., Pratt, M. and Rivera, J.
(1982). Incidence of Salmonella in fresh pork sausage in
1979 compared with 1969. J. Fd. Sci. 47, 1369-1371.
Jones, B.H., Farkas, G., Ghosh, A. and Hobbs, B.C. (1964). Salmonella
hrandehburg: an epidemiological study. Mon. Bull. Minist.
Hlth. 23, 162.
Jones, D. (1975). A numerical taxonomic study of coryneform and
related bacteria. J._ gen. Microbiol. 87, 52-96.
Joslyn, M.A. and Braverman, J.B.S. (1954). The chemistry and
technology of the pretreatment and preservation of fruit and
vegetable products with sulphur dioxide and sulfites. Adv.
Fd. Res. 5, 97-160.
Juffs, H.S. (1973). Identification of Pseudomonas spp. isolated
from milk produced in south-eastern Queensland. J. appl.
Bact. 36, 585-598.
Kampelmacher, E.H., Guinee, P.A.M., Hofstra, K. and Van Keulen, A.
(1961). Studies on Salmonella in slaughter houses. Zentral
blatt fUr VeterinSrmedizin 10, 1025-1042.
Kampelmacher, E.H., Guinee, P.A.M. and Van Keulen, A. (1965).
Investigations on the occurrence of Salmonella in pigs fattened
with decontaminated and normal meal. Tijschr. Diergeneesk 90,
1270.
Kampelmacher, E.H. , Guiné’e, P.A.M., Hofstra, K. and Van Keulen, A.
(1963). Further studies on Salmonella in slaughter pigs.
Zbl. vet. Med. Serv. B., 10, 1.

358
Karmas, E. (1977). "Sausage products technology" Noyes Data
Corp., Park Ridge, New Jersey, U.S.A.
Keddie, R.M. (1951). The enumeration of lactobacilli on grass and
silage. Proc. Soc. appl. Bact. 157.
Kelly, D. and Dainty, R.H. (1983) . Biosynthesis of glutamate via
enzymes of the T.C.A. cycle in Brochothrix thermosphacta .
J. gen. Microbiol, in press.
Kilsby, D.C. and Pugh, M.E. (1981). The relevance of the distribution
of micro-organisms within batches of food to the control of
microbiological hazards from foods. J. appl. Bact. d , 345-354.
King, A.D.Jr., Ponting, J.D., Fanshuck, D.W., Jackson, R. and Mihara,
K. (1981). Factors affecting death of yeasts by sulphur dioxide.
J. Fd. Prot. 44, 92-97.
King, E.O., Ward, M.K. and Raney, D.E. (1954). Two simple media
for the demonstration of pyocyanin and fluorescin. Journal
of Labor^atory and Clinical Medicine 44, 301-307.
Kirsch, R.H., Berry, F.E., Baldwin, G.L. and Foster, E.M. (1952).
The bacteriology of refrigerated ground beef. Fd. Res. 17,
495.
Kitchell, A.G. and Ingram, M. (1956). A comparison of bacterial
growth on fresh meat and on frozen meat after thawing. Annales
Institut Pasteur, Lille 8, 121.
Kluyver, A.J. and van Niel, C.B. (1936). Prospects for a natural
system of classification of bacteria. Zbl. Bakt. II. Abt.
94, 369-403.
Kneteman, A. (1947). A reaction on the formation of hydrogen
peroxide by micro-organisms in solid media. Antonie v.
Leeuwenhoek 13, 55.

359
Komma, F. (1910). cited by Cary (1916). Zentralblatt fiir
Bakteriologie, Abt. I 55, 1-14.
Kopeloff, N. and Beerman, P. (1922). Modified Gram stains. J.
infect. Dis. 31, 480.
Koser, S.A. and Shinn, W.C. (1927). Aerobic spore-forming bacilli
which ferment lactose. J. Am. Water Works Assoc. 18, 328-336.
Kovacs, N. (1956). Identification of Pseudomonas pyocyanea by the
oxidase reaction. Nature, Lond. 178, 703.
Krol, B. and Moerman, P.C. (1959/60). Effect of sulphite on the
colour and bacteriological condition of mince. (Translation
only). Conserva 8, 1-11.
Lafon la fourcade, S. and Peynaud, E. (1974). Sur l'action anti-
bacterienne de l'anhydride sulfureux sous forme libre et
sous forme combinée. Connaiss. Vigne Vin. 74, 187-203.
Lafontaine, A., Goblet, J. and De Mayer-Cleempoel, S. (1955).
Action bactériologique des sulfites sur les viandes. Arch.
Belges Méd Soc. 13, 288-292.
Lapage, S.P. (1979). Proposai of Enterobacteriaceae nom. nov. as
a substitute for the illegitimate but conserved name
Enterobacteriaceae Rahn 1937 Jlequest for an opinion. Int.
J. System. Bacteriol. 29, 265-266.
Law, B.A., Cousins, C.M., Sharpe, M.E. and Davies, F.L. (1979).
Psychrotrophs and their effects on milk and dairy products.
In "Cold tolerant microbes in spoilage and the environment"
(eds. Russell, A.D., Fuller, R.) Academic Press, London, 137-152.
Lawrie, R.A. (1974). In "Meat Science" 2nd edition, Pergamon Press,
Oxford.
Lawrie, R.A. (1978). In "Meat Science" 3rd edition, Pergamon Press,
London.

