experimental studies on the intrinsic fibers of the cerebellum. ii. the cortico-nuclear projection
TRANSCRIPT

EXPERIMENTAL STUDIES ON THE INTRINSIC FIBERS O F THE CEREBELLUM
11. T H E CORTICO-NUCLEAR PROJECTION
JAN JANSEN AND ALF BRODAL Anatomical Znstitute and Psychiatric Clhic, The University, Oslo, Norwap
TWENTY-POUR FIGURES.
INTRODUCTION
Experimental investigations of the fiber connections of the central nervous system in a great many instances have re- vealed that a definite topographic localization exists between the interconnected centers and nuclei. As regards the cere- bellum a definite pattern of localization has recently been conclusively demonstrated for the olivo-cerebellar connec- tions by Brodal ( '40a) and studies now in progress suggest similar relations within the ponto-cerebellar system. The present publication forms part of a more comprehensive in- vestigation of the morphological foundation for localization within the cerebellum. In this paper we propose to deal with the cortico-nuclear fibers. The experiments to be reported be- low were performed for the purpose of determining to which extent a definite topographic localization exists in the con- nections between the cerebellar cortex and its nuclei.
The literature on the cortico-nuclear fibers of the cerebellum is not very comprehensive although some new papers have appeared since the first part of this investigation (Jansen, '33) was published. Most investigators of the cortico-nuclear systems have confined their studies to determination of the
This research waa aided by a grant to the Anatomical Institute, University of Oslo, by the Rockefeller Fondation. The authors also wish to express their gratitude to Prof. K. E. Schreiner and Prof. E. Langfeldt for their kind support.
267

268 J A N JANSEN A N D ALF BRODAL
main cerebellar subdivisions which project on the individual nuclei without attempting to define the cortico-nuclear rela- tions in such detail as is necessary for discussion of the question of localization. The conclusions arrived a t by the different authors a re assembled in table 1, where the results are chiefly arranged according to Bolk’s terminology which has been used by most of these investigators. (Fo r this reason it was difficult to arrange the table in accordance with one of the more recent principles of cerebellar subdivision). The braces indicate that the author has not distinguished between those divisions of the cerebellum which the braces include. It goes without saying that all details cannot find place in such a table. To some of them, however, we shall return in the discussion. Where two or more nuclei were found to be related to the same lobule, the one with the strongest connections is named first. The table speaks for itself, but a few supplementary remarks may be appropriate.
Clarke and Horsley’s work ( ’05) is the classical paper 011 this subject. Previous to their publication few and incon- clusive investigations had appeared. Fo r the older literature we refer to the review by Clarke and Horsley. The authors in some experiments give a rather detailed account of the cortico-nuclea~- projection. We shall return to this point later on. I n experiments with extensive lesions of the cerebellum Clarke and Horsley also observed degenerated fibers in the vestibular nuclei, suggesting, however, that secondary dis- turbances of the circulation possibly are responsible for this “more extensive association.” They found 110 evidence indi- cating crossing of the fibers in the midline.
Lowy ( ’16) and Brouwer and Coenen ( ’21) studied especially Bolk’s “formatio vermicularis.” Since these authors did not distinguish between the two main divisions of ‘ ‘formatio vermicularis, ” the flocculus and the paraflocculus, these two divisions have been combined in our table when listing the results of these investigators, in spite of the fact that flocculus and paraflocculus are entirely different structures.

INTRINSIC FIBERS OF THE CEREBELLUM 269
As appears from the table Saito's findings ('22 a, b) differ to some extent from Clarke and Horsley's results as he states that each of the main divisions of the cerebellar cortex are connected with practically all of the nuclei. His lesions, how- ever, rarely are confined to one lobulus and the descriptions do not always permit a satisfactory appraisal of the individual experiments. According to Saito some of the cortico-nuclear fibers cross in the midline (especially from the paramedian lobule and from vermis to the nucleus of Deiters).
Hohman ( '29) on the other hand maintains that all cortico- nuclear fibers pass to the homolateral nuclei and in some cases he gives detailed descriptions of where the fibers from the different cortical subdivisions terminate. His results agree quite well with those obtained by Clarke and Horsley. The same holds for Bender ('32) whose investigations, however, are confined to the connections of the vermis.
Of particular interest are the results of Dow ('36, '38) since his investigations lend strong support to the validity of the new conception of cerebellar morphology which Herrick ( '14, '24) and especially Larsell ('20, '23, '25, '26, '31, '32, '34, '35, '36, '37; Larsell and DOW, '35) arrived at on the basis of comparative-anatomical investigations.
Mussen ('27, '29) and Rasmussen ('32, '33) mention in papers dealing with other topics that most of the fibers from the vermis to the nucleus fastigii are crossed. Rasmussen, furthermore, states that some fibers from the vermis also pass to the vestibular nuclei and that presumably the fibers from the hemisphere to a great extent, if not exclusively, end in the nucleus dentatus and emboliformis.
The reason for the discrepancies in the results obtained by the different investigators which the table demonstrates, must to some extent be sought in the shortcomings of the Narchi-technique. As pointed out by Hohman et al., great caution must be used in appraising Marchi-preparations. Un- doubtedly pseudo-degeneration was frequently mistaken for real (thus by Kuzume, '26, who found practically all the cerebellar divisions to be interconnected). Another possi-

TA
BL
E 1
1 C
ER
MIS
II
HE
MIS
PH
ER
E8
i PI
KIC
C.
I,
Iven
tro-
, ton
of
caud
al p
or-
Nuc
. de
nt.
lat.
ex-
tr
emit
y of
11 L
ob. a
. I P
.~o
cc
. i~-
- ~
1 Lob. h
. ~
Loh
. c,
I Lob. c
2 1
I Fl
oe.-n
od. L
obe
~_
__
_~
.~
.
__
_
_.-_
__
__
-
Cla
rke
and
Hor
aley
, '0
5 ca
t, d
og,
mon
key
Lih
vy.
'16
1
Nuc
. Bec
hter
ew
rabb
it,
guin
ea p
ig
I I
L-.-"---i
Coe
nen.
'21
!I B
rouw
er a
nd
N
uc. f
asti
gii
( ?
) 1.
I
Nuc
. D
eite
rs
Uuc
ves
tib.
IN
uc. f
ast.
' N
uc.
fast
.
part
s of
Nu
t,. B
echt
.I/.
Nuc
. fas
t.
: N
uc. v
esti
b. 1
Nuc
. int
erp.
i
1, (f
ew)
j (fe
w)
~ (f
rom
late
ral
1 i!
'
, py
ram
.)
j ~
I _
_
i I - "---
NU
~. fa
st.
Nuc
. glo
b.
NU
C. D
eite
rs
I 'i /I
Sait
o, '2
2 ra
hbit
1 I I
1, I j 1
post
erio
r ha
lf
of N
uc. f
ast.
Hub
man
. '2
9 Iv
entr
olat
. ca
t
! I-.- B
ende
r, '3
2 ~
1 1
Nuc
. glo
b.
dog
! 1
B'e
wii
bers
to
NU
C.
glob
. and
1
.I
I
I I N
Uc.
emo
li I
Nut
. fas
t.
bu
c.
fast
. N
uc.
glob
. N
uc.
glob
. /IN
uc. d
ent.
I I
i u
__
~
Y
Nuc
. den
t. m
ostl
y la
tera
lly
I N
u'.
em?.
\
litt
le
Nuc
. fa
st.
"I~uc.
emb.
'
1 __
I N
UC
. gl
00.
) N
uc.
dent
. Il dors
o-
1 N
~C
. fa
st.
1 med
ialp
art
1 N
uc. D
eite
rs
I, N
UC
. ue
iter
sl
. _-
ante
rior
1
I NU
~. fast
. id
orso
: )a
t.-
Nuc
. in
terp
. I (
ant.
Nuc
. fas
t.
j N
uc. v
esti
b.
I es
pec.
fro
m
11 ,
linp
ula
, I
thro
ugh
hook
bun
dle
/I
Dow
. '36
ra
t. c
at
now
, '3
8 m
onke
y
I I I
__.
.....
.-
-_
__
~
Nut
. ci
ent.
Nuc
. in
terp
.
dors
o. l
at.
and
lat.
bo
rder
N
uc. d
ent.
of
NU
C. d
ent.
-~
1
Nuc
. de
nt.
, Nuc. i
nter
p.
(les
ion
near
to
the
I ve
rmis
)
I
~
Nuc
. in
terp
.
Nu
c.
intr
rp.
Nuc
. de
nt.

INTRINSIC FIBERS OF THE CEREBELLUM 271
bility for misinterpretation arises when the lesions comprise several lobuli, more particularly so when these lobuli repre- sent quite different structures, as for instance flocculus and paraflocculus.
Our own results mainly confirm the findings of Clarke and Horsley, Hohman and Dow. As will be seen in the general discussion, a critical survey of the literature leads to the conclusions that the diverging results of some investigators are untenable.
MATERTAL AND METHODS
Our experiments were performed on rabbits and cats. Lesions were produced in the different divisions of the cere- bellar cortex by a Graefe knife or thermocautery. Since Dow has published detailed investigations of the connections of the flocculonodular lobe (Larsell) we have deemed it superfluous to include this division in our experiments, and have preferred to concentrate our attention on the corpus cerebelli. The lesions in the anterior lobe in the cat were made after removal of the skull above the occipital lobe and cautiously lifting the latter, the cerebellum being reached through a trephine hole in the bony tentorium. A similar procedure was employed on injuring the rostra1 parts of the lobulus simplex and ansiformis. In rabbits it is difficult to produce lesions in the anterior lobe without simultaneously injuring other divisions of the cerebellum. Our material, at least, does not include rabbits with isolated lesions of the anterior lobe. The opera- tions were made under ether-narcosis, precautions being taken to avoid infections. Afterward the animals were kept alive for 12 to 20 days. They were then killed with chloro- form and the brains removed after perfusion of the head through the carotid arteries with Muller fluid. The procedure in preparing the Marchi series is outlined in the first part of this investigation (Jansen, '33). In all thirty-nine rabbits and thirty-two cats were operated. Of these animals eight

272 J A N J A N S E N A N D A L F BRODAL
rabbits and fourteen cats were selected as suitable for the study of the cortico-nuclear projections.2
Some explanatory remarks on the nomenclature and the diagrams employed are necessary. Bolk’s terminology is used with the exception of his “formatio vermicularis.” In- stead we use the terms flocculus and paraflocculus as the more recent comparative anatomical investigations, especially by Larsell, have shown conclusively that these parts are entirely different structures. This view has furthermore been substantiated by the experimental studies by Dow (’36, ’38). Although we use the terminology of Bolk, occasionally the old terms hemispheres and vermis will be employed, as will also the terms flocculonodular lobe (Larsell) and the corpus cere- belli (Herrick, Larsell).
Whereas in the cerebellum of the cat all subdivisions desig- nated by Bolk are easily identified, this is not the case in the cerebellum of the rabbit. The lobulus ansiformis cannot be subdivided into a crus I and a crus 11. Further, subdivision of lobulus c into a lobulus c1 and cz is hardly possible as long as no sulcus can be convincingly homologized with the sulcus prepyramidalis. The delimitation of the lobulus simplex is a matter of opinion. We have preferred to include only the most rostra1 folium of the lobulus ansiformis under this term. For a more detailed discussion of the points mentioned above and for the nomenclature used, the reader is referred to the atlas of the rabbit’s cerebellum published by Brodal ( ’40 b).
For the cerebellar nuclei we have adopted Brunner’s (’19) terminology, distinguishing between a nucleus medialis or fastigii, a nucleus interpositus (synonyms : nucleus inter- medius, intermediate cell mass) and a nucleus lateralis or dentatus. The intrinsic nuclei of the cerebellum in the cat and the rabbit form a more or less continuous cell mass making any sharp delimitation between the separate nuclei difficult.
* It should be mentioned that series suitable for a study of the cortico-nuclear connections will not always prove equally useful for study of the arcuate fibers, and vice versa. On this account the rabbit material selected for the present investigation differs to some extent from that on which the first paper (’33) was based.

INTRINSIC FIBERS OF T H E CEREBELLUM 273
In particular the boundary between the nucleus interpositus and the nucleus lateralis will to a great extent be arbitrary.
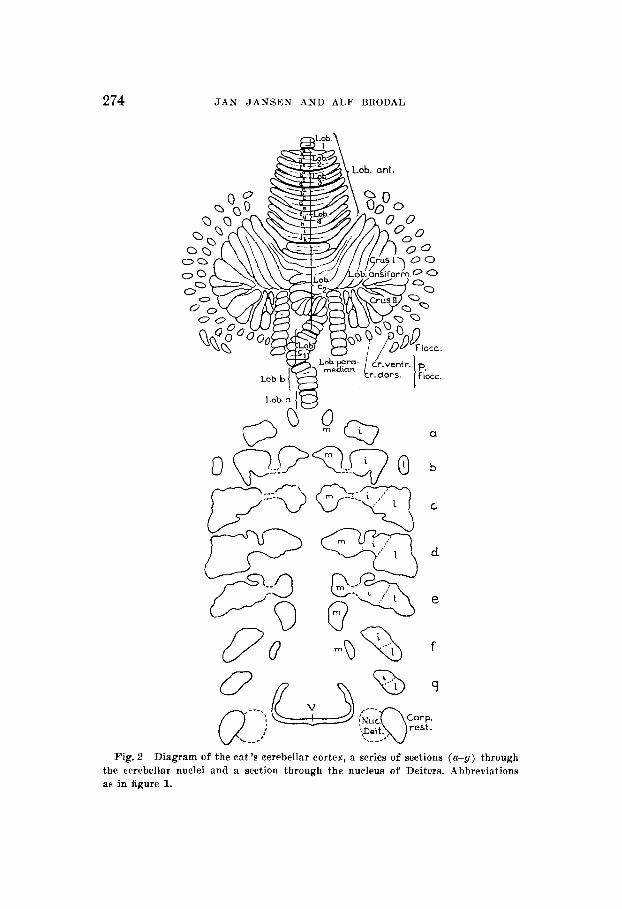
I n an investigation such as the present, however, distinctly defined nuclear boundaries are indispensible for descriptive purposes. The deliminations used in this paper are evident from figures 1 and 2. Like Brunner ('19) we were unable to
Fig. 1 I n the upper half of the figure is a diagram of the cerebellar cortex of the rabbit as unfolded in one plane. The different lobuli and folia are labelled according to Brodal ('40 b). Below is reproduced a series of sections (a-h, (I most rostral) through the cerebellar nuclei and a section through the nucleus of Deiters. Corp. rest = corpus restiforme, i = nucleus interpositus, 1= nucleus lateralis, m = nucleus medialis, nur. Deit. = nucleus of Deiters, v = fourth ventricle.
274 JAN JANSEN A N D ALF BRODAL
a
b
u n
Fig. 2 Diagram of the eat's cerebellar cortex, a series of sections (a-g) through the cerebellar nuclei and a section through the nucleus of Deiters. Abbreviations as in figure 1.

INTRINSIC FIBERS OF THE CEREBELLUM 275
discover distinct cytological differences suitable for a basis for subdivision of the nuclear mass.
Our subdivision rests partly on morphological features, partly on the findings in the present investigation. I n the cat (fig. 2) the nucleus fastigii forms a rather well-defined cell structure, oiily at the intermediate levels (c-d-e, fig. 2) are the boundaries indistinct. Here i t seems appropriate to draw the boundary toward the nucleus interpositus in the middle of the cell bridge which connects the two nuclei. The boundary between the nuclei interpositus and lateralis was drawn chiefly on the basis of the projection areas determined in this investi- gation since a distinct morphological subdivision is found at the rostra1 levels only. At the caudal levels the ventrolateral border of the projection area of the lobulus paramedianus was chosen as the boundary between the two nuclei.
As our rabbit material is incomplete, our subdivision of the nuclear mass in this animal, as indicated in figure 1, must be rather provisional.
I n the upper part of the diagrams the surface of the cere- bellum is presented as unfolded in one plane and the different subdivisions and folia are indicated, below a series of frontal sections through the cerebellar nuclei and a section through the vestibular nucleus are drawn. The diagrams for the rabbit's cerebellum are those published by Brodal ( '40 b).
I n presenting our results we will make extensive use of these diagrams. Solid black indicates the parts of the cortex which have been completely destroyed or undermined, verticals indi- cate more superficially injured cortical areas. The nuclear degenerations are marked by verticals and horizontals where dense, verticals alone where slight degeneration is found.
I n determining the site of the lesion we have made use of photographs of the cerebella, taken after fixation and of comparison of the series with normal ones.
Any one familiar with the Marchi-technique will know the difficulties involved in determining precisely where the de- generating fibers end. I t goes without saying, therefore, that the boundaries of the areas of degeneration indicated in the

276 J A N J A N S E N A N D ALF BRODAL
diagrams necessarily must be approximate. This source of possible error should not be over-emphasized, however. As a matter of fact, in studying our material we were rather struck by the comparative ease with which the areas of terminal intranuclear degeneration could be distinguished. The terminal areas a re characterized by diffusely arranged, rounded, osmiophilic droplets of various size, coarse and fine ones indiscriminately scattered, often giving the area a dust-like appearance, whereas the “non-terminal’ ’ degenera- tion is distinguished by the more irregular shape and the more definite arrangement of the droplets in strands and bundles. We feel convinced therefore that the determination of the areas of degeneration is accurate enough to justify the rather detailed analysis of the cortico-nuclear projection which is presented below.
EXPERIhlENTS ON THE RABBIT
Since we have so fa r not been able to overcome the technical difficulties involved in making isolated cortical lesions on the anterior surface of the rabbit’s cerebellum, the eight series which a re described below unfortunately do not give an exhaustive picture of the cortico-nuclear projection. How- ever, the main features will be brought out and form a useful basis for comparison with the cat.
I.’ e r m is
In our rabbit material two animals only are available for study of the nuclear projection from the vermis. In the other experiments on the vermis the lesions were so small that i t proved impossible to trace the degenerating fibers with certainty.
The lesion may be seen in figure 3, involving mainly the folia 3 to 5 of the lobulus C. Caudally and rostrally, the adjacent folia are slightly injured and to the side the lesion extends slightly into the area medullaris.
From the lesion heavy degenerated fibers in great numbers course ventrad toward the cerebellar nuclei. Here a consider-
Rahbit 39.

INTRINSIC FIBERS OF THE CEREBELLUM 277
able portion of the fibers participate in forming the fibrae perforantes which merely pass through the cerebellar nuclei and enter the medulla where they apparently terminate in the nucleus of Deiters. As far as can be ascertained in this series very few if any fibers go beyond this nucleus.
Another portion of the degenerated fibers from the vermis end bilaterally in the medial cerebellar nucleus as shown in the diagram (fig. 3) . Note the fact that the ventrolateral sec- tion of the medial nucleus at the levels f and g is free of degeneration.
Fig.3 Rabbit. In the diagrams, figures 3 to 23, the cortical lesions are indicated by solid black and verticals (where the cortex is only partially de- stroyed). The nuclear degeneration is marked with verticals and horizontals where intense, with verticals alone where less intense degeneration is present.

278 J A N J A N S E N AND ALF BRODAL
This experiment justifies the statement that the cortex of the lobulus c of the vermis projects on the medial cerebellar nucleus (nuc. fastigii) and the nucleus of Deiters. Since the Iesioii is bilateral the experiment does not answer the question whether the projection is ipsilateral, contralateral or both.
Fig.4 Rabbit 18.
Rabbit 18. The tiny lesion in this experiment involves parts of the three caudalmost folia of lobulus c as shown in figure 4. As appears from the same diagram, the intranuclear degenera- tion is confined to the caudal pole of the medial cerebellar nucleus and the nucleus of Deiters, on both sides.

INTRINSIC FIBERS OF THE CEREBELLUM 279
Paramedian 1 obul e Rabbi t 13. In this experiment the injury is located medially,
involving three folia of the right paramedian lobule. Further- more slight degenerative changes, presumably caused by circu- latory disturbances, are found in one folium of the vermis and in tiny areas of 2 or 3 ansiform folia, as indicated in the diagram.
The distribution of the degenerated cortico-nuclear fibers may be seen in the diagram (fig. 5 ) . The fibers spread over practically the entire rostro-caudal length of the nuclear mass occupying its middle portion. From the lesion the fibers follow two main directions. A smaller portion which apparently origi- nates in the rostro-medial part of the paramedian lobule forms a well defined bundle that is easily traced ventralward in the medullary mass between the vermis and the paramedian lobule. These fibers terminate in the characteristic crest like extension of the nuclear mass which is seen in the diagram at the levels f and g, and in an area lying ventro-medial to this crest, a region which, as we have just seen, did not receive degenerating fibers in the experiments on the vermis (fig. 3) . The majority of the fibers from the paramedian lobule, how- ever, take a more lateral course in the medullary rays of the lobule and give rise to the degeneration which is found a t the more rostral levels of the cerebellar nuclei (a - f , fig. 5). The slight degeneration in the medial nucleus at level h is reason- ably explained by the superficial injury to the cortex of lobulus c.
The results obtained in rabbit 13 may be corroborated by the findings in further experi'ments, a brief mention of which will suffice in this connection.
Rabbi t 12. As appears from the diagram figure 6 the injury is confined to the lateral parts of the four rostral folia of the right paramedian lobule, rostrally touching adjacent parts of the two lateral folia of the ansiform lobule.
The distribution of the degenerated cortico-nuclear fibers conforms in principle to the findings in the preceding experi- ment. The differences in details which are evident on compari- son of the diagrams, are eastly explained by the somewhat different location of the lesions within the cortex.

280 J A N JANSEN AND ALF BRODAL
Our experiments on the paramedian lobule clearly demon- strate that this par t of the cortex is related mainly to the intermediate portion of the nuclear mass, including the “crest”
d
e
9 @ h
Fig.5 Rabbit 13.
0
Fig. 6 Rabbit 12.
and an area ventromedial to it, the latter topographically being a part of the medial nucleus.
Unfortunately we have not been able so far to produce lesions in the rabbit suitable for the study of the cortico-nuclear projections of the ansiform lobule.

INTRINSIC FIBERS OF THE CEREBELLUM 281
Para flocculus This group comprises four rabbits in one of which the lesions
are strictly confined to the paraflocculus, whereas in the three remaining animals the experiments are complicated by more or less extensive injuries to the lobulus paramedianus as well.
Rabbit 20. As appears from the diagram figure 7 the lesion involves parts of 6 or 7 folia of the paraflocculus, the flocculus remaining intact. Degenerated fibers may be traced from the lesion in great numbers in medial direction. For more detailed description of their course see Jansen ( '33! p. 375). The termination of the cortico-nuclear fibers is indicated diagram- matically in figure 7. As will be seen the intranuclear de- generation is located laterally, mainly confined to the dorsal part of the so-called lateral extension of the dentate nucleus.
Rabbit 14. In the diagram figure 8 the lesion is seen to involve parts of four folia in the middle of the paraflocculus, to a lesser degree the surrounding folia and a couple of floc- cular folia. (For details see Jansen, '33, p. 378.) The pro- jection of the injured area on the cerebellar nuclei is likewise indicated in the diagram. If one compares with figure 7 i t becomes evident that the distribution of the degenerated fibers within the nuclei conforms essentially in both instances, the degeneration being confined mainly to the lateral portion of the nuclear mass. It should be noticed, however, that the degeneration in rabbit 14 is more extensive and particularly that it extends farther medial than in the preceding experi- ment. From the flocculus the fibers run to the vestibular nucleus.
Rabbit 38. In this animal small lesions in the lateral region of the paramedian lobule and adjacent folia of the para- flocculus are responsible for degenerations in the lateral and intermediate nuclei (fig. 9) . The picture is in good agreement with what might be expected on the basis of the experiments described above.
Rabbit 31. In this experiment (fig. 10) parts of practically all parafloccular and floccular folia are injured in addition to a circumscribed lesion of the paramedian lobule.
A glance a t the diagram figure 10 makes clear that the distribution of the degenerated cortico-nuclear fibers (from the paraflocculus) shows great similarity to the condition found in rabbit 14, the main difference being a more massive

282 JAN JANSEN AND ALF BEODAL
degeneration at the rostra1 nuclear levels in the latter animal and more degeneration a t level g in the present experimnt.
The degenerated fibers from the flocculus pass medially in two parallel bundles, one curving around the dorsal aspect of the restiform body, the other passing along the ventral surface of the cerebellar riaclei before turning ventrad toward the superior and dorsal vestibular nuclei where the fibers seem to end. The fibers of the first mentioned bundle seem to terminate mainly in the nucleus of Deiters, to a smaller extent in the other vestibular nuclei. There can be little
Fig. 7 Rabbit S O . Fig. 8 Rabbit 14.

INTRINSIC FIBERS OF THE CEREBELLUM 283
doubt that the majority of the efferent fibers from the floc- culus end in the vestibular nuclei. However, the present series also indicates that some of the floccular fibers terminate, or perhaps give off collaterals, in a small group of cells located in the dorsal bundle, immediately ventrolateral to the cere- bellar nuclear mass.
It appears from these experiments that the paraflocculus projects on the lateral division of the cerebellar nuclear mass, i.e., the paraflocculus is related to the dentate nucleus. It
Fig9 Rabbit 38. Fig. 10 Rabbit 31.

284 JAN JANSEN A N D ALF BRODAL
was not possible in our material to determine the presence of possible cortico-vestibular connections.
I t is furthermore evident that the floccular cortex is pro- jected on the vestibular nuclei, probably on all of them, but preferably on the nuclei of Deiters and Bechterew. There can be little doubt that a great majority of the cortico-nuclear fibers from the flocculus pass to these nuclei. Particularly the findings in rabbit 31, however, where the floccular degenera- tion is fairly extensive, tend to show that some of the floccular fibers a re related to small groups of cells, lying within the fiber bundles in the space between the cerebellar nuclei and the dorsal aspect of the restiform body.
EXPERIMENTS ON THE CAT
Vermis
We will first consider four experiments shedding light on the cortico-nuclear projection of the vermis.
Cat 29. Small strictly superficial lesion in the midline involviiig three folia (c-d-e) of the lobulus 4 of Bolk (fig. 11). Owing to the small lesion the number of degenerated fibers is moderate, but they can be traced without difficulty toward the cerebellar nuclei. Part of the fibers are seen to terminate in the anterior half of the medial cerebellar nucleus on either side as indicated in figure 11. Another portion of the fibers merely penetrate the nuclear mass and run ventrad into the medulla oblongata where the majority end within the nucleus of Deiters. Some of the fibers seem to pass into the superior nucleus and a few may even enter the reticular formation.
Cat 7. I n this experiment the injury is quite extensive but confined to the verrnis, as appears from the diagram (fig. 12). Practically all folia of the lobulus c2 of Bolk a re involved and in addition the caudal, superficial folia of the lobulus 4. As shown in figure 12 degeneration is found almost through- out the entire length of the medial nucleus on both sides. I n addition a considerable number of degenerated fibers are found within the intermediate nucleus forming bundles with a somewhat irregular dorso-ventral direction. Even though it cannot be definitely stated that none of these fibers terminate within the nucleus interpositus, all the indications are that

INTRINSIC FIBERS OF THE CEREBELLUM 285
U
Fig. 12 Cat 7.

286 JAN JANSEN AND ALF BRODAL
they merely pass through the nucleus forming fibrae perfo- rantes which may be traced into the medulla. Here they a re lost in the vicinity of Deiters’ nucleus where most of the fibers apparently end. Some fibers take a medial direction toward the superior vestibular nucleus, others seem to enter the reticular formation, but it is impossible to determine exactly where these fibers terminate.
Cat 11. A lesion was produced with thermocautery in the vermis involving parts of 10 folia of lobulus el and c2 (fig. 13). As indicated in the diagram the degenerating cortico- nuclear fibers were traced to the caudal half of the medial cerebellar nucleus on both sides. It is difficult to decide to what extent fibers possibly pass beyond this nucleus and enter the medulla oblongata, the picture being obscured by pseudo- degeneration within the fibrae perforantes.
Cat 1. Here again the lesion is small and strictly confined to the cortex, but situated considerably farther caudal in the vermis, involving 3 folia of the lobulus c2 of Bolk. The vast majority of the degenerated fibers course ventralward and can easily be traced into the caudal par t of the medial cerebellar nucleus where they terminate on both sides cor- responding to the levels f and g of the diagram. Whether all fibers terminate here remains an open question in this case. The indications a re that some fibers penetrate the nuclei and pass farther ventrad into the medulla oblongata, but the evidence is not conclusive.
These four experiments on the vermis clearly show that this par t of the cerebellar cortex preponderantly projects on the medial cerebellar nucleus. The findings in cats 1 and 29 where the lesions a re restricted to small cortical area, further- more indicate that the anterior lobe portion of the vermis is related to the rostra1 levels of the nucleus, whereas the caudaI part of the vermis is connected with the caudal part of the nucleus.
In addition to projecting on the medial cerebellar nuclei, the cortex of the lobus anterior, probably also the cortex of lobulus c is connected with the vestibular nuclei, chiefly the nucleus of Deiters. Some fibers, however, also seem to be related to the superior vestibular nucleus and possibly the reticular format ion.

INTRINSIC FIBERS OF THE CEREBELLUM 287
,-
B
Fig. 13 Cat 11. The " ?" in the vestibular nuclei indicates uncertaiii cortico- vestibular connections.

288 JAN JANSEN AND ALF BRODAL
Lateral part of anterior lobe
Having ascertained that the median part of the cerebellar cortex (vermis) projects on the medial cerebellar nucleus and the vestibular nuclei we now turn our attention to the projections of the hemispheral parts. It seems natural to begin with the lateral division of the anterior lobe.
Cat 27. The lesion is located in lobulus 3 and 4 of Bolk, destroying the cortex over the lateral parts of 6 to 7 folia, as appears from the diagram in figure 14. The microscopic examination furthermore revealed a slight (presumably vascu- lar) , disturbance of the cortex in a couple of folia of the crus I (shaded in the diagram) causing degeneration of scat- tered neurons in this region. A glance at the diagram shows that the cortex of the injured area projects on the nucleus interpositus where the degeneration is found particularly dorsomedially in the rostra1 half of the nucleus. There can be little doubt that this represents the nuclear area of the injured lateral part of the anterior lobe. There is also some degeneration within the homolateral medial nucleus, a fact which is most reasonably explained by the overlapping of the lesion on the median (vermal) part of the anterior lobe.
A slight degeneration which is encountered rostrally in the dorsolateral angle of the lateral nucleus is accounted for by the superficial lesion of crus I (vide irifra cat 12, fig. 16).
In our opinion this experiment shows that the lateral parts of the anterior lobe mainly project on the nucleus inter- positus. In other experiments we have never seen degenera- tion within this part of the nucleus interpositus except when the lateral part of the anterior lobe was injured. Beside the fibers which terminate in this nucleus there are quite a few, however, which join the fibrae perforantes and enter the medulla oblongata. The majority of these fibers seem to end in the nucleus of Deiters. Some fibers, however, were traced to the dorsal aspect of the nucleus vestibularis superior. It is not quite excluded that these fibers originate in the vermis portion of lobus anterior.
The conclusions drawn from this experiment find further substantiation in cat 30.

I N T R I N S I C FIBERS OF T H E CEREBELLUM 289
Fig. 14 Cat 27. Fig. 15 Cat 30. The “ a ’ in the cerebellar nuclei at level e indicates a defect in the series at this level.

290 J A N J A N S E N A N D ALF BRODAL
Cut 30. A small lesion was produced with tliermocautery ill the lobus anterior involving the lateral part of the lobulus 4 of Bolk (folia c - A ) . As appears from the diagram (fig. 15) ill addition small areas of 3 or 4 adjacent folia of the lobulus simplex and the ansiform lobule are also injured. The lesion in this case is rather deep, touching the central white matter in a restricted area. As in the preceding experiment the intranuclear degeneration is found in the rostral half of the nuclear mass located (see diagram) in adjacent areas of the interniediate and lateral nuclei. A comparison with cat 27 (fig. 14) makes clear that the distributioii of the degeneration within the nucleus interpositus in the two experiments coincides to a reasonable extent in view of the somewhat different locali- zation of the lesion within the anterior lobe in the two animals. The assumption lies near at haiid, therefore, that the small injuries in the lobuli simplex and ansiformis a re responsible for the degeneration within the lateral cerebellar nucleus. Before describing the next experiment which lends strong support to such a conclusion, we call attention to a slight and questionable degeneration within the medial cerebellar iiucleus on both sides. It was difficult to decide whether this was a pseudo-reaction or not. IJnder all circumstances the number of degenerated fibers would be negligible compared with those entering the nucleus interpositus.
A us if o rwi I (I 0 111 e
On the basis of the two preceding experiments the conelusion is drawn that the lateral (liemispheral) parts of the anterior lobe project on the rostral half of the nucleus interpositus and to a smaller extent possibly on the vestibular nuclei (mainly the nucleus of Deiters). This being so, the degeiiera- tion which occurs in the lateral cerebellar nucleus in cat 30 should be due to the lesion in lobuli simplex aiid ansiforniis. The experiment to the described next strongly favors this conclusion.
The preceding experiments shed some light 011 the projection of the rostral parts of the ansiform lobe. We shall now turn to cat 12.
Cat 12. In this experiment most of the lesion is situated within crus 11, involving the major parts of three folia,

INTRINSIC FIBERS OF THE CEREBELLUM 291
ventrolaterally slightly encroaching on the neighboring folia of the parafloccular lobe (fig. 16). A glance at the diagram shows that degenerating fibers enter the lateral cerebellar nucleus' and terminate in sections of the nucleus which were virtually free of degeneration in the experiments described above. The degeneration is mainly present rostrally in the dorsal parts of the nucleus, extending particularly far caudal- ward along the medial border of the lateral nucleus, here occupying well-defined areas a t the levels d and e , but disap- pearing at level f . Other degenerating fibers enter the nucleus more ventrally and form a narrow zone of degeneration along its ventrolateral aspect. It is evident from experiments to be described below that this latter degeneration is related to the slight injury inflicted upon folia of the parafloccular lobule.
Thus this experiment strongly suggests that the cortex of crus I1 projects on the dorsomedial parts of the lateral cere- bellar nucleus.
Paraflocculus
Before entering on a more detailed discussion of the pro- jection of the ansiform lobe it seems practical to describe an experiment where the lesion is largely confined to the para- floccular lobule.
Cat 8. The left hemisphere was injured with thermocautery, interrupting the fibers from the area which is marked in the diagram (fig. 17) . As may be seen the lesion comprises some 8 or 10 folia of the parafloccular lobe, dorsally affecting adjacent folia of the ansiform lobule and ventrally affecting the flocculus. The point of the instrument has even reached beyond the flocculus and made its way into the acoustic nerve, severing a great number of the fibers near their entrance into the medulla. