(chemotype - infection and immunityiai.asm.org/content/61/8/3216.full.pdf · medizin, w-2061...
TRANSCRIPT

Vol. 61, No. 8INFEcrION AND IMMUNrrY, Aug. 1993, p. 3216-32210019-9567/93/083216-06$02.00/0Copyright X 1993, American Society for Microbiology
Specificity of Rabbit Antisera against the RoughLipopolysaccharide of Salmonella minnesota R4
(Chemotype Rd2P)ANNA SWIERZKO,1t LORE BRADE,1 HANS PAULSEN,2 AND HELMUT BRADEl*
Division ofBiochemical Microbiology, Forschungsinstitut Borstel, Institut fir Experimentelle Biologie undMedizin, W-2061 Borstel,j and Institut fur Organische Chemie der Universitat Hamburg,
W-2000 Hamburg 2 Gerany
Received 19 October 1992/Accepted 12 May 1993
Rabbit polyclonal antibodies against the rough mutant lipopolysaccharide (LPS) ofSalmoneUla minnesota R4(chemotype Rd2P-) were serologically characterized by using R4 LPS, deacylated LPS, dephosphorylated LPS,and synthetic partial structures, including compounds comprising the core region of Rd2P- LPS bound to the131-6-linked glucosamine disaccharide with two amide-linked 3-hydroxytetradecanoic acid residues or coupledto bovine serum albumin. By using a passive hemolysis assay and an enzyme immunoassay and absorption andinhibition experiments, the antibody specificities present could be determined. One group of antibodiesrequired components of the core oligosaccharide (with or without the side chain 3-deoxy-D-manno-octulosonicacid [KdoJ) and the phosphorylated glucosamine disaccharide of the lipid A moiety for binding. Thephosphate-independent antibodies were directed against the core oligosaccharide, recognizing an epitopeconsisting of one terminal heptose linked to Kdo or to the reducing moiety of the ca2-4-linked Kdodisaccharide. Antibodies requiring the presence of acyl residues and those reacting with a single heptose or Kdoresidue were not detected.
Lipopolysaccharides (LPS) are common constituents ofthe outer membrane of gram-negative bacteria. They func-tion as the major surface antigens and have numerousphysiological and pathophysiological activities, for whichLPS are also called endotoxins. LPS are chemically com-posed of a lipid part, called lipid A, which is responsible formost of the endotoxic activities of LPS, and a heteropolysac-charide which generally consists of an oligosaccharide,termed the core region, and, in some bacterial families, ofthe so-called 0 chain, composed of repeating oligosaccha-ride units (17).
Antibodies against the evolutionarily well conserved andsimilarly structured core and lipid A region have beenconsidered to exhibit in vitro cross-reactivity and in vivocross-protection against the deleterious effects of endotoxins(1, 16). In order to provide a molecular basis for theunderstanding of the immunoreactive properties of bacterialLPS, we have characterized poly- and monoclonal antibod-ies against lipid A (4, 11), and against the LPS of chlamydiae(5) and Re chemotype mutants of enterobacteria (18). Allinvestigations were carried out with natural LPS and definednatural or synthetic partial structures. We found to oursurprise that the majority of antibodies in antisera againstRe-type and chlamydial LPS bound to a complex epitopecomposed of core and lipid A constituents and that onlyminor amounts of antibodies were directed against terminalcore structures, i.e., against 3-deoxy-D-manno-octulosonicacid (Kdo). Kdo is a polyfunctional ketosugar with charged(carboxyl group), hydrophilic (hydroxyl groups), and hydro-phobic (deoxy function) properties which certainly influencethe immunogenic properties of Kdo-containing antigens.
* Corresponding author.t Permanent address: Center for Microbiology and Virology of
the Polish Academy of Sciences, ul. Piotrkowska 179, PL-90-237Lodz, Poland.
Consequently, the immunoreactive properties of chlamydialand Re-type LPS, containing Kdo as the main core sugar,cannot be regarded as representative of the enterobacterialcore region. Therefore, we started to investigate the immu-nogenic and antigenic properties of Rd chemotype LPScarrying heptosyl residues in terminal positions.Here we report on the results obtained with antisera
against the LPS of the Rd2P chemotype.(Some of the results reported here were presented at the
2nd Conference of the International Endotoxin Society,Vienna, Austria, August 1992.)
MATERIALS AND METHODS
Bacteria and bacterial LPS. Rough mutant strain R4 ofSalmonella minnesota (chemotype Rd2P-) was grown in afermentor, killed with phenol (0.5%), washed successivelywith ethanol, acetone, and ether, and then dried. LPS wasextracted from dry bacteria by the phenol-chloroform-petro-leum ether method (8) and purified by repeated ultracentrif-ugation followed by conversion to the uniform triethylam-monium salt after electrodialysis (7). Alkali-treated LPS anddephosphorylated LPS were prepared as described before(18).
Synthetic antigens. (i) Amphiphilic compounds. The N,N'-acylated 01--6-linked glucosamine disaccharide 2-deoxy-2-[(R)-3-hydroxytetradecanamido]-oi-D-glucopyranosyl-(1l-6)-2-deoxy - 2 - [(R) - 3 - hydroxytetradecanamido] - D - glucose wasglycosylated with the following oligosaccharides containingKdo or L-glycero-D-mannoheptopyranose (Hep) as reportedbefore (14, 15): Kdo(2-*6'), Kdo(2-*4)Kdo(2--36'), Hep(1-+5)Kdo(2-- 6'), and Hep(1--'5)[Kdo(2--4)Kdo](2--6') (Fig. 1and 2; Table 1).
(ii) Partial structures of the core oligosaccharide. The allylglycosides of Hep(1-), Hep(1-5)Kdo(ac2-*), Hep(1->5)Kdo (02--), Hep(1-5)[Kdo(2-*4)Kdo] (a2- ), and Hep(1-
3216
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

ANTIBODIES AGAINST S. MINNESOTA Rd2P- LPS 3217
OR
0
HO 0 HO
HO NH HO NH H
SOH FOH
FIG. 1. Chemical structure of synthetic amphiphiles represent-ing partial structures of Re and Rd2P LPS. See also Table 1.
5)[Kdo(2--4)Kdo] (P2-3) were reacted with cysteamine asreported before (12) to give spacer ligands of the generalformula R-(CH2)3-S-(CH2)2-NH3', where R represents thecarbohydrate ligand. The synthetic antigens used in thisstudy are designated as shown in Tables 1 and 2. Syntheticlipid A from Escherichia coli (compound 506 [10]) was kindlyprovided by S. Kusumoto, Osaka, Japan.
Preparation of the deacylated, phosphorylated carbohy-drate backbone. LPS (1 g) was de-O-acylated in absolute
OH
HO
HOOH HO -
HO0
H o-OR
A B
HO OH
0
HO
OH
Ho O
OR
D
HO OH
O0
OOOOH~~~O
0 HO
HO 0 0
0 0
0-OR
F
TABLE 1. Chemical structures of synthetic LPS partialstructures used in this study'
Nature of R Designation
Kdo(2-)............. Kdo-GlcNhm2Kdo(2--4)Kdo(2-*)............. Kdo2-GlcNhm2Hep(l- 5)Kdo(2--) ............. Hep-Kdo-GlcNhm2Hep(1-l5)Kdo(2- ) ............. Hep-Kdo2-GlcNhm2
4t2
Kdo
a See also Fig. 1 and 2. Abbreviations: Kdo, 3-deoxy-D-manno-octulo-sonic acid present as a-pyranoside; Hep, L-glycero-a-D-mannoheptopyra-nose; GlcNhm2, 2-deoxy-2-[(R)3-hydroxytetradecanamidoJ-j-D-glucopyrano-syl-(1-6)2-deoxy-2-[(R)-3-hydroxytetradecanamidoJ-D-glucose.
hydrazine (40 ml) at 37°C for 30 min. The mixture was cooledon ice and slowly poured into chilled acetone (500 ml),sedimented by centrifugation, washed three times in chilledacetone (50 ml each), and lyophilized. The preparation wasthen treated with alkali (4 M KOH, 4 h, 120'C) to cleave theamide-linked fatty acids. The deacylated LPS (LPSdeac) wasprecipitated with ethanol, dissolved in water, neutralized(Amberlite IR 120H+), and extracted with CH2Cl2 to removethe released fatty acids. The water phase was lyophilized to
OHO HOu OH
Ho0
0_ HO
OR OR
C
HO OH
HO 0
HO
OHHO
00-E
HO OH
0
HO
HO
HO OHOH
HOHO
O\~ HO_ eHO o-
0
G
FIG. 2. Chemical structures of synthetic oligosaccharides representing partial structures of the core oligosaccharide of Re (A and B) andRd2P (C, D, and F) LPS. The oligosaccharides shown in E and G are analogs with a ,B-configured Kdo at the reducing end, not occurringin natural LPS. The compounds were synthesized as allyl glycosides (R = allyl). In the antigens used in this study (see also Tables 1 and 2),R represents the acylated glucosamine disaccharide shown in Fig. 1 (amphipathic antigens) or BSA (artificial glycoconjugate antigens) towhich the allylglycosides were conjugated via a spacer molecule (see Materials and Methods).
VOL. 61, 1993
I
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

3218 gWIERZKO ET AL.
TABLE 2. Chemical structures of carbohydrate ligands presentin artificial synthetic glycoconjugates
Chemical structure of Desigation Amt of ligandcarbohydrate liganda (nmol/mg)
Hep(1- R) Hep-BSA 20Hep(1->5)Kdo(2-->R) Hep-Kdo-BSA 40Hep(1->5)Kdo(02--*R) Hep-KdoI3-BSA 24Hep(1->5)Kdo(2--R) Hep-Kdo2-BSA 28
4
2Kdo
Hep(1-5)Kdo(P2-+R) Hep-Kdo2I3-BSA 154I2
Kdoa Kdo was in the a-anomeric form if not stated otherwise. R, -(CH2)3-S-
(CH2)2-NH-(CH2)5-NH-BSA. See also Fig. 2.
give the phosphorylated carbohydrate backbone. Theamount of 3-hydroxytetradecanoic acid was determined bygas-liquid chromatography of the methyl ester to be less than10 nmol/mg.
Preparation of artificial glycoconjugates. The syntheticoligosaccharides described above and the phosphorylatedcarbohydrate backbone of R4 LPS were conjugated tobovine serum albumin (BSA) as described elsewhere (9)(Table 2). The amount of ligand incorporated was deter-mined by chemical analysis, measuring the amount of Kdoby the thiobarbiturate assay (2) or Hep by gas-liquid chro-matography of the alditol acetate or both.
Rabbit antisera. Rabbits with no detectable antibodiesagainst the LPS of S. minnesota rough mutants of the Rd2P-and Re chemotypes were immunized with heat-killed bacte-ria of S. minnesota R4 as follows. Animals received 50, 100,100, or 200 ,ug of heat-killed bacteria intravenously in 0.5 mlof phosphate-buffered saline (PBS) on days 0, 4, 7, and 11,respectively. On day 16, 20 ml of blood was obtained fromthe ear vein (early sera). The animals received a singlebooster injection (500 ,g) on day 51 and were exsanguinatedon day 58 (late sera).
Serological assays. The passive hemolysis assay, hemoly-sis inhibition assay, absorption experiments, and enzymeimmunoassay (EIA) with LPS as a solid-phase antigen wereperformed as described before (11). EIA with LPS-BSAcomplexes and artificial glycoconjugates as a solid-phaseantigen were carried out as reported before (6).
RESULTS
Synthesis of artificial glycoconjugates. The synthetic partialstructures listed in Table 2 were covalently linked to BSA;the amount of ligand incorporated was determined chemi-cally to be in the range of 15 to 40 nmol/mg of conjugate(Table 2). These artificial glycoconjugates allowed us todifferentiate antibodies directed against epitopes locatedexclusively in the core region and not requiring any constit-uents of the lipid A region. In addition, the phosphorylatedcarbohydrate backbone was prepared from natural R4 LPSafter successive de-O- and de-N-acylation. The core regionof R4 LPS consists mainly of the trisaccharide shown in Fig.2F linked to the lipid A backbone. In addition, there existsanother molecular species containing a third Kdo residuewhich is linked to position 4 of the side chain Kdo (19). Theoligosaccharide was coupled via the free amino groups of theglucosamine lipid A backbone to BSA by the glutardialde-hyde method (148 nmol of ligand per mg of conjugate). Thisglycoconjugate also allowed the binding of antibodies di-rected against more-complex epitopes composed of core andlipid A constituents.
Reactivity of rabbit sera against S. minnesota R4 in thepassive hemolysis assay. Four animals were immunized withheat-killed bacteria. Only late sera were collected on day 58from two rabbits, and early and late sera were obtained ondays 16 and 58, respectively, from the other two. Theantisera were tested in the passive hemolysis assay withnative and dephosphorylated LPS and synthetic partialstructures to sensitize sheep erythrocytes (SRBCs). Thedata obtained for animal K236 are representative and areshown in Table 3. All animals had antibodies against LPS,with titers ranging from 3,200 to 25,600; higher titers werefound in early sera. The same reactivity was observed whenSRBCs were coated with de-O-acylated LPS (data notshown). Titers against dephosphorylated LPS and the syn-thetic partial structures Hep-Kdo-GlcNhm2 and Hep-Kdo2-GlcNhm2 were similar, not differing by more than twodilution steps. However, in some cases they were up to twotiter steps lower than those against LPS. Reactivity was notobserved with the partial structures Kdo2-GlcNhm2 andKdo-GlcNhm2, representing the carbohydrate backbone ofRe chemotype LPS (3), or with synthetic E. coli lipid A.Absorption experiments. To differentiate distinct antibody
specificities, absorption experiments were carried out on allsera. The data obtained for antiserum K236 are representa-tive and are shown in Table 4. It is noted that individualsamples were absorbed with only one antigen and notsuccessively with different antigens. Absorption with LPSabolished reactivity against the homologous LPS as well asagainst natural and synthetic partial structures. This indi-
TABLE 3. Antibody titers of sera from rabbit K236 against S. minnesota R4 with homologous LPS and partial structures in thepassive hemolysis assay
Titer obtained with SRBCsa coated with:
Antiserum Natural LPS Synthetic partial structureb
LPS LPS-HFC Hep-Kdo2-GlcNhm2 Hep-Kdo-GlcNhm2 Kdo2-GlcNhm2 Kdo-GlcNhm2 Lipid A
Early (day 16) 25,600 25,600 25,600 25,600 <50 <50 <50Late (day 58) 6,400 1,600 1,600 3,200 <50 <50 <50
a Sensitization of SRBCs was optimized for each antigen.b For detailed structures, see Fig. 1 and 2 and Table 1.c LPS-HF, dephosphorylated LPS.
INF-ECT. IMMUN.
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

ANTIBODIES AGAINST S. MINNESOTA Rd2P- LPS 3219
TABLE 4. Antibody titers of sera from rabbit K236 against S. minnesota R4 LPS and partial structures in the passive hemolysis assaybefore and after absorption'
Titer obtained with SRBCs coated with:
Antiserum Absorbed withb: Natural LPS Synthetic structure
LPS LPS-HF Hep-Kdo2-GlcNhm2 Hep-Kdo-GlcNhm2
Early (day 16)C SRBCs 25,600 25,600 25,600 25,600LPS 100 < 100 < 100 < 100LPS-HF 6,400 < 100 < 100 < 100Hep-Kdo2-GlcNhm2 6,400 < 100 < 100 800Hep-Kdo-GlcNhm2 12,800 1,600 1,600 < 100
Late (day 58) SRBCs 6,400 1,600 1,600 3,200LPS < 100 < 100 < 100 < 100LPS-HF 400 < 100 < 100 < 100Hep-Kdo2-GlcNhm2 800 < 100 < 100 200Hep-Kdo-GlcNhm2 1,600 200 200 < 100
a See Table 3, footnotes a, b, and c.b Antisera (500 .1l, prediluted 1:50) were absorbed at 4°C with 50 p1l of SRBCs or SRBCs coated with the indicated antigens.c Sera taken at day indicated after first immunization.
cated that antibodies directed against partial structures alsorecognized their epitope in native LPS. De-O-acylated LPSgave similar results (data not shown). On the other hand,absorption with any of the partial structures could notremove the antibodies against LPS but led to a titer reduc-tion of two to four dilution steps. Thus, the remainingLPS-reactive antibodies required the presence of phosphorylgroups. These antibodies had higher titers in early sera thanin late sera. Interestingly, absorption with the syntheticpartial structure Hep-Kdo2-GlcNhM2 did not abolish thereactivity towards the even smaller structure Hep-Kdo-GlcNhm2.
Inhibition experiments. To determine whether the phos-phate-independent antibodies against Hep-Kdo-GlcNhm2and Hep-Kdo2-GlcNhm2 required the glucosamine disaccha-ride moiety for binding, inhibition experiments were carriedout. The oligosaccharides Hep-Kdo and Hep-Kdo2, linkedeither to the N-acylated glucosamine disaccharide or toBSA, were used to inhibit the reaction of antiserum K236with SRBCs coated with Hep-Kdo-GlcNhm2 or Hep-Kdo2-GlcNhm2. It is noted that this inhibition system is notaffected by phosphate-dependent antibodies. Both types ofinhibitors were equally active, indicating that the glu-cosamine disaccharide was not necessary for binding (datanot shown). This was also confirmed by EIA (see below).
Reactivity of rabbit sera against S. minnesota R4 in ELT.Late antisera were tested in ETA with LPS, LPS-BSAcomplexes, or natural and synthetic partial structures co-valently linked to BSA as solid-phase antigens. Checker-board titrations were performed in an antigen range from 50fmol to 15 pmol of ligand per well and serial twofold dilutionsof antiserum, starting at a dilution of 1:1,000. Two late seraare shown in Table 5, where only positive reactivities are
listed. None of the sera reacted with Hep-BSA, Hep-Kdo3-BSA, BSA, or glutardialdehyde-activated BSA. The titersagainst LPS, LPS-BSA complexes, and the LPSdCac-BSAconjugate were similar (differences not exceeding one dilu-tion step), in the range of 16,000 to 128,000. Reactivities withthe synthetic glycoconjugates Hep-Kdo-BSA (titers of 4,000to 8,000) and Hep-Kdo2-BSA (titers of 8,000 to 32,000) werealso observed. The antigen Hep-Kdo2j3-BSA, in which thereducing Kdo residue possesses the ,3-anomeric configura-tion, yielded significantly lower titers than the a-configu-rated analog, representing the natural type of linkage. Thetwo other sera not shown in Table 5 had similar reactivities.The reactivity of the late serum K236 with different
amounts of antigen is illustrated in Fig. 3. The results arerepresentative of those with the other sera as well. WhereasLPS-BSA complexes, LPSdeaC-BSA, and Hep-Kdo2-BSAgave a plateau, this was not the case with LPS and Hep-Kdo-BSA, suggesting a lower binding affinity to the lattertwo antigens or a poor coating efficiency. Therefore, anti-body-binding curves were determined with an amount ofantigen at which the antibody binding reached a plateau(between 0.1 and 15 pmol per well). The results are shown inFig. 4. LPS-BSA complexes and LPSdeac-BSA yieldedhigher titers than the other antigens, which also gave flattercurves.
DISCUSSION
In the present study, chemically defined partial structuresobtained from natural sources or by chemical synthesis wereused to characterize hyperimmune sera from rabbits immu-nized with heat-killed bacteria of S. minnesota R4 (chemo-type Rd2P-). All animals had made antibodies against LPS
TABLE 5. Antibody titers of rabbit sera against S. minnesota R4 with homologous LPS and partial structures in EIA
Titer' obtained with:
Late serum Natural LPS Synthetic partial structure
LPS LPS-BSA complex LPSdeac-BSA Hep-Kdo2-BSA Hep-Kdo2[-BSA Hep-Kdo-BSA
K4 32,000 64,000 64,000 32,000 8,000 8,000K236 128,000 64,000 128,000 32,000 1,000 8,000
a Highest dilution yielding an A405 of >0.2 with 7.5 pmol of antigen per well.
VOL. 61, 1993
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
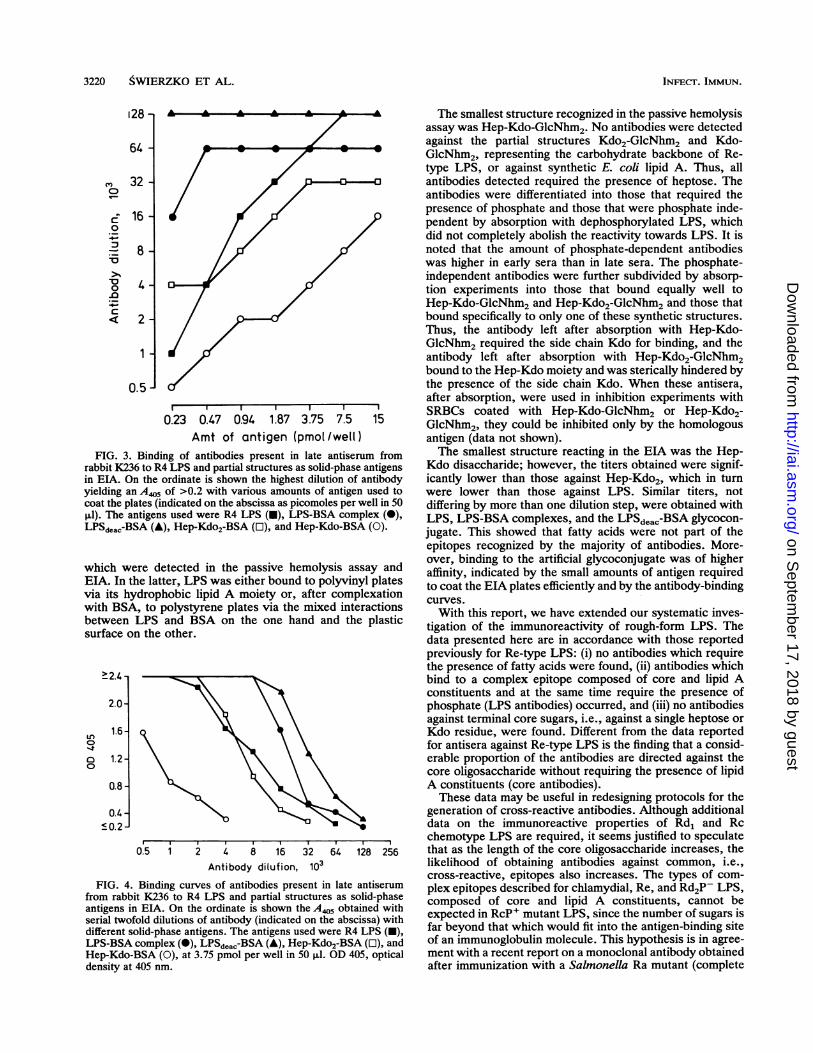
3220 SWIERZKO ET AL.
i28
64
32
16
8
2
FIG. 3. Erabbit K236in ETA. Onyielding an.,coat the plat4.1l). The antiLPSdeac-BSA
which weriEIA. In thevia its hydwith BSA,between Lsurface on
FIG. 4. Efrom rabbitantigens in I
serial twofoldifferent soliLPS-BSA cc
Hep-Kdo-B'density at 4(
The smallest structure recognized in the passive hemolysisassay was Hep-Kdo-GlcNhm2. No antibodies were detectedagainst the partial structures Kdo2-GlcNhm2 and Kdo-GlcNhm2, representing the carbohydrate backbone of Re-type LPS, or against synthetic E. coli lipid A. Thus, allantibodies detected required the presence of heptose. Theantibodies were differentiated into those that required thepresence of phosphate and those that were phosphate inde-pendent by absorption with dephosphorylated LPS, whichdid not completely abolish the reactivity towards LPS. It isnoted that the amount of phosphate-dependent antibodieswas higher in early sera than in late sera. The phosphate-independent antibodies were further subdivided by absorp-tion experiments into those that bound equally well toHep-Kdo-GlcNhm2 and Hep-Kdo2-GlcNhm2 and those thatbound specifically to only one of these synthetic structures.Thus, the antibody left after absorption with Hep-Kdo-GlcNhm2 required the side chain Kdo for binding, and theantibody left after absorption with Hep-Kdo2-GlcNhm2bound to the Hep-Kdo moiety and was sterically hindered bythe presence of the side chain Kdo. When these antisera,after absorption, were used in inhibition experiments with
230470941 87 3 75 7 5 15 SRBCs coated with Hep-Kdo-GlcNhm2 or Hep-Kdo2-0.23 0.47 0.94 1.87 3.75 7.5 15 GlcNhm2, they could be inhibited only by the homologous
Amt of antigen (pmol/well) antigen (data not shown).linding of antibodies present in late antiserum from The smallest structure reacting in the EIA was the Hep-to R4 LPS and partial structures as solid-phase antigens Kdo disaccharide; however, the titers obtained were signif-the ordinate is shown the highest dilution of antibody icantly lower than those against Hep-Kdo2, which in turn4405 of >0.2 with various amounts of antigen used to were lower than those against LPS. Similar titers, notes (indicated on the abscissa as picomoles per well in 50 differing by more than one dilution step, were obtained with:igens used were R4 LPS (E), LPS-BSA complex (), LPS, LPS-BSA complexes, and the LPSdeac-BSA glycocon-k (A), Hep-Kdo2-BSA (E), and Hep-Kdo-BSA (0). jugate. This showed that fatty acids were not part of the
epitopes recognized by the majority of antibodies. More-over, binding to the artificial glycoconjugate was of highere detteced in the passive hemolysis assay and affinity, indicated by the small amounts of antigen required
latter, LPS was either bound to polyvinyl plates to coat the EIA plates efficiently and by the antibody-bindingIrophobic lipid A moiety or, after complexation curves.to polystyrene plates via the mixed interactions With this report, we have extended our systematic inves-tS and BSA on the one hand and the plastic tigation of the immunoreactivity of rough-form LPS. The
data presented here are in accordance with those reportedpreviously for Re-type LPS: (i) no antibodies which requirethe presence of fatty acids were found, (ii) antibodies whichbind to a complex epitope composed of core and lipid Aconstituents and at the same time require the presence ofphosphate (LPS antibodies) occurred, and (iii) no antibodiesagainst terminal core sugars, i.e., against a single heptose orKdo residue, were found. Different from the data reportedfor antisera against Re-type LPS is the finding that a consid-erable proportion of the antibodies are directed against thecore oligosaccharide without requiring the presence of lipidA constituents (core antibodies).These data may be useful in redesigning protocols for the
generation of cross-reactive antibodies. Although additionaldata on the immunoreactive properties of Rd, and Rc
I,,, , , , , , chemotype LPS are required, it seems justified to speculate.5 1 2 4 8 16 32 64 128 256 that as the length of the core oligosaccharide increases, the
Antibody dilufion, 103 likelihood of obtaining antibodies against common, i.e.,cross-reactive, epitopes also increases. The types of com-
3inding curves of antibodies present in late antiserum plex epitopes described for chlamydial, Re, and Rd2P- LPS,K236 to R4 LPS and partial structures as solid-phase plex of cred lamydA and cano beEIA. On the ordinate is shown the A405 obtained with composed of core and lipid A constituents, cannot beId dilutions of antibody (indicated on the abscissa) with expected in RcP' mutant LPS, since the number of sugars isid-phase antigens. The antigens used were R4 LPS (-), far beyond that which would fit into the antigen-binding siteDmplex (-), LPSdeac-BSA (A), Hep-Kdo2-BSA (E), and of an immunoglobulin molecule. This hypothesis is in agree-SA (0), at 3.75 pmol per well in 50 .1l. OD 405, optical ment with a recent report on a monoclonal antibody obtained05 nm. after immunization with a Salmonella Ra mutant (complete
0
c
.0
C -c
>2.4
2.0-
LA
00
1.6-
1.2-
0.8 -
0.4 -
'0.2-1
INFECT. IMMUN.
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

ANTIBODIES AGAINST S. MINNESOTA Rd2P- LPS 3221
core) which recognizes a heptose epitope in a variety ofSalmonella and E. coli LPS (13).
ACKNOWLEDGMENTS
We thank U. Albert and V. Susott for technical assistance and S.Kusomoto (Osaka, Japan) for providing synthetic E. coli lipid A.We thank the Deutscher Akademischer Austauschdienst for fi-
nancial support.
REFERENCES1. Baumgartner, J.-D. 1991. Immunotherapy with antibodies to
core lipopolysaccharide: a critical appraisal. Infect. Dis. Clin.N. Am. 5:753-779.
2. Brade, H., C. Galanos, and 0. Luideritz. 1983. Differentialdetermination of the 3-deoxy-D-mannooctulosonic acid residuesin lipopolysaccharides of Salmonella minnesota rough mutants.Eur. J. Biochem. 131:195-200.
3. Brade, H., and E. T. Rietschel. 1984. a-2- 4-Interlinked3-deoxy-D-mannooctulosonic acid disaccharide: a common con-stituent of enterobacterial lipopolysaccharides. Eur. J. Bio-chem. 145:231-236.
4. Brade, L., K. Brandenburg, H.-M. Kuhn, S. Kusumoto, E. T.Rietschel, and H. Brade. 1987. The immunogenicity and antige-nicity of lipidA are influenced by its physicochemical state andenvironment. Infect. Immun. 55:2636-2644.
5. Brade, L., 0. Holst, P. Kosma, Y.-X. Zhang, H. Paulsen, R.Krausse, and H. Brade. 1990. Characterization of murine mono-clonal and murine, rabbit, and human polyclonal antibodiesagainst chlamydial lipopolysaccharide. Infect. Immun. 58:205-213.
6. Fu, Y., M. Baumann, P. Kosma, L. Brade, and H. Brade. 1992.A synthetic glycoconjugate representing the genus-specificepitope of chlamydial lipopolysaccharide exhibits the sameimmunoreactivity as its natural counterpart. Infect. Immun.60:1314-1321.
7. Galanos, C., and 0. Luderitz. 1975. Electrodialysis of lipopoly-saccharides and their conversion to uniform salt forms. Eur. J.Biochem. 54:603-610.
8. Galanos, C., 0. Liideritz, and 0. Westphal. 1969. A new methodfor the extraction of R lipopolysaccharides. Eur. J. Biochem.9:245-249.
9. Holst, O., L. Brade, P. Kosma, and H. Brade. 1991. Structure,serological specificity, and synthesis of artificial glycoconju-gates representing the genus-specific lipopolysaccharide epitope
of Chlamydia spp. J. Bacteriol. 173:1862-1866.10. Imoto, M., H. Yoshimura, S. Kusomoto, and T. Shiba. 1984.
Total synthesis of lipid A, active principle of bacterial endo-toxin. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 60:285-288.
11. Kuhn, H.-M., L. Brade, B. J. Appelmelk, S. Kusomoto, E. T.Rietschel, and H. Brade. 1992. Characterization of the epitopespecificity of murine monoclonal antibodies directed againstlipid A. Infect. Immun. 60:2201-2210.
12. Lee, R. T., and Y. C. Lee. 1974. Synthesis of 3-(3-aminoethyl-thio)propyl glycosides. Carbohydr. Res. 37:193-201.
13. Nnalue, N. N., S. M. Lind, and A. A. Lindberg. 1992. Thedisaccharide L-a-D-heptosel-- 7-L-a-D-heptosel--* of the innercore domain of Salmonella lipopolysaccharide is accessible toantibody and is the epitope of a broadly reactive monoclonalantibody. J. Immunol. 149:2722-2728.
14. Paulsen, H., and M. Brenken. 1991. Bausteine von Oligosaccha-riden. XCVIII. Synthese von Oligosacchariden der innerenCore- und Lipoid A-Region von Lipopolysacchariden. JustusLiebigs Ann. Chem. 1991:1113-1126.
15. Paulsen, H., and C. Krogmann. 1989. Bausteine von Oligosac-chariden. XCIV. Synthese von KDO-haltigen Tri- und Tetrasac-charid-Sequenzen der inneren Core- und Lipoid A-Region vonLipopolysacchariden. Justus Liebigs Ann. Chem. 1989:1203-1213.
16. Poliack, M. 1990. New therapeutic strategies in gram-negativesepsis and septic shock based on molecular mechanisms ofpathogenesis, p. 1-18. In G. L. Mandell, R. G. Douglas, andJ. E. Bennet (ed.), Principles and practice of infectious diseases,update 8. Churchill-Livingstone, New York.
17. Rietschel, E. T., T. Klrikae, W. Feist, H. Loppnow, P. Zabel, L.Brade, A. J. Ulmer, H. Brade, U. Seydel, U. Zahnnger, M.Schlaak, H.-D. Flad, and U. Schade. 1991. Molecular aspects ofthe chemistry and biology of endotoxin, p. 207-231. 42ndColloquim Mosbach 1991: Molecular aspects of inflammation.Springer-Verlag, Berlin.
18. Rozalsli, A., L. Brade, P. Kosma, B. J. Appelmelk, C. Krog-mann, and H. Brade. 1989. Epitope specificities of murinemonoclonal and rabbit polyclonal antibodies against enterobac-terial lipopolysaccharides of the Re chemotype. Infect. Immun.57:2645-2652.
19. Tacken, A., E. T. Rietschel, and H. Brade. 1986. Methylationanalysis of the heptose 3-deoxy-D-manno-2-octulosonic acidregion (inner core) of lipopolysaccharide from Salmonella min-nesota rough mutants. Carbohydr. Res. 149:279-291.
VOL. 61, 1993
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from