360,
Lea, C.H. (1931a). Chemical changes in the fat of frozen and chilled
meat. Part I: frozen mutton and lamb. J. Soc, Chem. Ind. Lond.
50, 207T-213T.
Lea, C.H. (19312?) . Chemical changes in the fat of frozen and chilled
meat Part II: chilled beef. J. Soc. Chem. Ind. Lond. 50,
215T-220T.
Leads, C. (1979). The organoleptic stability of British fresh sausage.
PhD Thesis, University of Bath, England.
Leaton, J. (1968). Studies on Microbacterium thermosphactum, a spoilage
bacterium in meat and meat products. B.Sc. dissertation,
Nottingham University.
Lechowich, R.F. (1971). Microbiology of meat. In "Science of
meat and meat products" 2nd edition (eds. Price, J.F.,
Schweigert, B.S.). Freeman and Co., San Francisco.
Lee, W.H., Harris, M.E., McClain, D., Smith, R.E. and Johnston, R.W.
(1980). Two modified selenite media for the recovery of
Yersinia enterocolitica from meats. Appl. & Environ. Microbiol.
205-209.
Legan, J.D. (1981). Microbial growth and sulphite loss in skinless
pork sausage. B.Sc. Project Report No. 492 University of Bath.
Lehmann, W. (1964). Vorkommen und lokalisation von Salmonellen
in fleischverarbeitenden Betrieben. Monatshefte fiir Veterin'dr-
medizin 19, 744.
Leitch, G.B. (1925). Disturbing factors in the presumptive test
for B. coli. J. Am. Water Works Assoc. 13, 186-192.
Lepovetsky, B.C., Weiser, H.H. and Deatherage, F.E. (1953). A
microbiological study of lymph nodes,bone marrow and muscle
tissue obtained fron slaughtered cattle. Appl. Microbiol. 1,
57-59.

351
Levine, N.D., Peterson, E.H. and Graham, R. (1945). Studies on
swine enteritis. Am. J. Vet. Res. 4, 242-247.
Lo, C.B., Huang, C.T. and Chan, D.S.S. (1967). Salmonella serotypes
isolated from pigs slaughtered in an abattoir in Hong Kong.
Tropical Medicine 9, 71-78.
Lodder, J. (1970). In "The Yeasts - a taxonomic study" 2nd edition.
North-Holland, Amsterdam.
London, J. (1976). The ecology and taxonomic status of the
lactobacilli. Ann. Rev. Microbiol. 30, 279-301.
Luck , E. (1977). Effect of food preservatives on micro-organisms.
In "Chemische Lebensmittelkonservierung-Stoffe, Wirkungen,
Methoden" Springer-Verlag, Berlin.
Lund, B.M. (1968). A bacteriological study of stored, sulphite
treated peeled potatoes. J. appl. Bact. 31, 479-492.
Mackey, B.M. and Derrick, C.M. (1979). Contamination of deep
tissues of carcasses by bacteria present on slaughter
instruments or in the gut. J. appl. Bact. 46, 355-366.
Maoris, B. (1972). Transport and toxicity of sulfur dioxide in
the yeast Saccharomyces cerevisiae var. ellipsoideus.
PhD Thesis, University of Michigan, U.S.A.
Maoris, B.J. and Markakis, P. (1974) . Transport and toxicity of
sulfur dioxide in Saccharomyces cerevisiae var .ellipsoides.
J. Sci. Food Agric. 25, 21-29.
Martin, T.A. and Patterson, J.T. (1982). Some characteristics of
pseudomonads from fresh meat. J. appl. Bact. 53, xii.
Masurovsky, E.B., Goldblith, S.A. and Voss, J. (1963). Differential
medium for selection and enumeration of members of the genus
Pseudomonas. J. Bact. 85, 722.

362
Mayfield, C.I. and Inniss, W.E. (1977). A rapid, simple method
for staining bacterial flagella. Can. J. Microbiol.
23, 1311-1313.
Mead, G.C. and Adams, B.W. (1977). A selective medium for the
rapid isolation of pseudomonads associated with poultry
spoilage. Brit. Poult. Sci. 661-670.
Mercuri, A.J. and Cox, N.A. (1979). Coliforms and Enterobacteriaceae
isolates from selected foods. J. Fd. Prot. 42, 712-714.
Meyer, E.M. (1918). An aerobic spore-forming bacillus giving gas
in lactose broth isolated in routine water examination. J.
Bact. 3, 9-14.
Meyerhof, O., Ohlmeyer, P. and Mohle, W. (1938). Coupling between
oxidation-reduction and phosphorylation in the anaerobic
carbohydrate splitting. II. Coupling as an equilibrium
reaction. Biochem. Z. 297, 113.
Michels, P.A.M., Michels, J.P.J., Boonstra, J. and Konings, W.N.
(1979) . Generation of an electrochemical proton gradient in
bacteria by the excretion of metabolic end products. F.E.M.S.
Microbiol. Lett. 5, 357-364.
Migula, (1894). Cited by Murray, E.G.D., Smith, N.R. and
Breed, R.S. (eds) in "Bergey's Manual of determinative
bacteriology" 7th edition 1957, Williams and Wilkins,
Baltimore, U.S.A.
Miles, A.A. and Misra, S.S. (1938). The estimation of the bacterio
cidal power of the blood. J. Hyg. Camb. 38, 732-749.
Miller , W.A. (1964). The microbiology of some self-service
packaged sausage meats. J. Milk Food Technol. 27, 1-3.
Moerman, P.C., Krol, B., Kampelmacher, E.H. and van Noorle
Jansen, L.M. (1966). Invloed van natriumsulfiet op de groei

363,
van Salmonellae en E. coli in rauw gehakt. Conserva 14, 172-175.
Molin, G. (1981). The taxonomy of psychrotrophic Pseudomonas spp.
and their distribution on meat and fish. J. appl. Bact. 51, x.
Molin, G . and Ternstrom, A. (1982). Numerical taxonomy of psychro
trophic pseudomonads. J. gen. Microbiol. 128, 1249-1264.
Miller, V. (1955). Simplified tests for some amino acid decarboxylases
and for the arginine dihydrolase system. Acta pathologica et
microbiologica Scandinavica 36, 158-217.
Monier-Williams, G.W. (1927). Determination of sulphur dioxide in
foods. Reports of Public Health and Medical Subjects. Ministry
of Health, London, 43, 1-56.
Mossel, D.A.A. (1957). The presumptive enumeration of lactose
negative as well as lactose positive Enterobacteriaceae in
foods. Appl. Microbiol. 5, 379.
Mossel, D.A.A. and Ingram, M. (1955). Physiology of the microbial
spoilage of foods. J. appl. Bact. 18, 233-268.
Mossel, D.A.A. and Ratto, M.A. (1970). Rapid detection of sub-
lethally-impaired cells of Enterobacteriaceae in dried foods.
Appl. Microbiol. 20, 273-275.
Mossel, D.A.A., Koopmans, M.J. and Jongerius, E. (1967). Enumeration
of Bacillus cereus in foods. Appl. Microbiol. 15, 650-653.
Moussa, R.S. (1973). The effect of potassium sulphite on the recovery
of micro-organisms from dehydrated onion. J. Sci. Fd. Agric.
24, 401-406.
Mrak, E.M. and Bonar, L. (1938). A note on yeasts obtained from
slimy sausage. Fd. Res. 3, 615-618 .
Mrak, E.M. andPhaff, H.J. (1948). Yeasts Ann. Rev. Microbiol.
2, 1 - 4 6 .

36 4 .
Mulder, S.J. (1969). Bewaren van gehakt bij lage temperaturen.
Koeltechniek 228-230.
McCall, C.E., Martin, W.T. and Boring, J.R. (1966). Efficiency of
cultures of rectal swabs and faecal specimens in detecting
Salmonella carriers: correlation with numbers of Salmonellas
excreted. J. Hyg. 64, 261.
McCrady, M.H. (1915). The numerical interpretation of fermentation
tube results. J. infect, dis. 17, 183.
McDonagh, V.P. and Smith, H.J. (1958). The significance of the
abattoir in Salmonella infection in Bradford. J. Hyg. Camb.
271.
McKinley, G.A., Fagerberg, D.J., Quarles, C.L., George, B.A.,
Wagner , D.E. and Rollins, L.D. (1980). Incidence of
Salmonellae in fecal samples of production swine and swine at
slaughter plants in the United States in 1978. Appl. &
Environ. Microbiol. 40, 562-566.
McLean, R.A. and Sulzbacher, W.L. (1953). Microbacterium thermo
sphactum, spec. nov., a non-heat resistant bacterium from
fresh pork sausage. J. Bact. 65, 428-433.
Narayan, K.G. (1966). Studies on the clostridia incidence in the
beef cattle. Acta vet. Acad. sci. hung. 16, 65-72.
Narayan, K.G. and Takacs, J. (1966). Incidence of clostridia in
emergency-slaughtered cattle. Acta vet. Acad. sci. hung. 16,
345-350.
Neuberg, C. (1929). Uber das verhalten des glucose-schwefligsauren
natriums zu hefe und damit zugammenhangende fragen. Biochem.
Z. 21^ ,477-517.
Neuberg, C. and Reinfurth, E. (1918). Cited by Stratford (1983).

365
Biochemische Zeitschrift 92, 234.
Neuberg, C. and Reinfurth, E. (1919). cited by Stratford (1983).
Berichte de Deutschen Chemie Gesellschaft 52, 1677.
Newbold, R.P. and Scopes, R.K. (1971). Post-mortem glycolysis in
ox skeletal muscle: effects of mincing and of dilution with
or without addition of orthophosphate. J. Fd. Sci. 3j6, 209.
Newell, K.W., McClarin, R., Murdock, C.R., MacDonald, W.H. and
Hutchinson, H.L. (1959). Salmonellosis in Northern Ireland
with special reference to pigs and Salmonella contaminated pig
meal. J. Hyg. Camb. 57, 92-99.
Newton, K.G. (1979). Value of coliform tests for assessing meat
quality. J. appl. Bact. 47, 303-307.
Newton, K.G. and Gill, C.O. (1978). The development of the anaerobic
spoilage flora of meat stored at chill temperatures. J. appl.
Bact. 44, 91-95.
Newton, K.G. and Gill, C.O. (1980). Control of spoilage in vacuum
packaged dark, firm, dry (DFD) meat. J. Food Technol. 15,
227-234.
Newton, K.G., Harrison, J.C.L. and Wauters, A.M. (1978). Sources
of psychrotrophic bacteria on meat at the abattoir. J. appl.
Bact. 45, 75-78.
Ng, L JC. and Stiles, M.E. (1978). Enterobacteriaceae in ground
meats. Can. J. Microbiol. 1574-1582.
Nickel, R. and Gisske, W. (1941). Blut-und lymphgefasssystem des
darmes als infektionspforte. Zeitschrift fiir fleisch-und
milchhygiene d , 225-227, 239-241, 257-259.
Nishimura, S. (1972). Progr. Nucl. Acid Res. Mol. Biol. 12,
49.
Njoku-Obi, A.N., Spencer, J.V., Sauter, E.A. and Eklund, M.W. (1957).

366,
A study of the fungal flora of spoiled chlortetracycline
treated chicken meat. Appl. Microbiol. 319-321.
Norval, J. (1961). Hygiene in slaughterhouses. Vet. Rec. ^ , 781-784.
Nottingham, P.M., Penney, N. and Harrison, J.C.L. (1974). Microbiology
of beef processing. New Zealand J. Agric. Res. 27, 79-83.
Ockerman, H.W., Cahill, V.R., Weiser, H.H., Davies, C.E. and
Siefker, J.R. (1969). Comparison of sterile and inoculated
beef tissue. J. Fd. Sci. 34, 93-99.
Oglivy, W.S. and Ayres, J.C. (1953). Post-mortem changes in stored
meats. V. Effect of carbon dioxide on microbial growth on stored
frankfurters and characteristics of some micro-organisms isolated
from them. Food Res. 18, 121.
Oka, S. (1964). Mechanism of antimicrobial effect of various food
preservatives. In"Microbial inhibitors in Foods", 4th Int.
Sÿmp. Food Microbiol, Goteborg, Sweden.
Okafor, N. (1975). Preliminary microbiological studies on the
preservation of palm wine. J. appl. Bact. 38, 1-7.
Orla-Jensen, S. (1919). In "The lactic acid bacteria." Host and
Son, Copenhagen.
Pain, J. (1981). The recovery of Salmonella from British fresh pork
sausages. Project Rpt. No. 494, University of Bath,England.
Palleroni, N.J. and Doudoroff, M. (1972). Some properties of taxonomic
sub-divisions of the genus Pseudomonas. In "Annual Review of
Phytopathology, 1£, 73-100, (ed K.F. Baker, G.A. Zentmyer,
E.B. Cowling) Palo Alto, California, Ann. Reviews Inc.
Parnow, R.J. (1972). Computer program estimates - bacterial densities
by means of MPN. J.Food Technol, 26, 56-62.
Patterson, J.T. (1969). Salmonellae in meat and poultry, poultry plant

367
cooling waters and effluents and animal feedings stuffs. J. appl.
Bact. 32, 329-337.
Patterson, J.T. and Gibbs, P.A. (1977). Incidence and spoilage
potential of isolates from vacuum-packaged meat of high pH value.
J. appl. Bact. ^ , 25-38.
Patterson, J.T. and Gibbs, P.A. (1978). Sources and properties of
some organisms isolated in two abattoirs Meat Sci . 2, 263-273.
Paule, R. (1971). Contribution h l'étude biochimique du genre
Lactobacillus par une méthode normalisée. Ph.D.Thesis,
University of Lyon, France.
Pfleiderer, G., Jeckel, D. and Wieland, T.H. (1956). Action of
sulphite on diphosphopyridine nucleotide (DPN) hydrogenating
enzymes. Biochem. Z. 325, 187.
Pomerary, B.S. and Grady, M.K. (1961). Salmonella organisms isolated
from feed ingredients. Proc. 65th Ann. Meeting U.S. Livestock
Sanitary Assoc. 449.
Ponting, J.D. and Johnson, G. (1945). Determination of sulfur
dioxide in fruits. Ind. Eng. Chem. Anal. Ed. 17, 682-686.
Prater, A.N., Johnson, C.M., Pool, M.F. and MacKinney, G. (1944).
Determination of sulphur dioxide in dehydrated foods. Ind.
Eng. Chem. Anal. Ed. 16, 153-157.
Pringle, J.R. and Mor, J.R. (1975). Methods for monitoring the
growth of yeast cultures and for dealing with the clumping
problem. In "Methods in Cell Biology" (ed. Prescott, D.M.)
Vol. XI., Academic Press, London, 131-167.
Pusztai, S. (1970). Enterococcus occurrence in organs and muscles
of slaughtery swine. Acta vet. Acad. sci. hung. 20, 391-395.
Radouco-Thomas, C ., Lataste-Dorolle, C., Zender, R., Busset, R.,

368
Meyer, H.M. and Mouton, R.F. (1959). The anti-autolytic effect
of epinephrine in skeletal muscle: non additive process for
preservation of meat. Food Res. 24, 453-482.
Rahn, O. and Conn, J.E. (1944). Effect of increase in acidity
on antiseptic effiency. Ind. Engng. Chem., ind. Edn. 185.
Rankine, B.C. and Pocock, K.F. (1969). Influence of yeast strain
on binding of sulphur dioxide in wines and on its formation during
fermentation. J. Sci. Food S Agric. 20, 104-109.
Rasch, B., Lie, 0 and Yndestad, M. (1978). Bakteriefloraen i
svinesvor og forskjellige svimetoders innflytelse p5 floraen.
Nordisk. Veterinaermedicin 30, 274-281.
Reed, G. and Peppier, H.J. (1973). In "Yeast Technology" Westport,
Connecticut : AVI.
Rehm, H.J. (1964). The antimicrobial action of sulphurous acid.
In "Microbial inhibitors in food" (ed. Molin, N.) Almquist and
Wiksells, Uppsala.
Rehm, H.J. and Wittman, H. (1962). Beitrag zur kenntnis der anti-
mikrobiellen wirkung der schwefligen saure. I. Ubersicht tlber
einflussnehmende faktoren auf die antimikrobielle wirkung
der schwefligen s3ure. Z. Lebensmittelunters. und-Forsch.
413.
Rehm, H.J. and Wittman, H. (1963). Beitrag zur kenntnis der anti-
mikrobiellen wirkung der schwefligen saure. II. Die wirkung
der dissoziierten und undissoziierten S*aure auf verschiedene
mikroorgaraismen. Z. Lebensmittelunters. und-Forsch 120, 465.
Reith, A.F. (1929). Bacteria in the muscular tissue and blood
of apparently normal animals. J. Bact. 12, 367-383.
Richardson, K.C. (1970). Chemical preservatives in foods in
Australia. Fd . Pres. Quart. 30, 2-8.

369 .
Robakis, N.K. (1980)• Growth inhibition of bacteria by bisulphite.
Ph.D.Thesis, University of New York, U.S.A.
Roberts, A.C. and McWeeny, D.J. (1972). The uses of sulphur dioxide
in the food industry. J. Fd. Technol. 7, 221-238.
Roberts, D., Boag, K., Hall, M.L.M. and Shipp, C.R. (1975).
The isolation of salmonellae from British pork sausages and
sausage meat. J. Eyg. Camb. 75, 173-184.
Roberts, T.A. (1980). Contamination of meat. The effects of slaughter
practices on the bacteriology of the red meat carcass.
Royal Soc. Hlth. J. 100, 3-9.
Roberts, T.A , Britton, C.R. and Hudson, W.R. (1980). The
bacterial quality of minced beef in the United Kingdom. J.
Hyg. Camb. S5, 211-218.
Robson, N.C. (1968). Sulphur dioxide as a fruit preservative. Proa.
I.F.S.T. 1, 8-11.
Rogers, R.E. and McCleskey, C.S. (1957). Bacteriological quality of
ground beef in retail markets. Food Technol. ^ , 318-320.
Rogers, P.J., Taylor, V.K. and Egan, A.F. (1980). Energetics of
growth of Microbacterium thermosphactum at low temperatures.
Arch. Microbiol. 128, 152-156.
Rommeler, (1909). cited by Cary, (1916). Zentralblatt fUr
Bakteriologie., Abt. I, 50, 501.
Rose, A.H. (1976). In "Chemical Microbiology" 3rd edition.
Butterwo rth s, London.
Ruben, H.L., Scherago, M. and Weaver, R.H. (1942). The occurrence of
Salmonella in the lymph glands of normal hogs. Amer. J. Hyg.
36, 43 .

370.
Sackin, M.J. and Jones, D. (1976). Numerical methods in the
classification and identification of bacteria with especial
reference to the Enterobacteriaceae. In "Microbiological
classification and identification" (eds. Goodfellow, M. and
Board, R.G. ). Academic Press, London, 73-106.
Sakazaki, R ., Tamura, K., Johnson, R. and Colwell, R.R. (1976).
Taxonomy of some recently described species in the family
Enterobacteriaceae. Int. J. Syst. Bacterial. 26, 158-79.
Sanwal, B.D. (1970). Allosteric controls of amphibolic pathways
in bacteria. Bacterid. Rev. 20-39.
Savage, W.G. (1908). cited by Cary (1916). Med. Press & Circ.
London 86, 167.
Schanderl, H. (1959). In "Die mikrobiologie des mostes und weines"
Eugen Ulmer, Stuttgart, 244.
Schimz, K.L. (1980). Effect of sulphite on the yeast Saccharomyces
cerevisiae. Archives of Microbiol. 125, 89-95.
Schimz, K.L. and Holzer, H. (1979). Rapid decrease of ATP content
in intact cells of Saccharomyces cerevisiae after incubation
of sulfite. Archives of Microbiol. 121, 225-229.
Schleifer, K.H. (1970). Die mureintypen in der gattung Microbacterium
Arch. Mikrobiol. 71, 271-282.
Schleifer, K.H. and Kandler, O. (1972). Peptidoglycan types of
bacterial cell walls and their taxonomic implications. Bacterid.
Rev. 56, 407-477.
Scholfield, J., Menon, T.G. and Lam, C.W. (1981). Psychrotroph
contamination of pig carcasses. Proc. Europ. Meat Workers
Assoc. Vienna G2 , 621-624.
Schotts, E.B., Martin, W.T. and Galton, M.M. (1961). Further studies

371
on Salmonella in human and animal foods and in the environment
of processing plants. Proc. 65th Ann. Meeting U.S. Livestock
Sanitary Assoc, 309.
Schroeter, L.C. (1966). In "Sulphur dioxide applications in foods,
beverages and pharmaceuticals." Pergamon Press, Oxford.
Scott, W.J. (1936). The growth of micro-organisms on ox muscle.
The influence of water content of substrate on rate of growth
at -1°C. J. Coun . Sci. Ind. Res. 9, 177-190.
Scott, W.J. (1957). Water relations of food spoilage micro-organisms.
Adv. in Food Res. 7, 83-127.
Sears, H.J. and Putnam, J.J. (1923). Gas production by bacteria in
symbiosis. J. infect. Dis. 32, 270-279.
Sharp, J.G. (1963). Aseptic autolysis in rabbit and bovine muscle
during storage at 37°C. J. Sci. Food Agric. 14, 468-479.v W
Sharpe, M.E. (1979). Indentification of the lactic acid bacteria
In "Identification methods for microbiologists"(eds.
Skinner, F.A. and Lovelock, D.W. ). Academic Press, London.
233-259.
Shaw, B.G. and Latty, J.B. (1982). A numerical taxonomic study of
Pseudomonas strains from spoiled meat. J. appl. Bact. 52,
219-228.
Shaw, N. and Stead, D. (1970). A study of the lipid composition of
Microbacterium thermosphactum as a guide to its taxonomy.
J. appl. Bact. 33, 470-473.
Shaw, S. and Keddie, R.M. (1983). A numerical taxonomic study of
the genus Kurthia with a revised description of Kurthia zopfii
and a description of Kurthia gibsonii sp. nov. System. Appl.
Microbiol. 4, 253-276.

372
Shay, B.J., Grau, F.H., Ford, A.L., Egan, A.F. and Ratcliff, D.
(1978). Microbiological quality and storage life of sliced
vacuum-packed small goods. Food Technol, Aust. 30, 48-54.
Shewan, J.M. (1974). The biodeterioration of certain proteinaceous
foodstuffs at chill temperatures. In "Industrial aspects of
biochemistry" (ed. B. Spencer). F.E.B.S.
Shewan, J.M., Hobbs, G . and Hodgkiss, W. (1960a). A determinative
scheme for the identification of certain groups of Gram-negative
bacteria, with special reference to Pseudomonadaceae. J. appl.
Bact.23, 379-390.
Shewan, J.M., Hobbs, G. and Hodgkiss, W. (1960h). The Pseudomonas
and Achromohacter groups of bacteria in the spoilage of marine
white fish. J. appl. Bact. 23, (3), 463-468.
Shipton, J. (1959). Estimation of sulphur dioxide in foods. Food
Pres. Quart. 14, 54.
Sierra, G . (1957). A simple method for the detection of lipolytic
activity of micro-organisms and some observations on the influence
of the contact between cells and fatty substrates. Antonie
van Leeuwenhoek 23, 15-32.
Silliker, J.H. (1980). Status of Salmonella - Ten years later.
J. Food Prot. 43, 307.
Skerman, V.B.D., McGowan, V. and Sneath, P.H.A. (1980). Approved
list of bacterial names. Int. J. Sytematic Bacteriol. 30,
225-420,
Sneath, P.H.A. (1977). A method for testing the distinctness of
clusters: A test of the disjunction of two clusters in
Euclidean space as measured by their overlap. Math.Geol. 9,
123-143.
Sneath, P.H.A. (1978). Classification of micro-organisms. In "Essays

373
in microbiology" (eds. J.R. Norris, M.H. Richmond) Wiley and
Sons, Chichester.
Sneath, P.H.A. (1979). Basic program for a significance test for
two clusters in Euclidean space as measured by their overlap.
Comput. Geosci. 5, 143-155.
Sneath, P.H.A. (1980). BASIC program for determining overlap
between groups in an identification matrix of percent positive
characters. Computers and Geosci. 6, 27-34.
Sneath, P.H.A. and Johnson, R. (1972). The influence on numerical
taxonomic similarities of errors in microbiological tests. J.
gen. Microbiol. 72, 377-392.
Sneath, P.H.A. and Jones, D. (1976). Brochothrix, a new genus
tentatively placed in the family Lactobacillaceae. Int. J.
Syst. Bacteriol. 26, 102-104.
Sneath, P.H.A. and Sokal, R.R. (1973). In "Numerical taxonomy:
The principles and practice of numerical classification" W.
Freeman, San Francisco.
Sneath, P.H.A., Stevens, M. and Sackin, M.J. (1981). Numerical
taxonomy of Pseudomonashased on published records of substrate
utilization. Antonie v. Leeuwenhoek 47, 423-448.
Snijders, J.M.A. (1976). Pig slaughtering hygiene. Vet. Med.
Diss. Utrecht.
Snijders, J.M.A. and Gerats, G.E. (1976). Hygiene bei der schlachtung
von schweinen. IV. Bakteriologische beschaffenheit der Schlach-
ttierkorper wahrend verschiedener schlachtphasen. Fleischwirt-
schaft 56, 717-721.
Sojka, W.J., Wray, C ., Shreeve, J. and Benson, A.J. (1977).
Incidence of salmonella infection in animals in England and

374 .
Wales, 1968-1974. J. Hyg. Camb. 7^, 43-56.
Spencer, R. (1964). Cited as pers. comm, by Blood, R.M. (1969) ,
"Salmonellae in Food" B.F.M.I.R.A.
Stanier, R.Y., Palleroni, N.J. and Doudoroff, M. (1966). The aerobic
pseudomonads: a taxonomic study. J. Gen. Microbiol. 43, 159-271.
Stanley, G., Shaw, K.J. and Egan, A F. (1981). Volatile compounds
associated with spoilage of vacuum-packed sliced luncheon
meat by Brochothrix thermosphacta. Appl. & Environ. Microbiol.
41, 816-818.
Stiles , M.E. and Ng, L.K. (1981a). Biochemical characteristics
and identification of Enterobacteriaceae isolated from meats.
Appl. & Environ. Microbiol. 41, 639-645.
Stiles, M.E. and Ng, L.K. (1981b). Enterobacteriaceae associated
with meats and meat handling. Appl. Environ. Microbiol. ^ ,
867-872.
Stolle, A. (1981). Spreading of Salmonellae during cattle slaughtering.
J. appl. Bact. 50, 239-245.
Stolle, A. and Reuter, G. (1978) . Die nachweisbarkeit von
Salmonellen bei Klinisch gesunden schlachtrindern im bestand,
nach dem transport zum schlachthof und wahrend des schlach-
tprozesses. Berliner und Münchener Tierarztliche Wochenschrift
9i, 188-193.
Stott, J.A., Hodgson, J.E. and Chaney, J.C. (1975). Incidence of
salmonellae in animal feed and the effect of pelleting on
content of Enterobacteriaceae. J. appl. Bact. 39, 41-46.
Stratford, M. (1983). Sulphite metabolism and toxicity in Saccharomyces
cerevisiae and Saccharomycodes lugwidii. PhD Thesis,
University of Bath, England.

375
Sullivan, S.M. (1971). The use of sulphur dioxide in foods.
B.F.M.I.R.A. Tech. Circ. No. 479.
Sulzbacher, W.L. (1950). Survival of micro-organisms in frozen meat.
Food Technol. 4, 385-390.
Sulzbacher, W.L. and McLean, R.A. (1951). The bacterial flora of
fresh pork sausage. Food Tech. Champaign 5, 7.
Surkiewicz, B.F., Johnston, R.W., Elliott, R.P. and Simmons, E.R.
(1972) . Bacteriological survey of fresh pork sausage produced
at establishments under federal inspection. Appl. Microbiol.
23, 515 .
Sutherland, J.P., Patterson, J.T. and Murray, J.G. (1975a). Changes
in the microbiology of vacuum-packaged beef. J. appl. Bact. 39,
227-237.
Sutherland, J.P., Patterson, J.T., Gibbs, P.A. and Murray, J.G.
(1975b). Some metabolic and biochemical characteristics of
representative microbial isolates fron vacuum-packaged beef.
J. appl. Bact. 39, 239-249.
Tanaka, N. and Luker, C. (1978). Effect of sulphite on micro-organisms,
(abstract only). Ann. Meeting Am. Soc. Microbiol. No. P12
p. 188.
Theno, D.M., Siegal, D.G. and Schmidt, G.R. (1978). Meat massaging:
effects of salt and phosphates on the ultrastructure of cured
porcine muscle. J. Food Sci. 488.
Thompson, R.E. (1927). Irregularities in the test for B. coli
in water. J. Bact. 13, 209-222.
Thornley, M.J. (1957). Observations on the microflora of minced
c h i c k e n meat irradiated with 4 MeV cathode rays. J. appl.
Bact. 20, 286-298.

376
Threifeil, E.J., Hall, M.L.M. and Rowe, B. (1983). Lactose-fermenting
salmonellae in Britain. F.E.M.S. Microbiol. Letts. 17, 127-130.
Tompkin, R.B., Christiansen, L.N. and Shaparis, A.B. (1980).
Antibotulinal efficacy of sulfur dioxide in meat. Appl.
Environ. Microbiol. 39, 1096-1099.
Tsai, R.Y.T., Goepfert, J.M., Cassens, R.G. and Briskey, E.J. (1971).
A comparison of Salmonella-excretion by stress-susceptible and
stress-resistant pigs. J. Fd. Sci. 36, 889-891.
Turnbull, P.C.B. and Rose, P. (1982). Campylobacter jejuni and
Salmonella in raw red meats. J. Hyg. 88, 29-37.
Tyson, C.B. (1976). A study of sulphite, sorbate and polyphosphate
as preservatives of pork sausage meat. B.Sc. Project Report.
No. 302, University of Bath,England,
van der Slooten, (1907). Cited by Cary (1916). Inaug. Diss.
Bern.
Vanderzant, C. and Nickelson, R. (1969). A microbiological examination
of muscle tissue of beef, pork and lamb. J. Milk Food Technol.
32, 357-361.
Vas, K. (1949). The equilibrium between glucose and sulfurous acid.
J. Soc. Chem. Ind. 68, 340-343.
Vas, K. and Ingram, M. (1949). Preservation of fruit juices with
less sulphur dioxide. Food Manufact. 24, 414-416.
Vernon, E. and Tillet, H.E. (1974). Food poisoning and Sal.monella
infections in England and Wales, 1969-1972. Publ. Hlth., Lond.
85, 225-235.
Vickery, J.R. (1936). The action of micro-organisms on fat. I. The
hydrolysis of beef fat by some bacteria and yeasts tolerating
low temperatures. J. Coun. Sci. Industr. Res. Aust. 9, 107.

377 .
von Graevenitz, A. (1977). The role of opportunistic bacteria in
human disease. Ann. Revs. Microbiol. 31, 447-471.
Walker, A.W. and Ayres, J.C. (1959). Characteristics of yeasts
isolated from processed poultry and the influence of tetracyclines
on their growth. Appl. Microbiol. 7, 251-255.
Walker, A.W. and Ayres, J.C. (1970). Yeasts as spoilage organisms.
In "The Yeasts" vol. 3 (Eds. Rose, A.H. and Harrison, J.S.).
Academic Press, London, 464-527.
Walker, J.H.C. (1957). Organic fertilizers as a source of Salmonella
infection. Lancet, ii, 283-284.
Wallndfer, P. and Rehm, H.J. (1965). Antibiotic action of sulphurous
acid. Z. Lebensmittelumters. u-Forsch 127, 195-206.
Warth, A.D. (1977). Mechanism of resistance of Saccharomyces baileii
to benzoic, sorbic and other weak acids used as food preservatives,
J. appl. Bact. 43, 215-230.
Watson, W.A. (1975). Salmonellosis and meat hygiene: red meat.
Vet. Rec. 96, 374-376.
Wedzicha, B.L. and Bindra, N.J. (1980). A rapid method for the
determination of sulphur dioxide in dehydrated vegetables. J.
Food Sci. Agric. 31, 286-288.vw
Wedzicha, B.L. and Johnson, M.K. (1979). A variation of the Monier-
Williams distillation technique for the determination of sulphur
dioxide in ginger ale. Analyst 104, 694-696.
Weeks, C. (1969). Production of sulfur dioxide-binding compounds and
of sulfur dioxide by two Saccharomyces yeasts. Am. J. E n d .
vitic. 20, 32-39.
Weidemann, J.F. (1965). A note on the microflora of beef muscle
stored in nitrogen at 0°C. J. appl. Bact. 28, 365-367.
Weissman, M.A. and Carpenter, J.A. (1969). Incidence of salmonellae

3 7 8 .
in meat and meat products. Appl. Microbiol. 17, 899.
Weitzmann, P.D.J. (1966). Regulation of citrate synthase activity in
Escherichia coli. Biochim. et Biophys. Acta 128, 213-215.
Weitzmann, P.D.J, (1980). Citrate synthase and succinate thiokinase
in classification and identification. In "Microbiological
Classification and Identification" (eds. Goodfellow, M. and
Board, R.G.), Academic Press, London, 107-125.
Weitzmann, P.D.J. and Jones, D. (1968). Regulation of citrate synthase
and microbial taxonomy. Nature, Lond. 219, 270-272.
Wells, F.E. and Stadelman, W.J. (1958). Development of yeasts on
antibiotic-treated poultry. Appl. Microbiol. 6, 420-422.
West, R.L., Berry, B.W., Smith, G.C., Carpenter, Z.L. and Hoke, K.E.
(1972). Microbial studies in a beef distribution centre. J.
Animal Sci. 35, 209-210.
Whittenbury, R. (1963). The use of soft agar in the study of conditions
affecting the utilization of fermentable substrates by lactic
acid bacteria. J. gen. Microbiol. 32, 375-384.
Wickerham, L.J. (1946). Critical evaluation of nitrogen assimilation
tests commonly used in the classification of yeasts. J. Bact.
52, 293-301.
Wilkinson, B.J. and Jones, D. (1977). A numerical taxonomic survey
of Listeria and related bacteria. J. gen. Microbiol. 98, 399-421.
Williams, L.P. Jr. and Newell, K.W. (1970). Salmonella excretion
in joy-riding pigs. Amer. J. Pub. Hlth. 60, 926.
Willingale, J.M. and Briggs, C.A.E. (1955). The normal intestinal
flora of the pig. II. Quantitative bacteriological studies.
J. appl. Bact. 18, 284.
Wilson, E., Paffenbarger, R.S., Foter, K.J. and Lewis, K.H. (1961).
Prevalence of salmonellae in meat and poultry products. J.

379
infect, dis. 109, 165.
Wilson, G.S. and Miles, A.A. (1964). In "Topley and Wilson's
Principles of Bacteriology and Immunity" 5th Edition Edward
Arnold. London.
Wilson, G.S., Twigg, R.S., Wright, R.C., Hendry, C.B., Cowell, M.P.
and Maier, I. (1935). Spec. Rep. Ser. med. Res. Coun .
London No. 206.
Wishart, D. (1978). In "Clustan user manual" 3rd edition. Inter-
University /Research Councils Series Report No. 47. Program library
unit,Edinburgh University, Scotland.
Wolin, E.F., Evans, J.B., and Niven, C.F. (1957). The microbiology of
fresh and irradiated beef. Food Res. 22, 682-686.
Zagaevskii, I.S. (1973). Beef contamination with Clostridium
perfringens. Veterinariya 1, 101-103.
Zender, R., Lataste-Dorolle, C., Collet, R.A., Rowinski, P. and
Mouton, R.F. (1958). Aseptic autolysis of muscle: biochemical
and microscopic modifications occurring in rabbit and lamb
muscle during aseptic and anaerobic storage. Food Res. ^ ,
305-326.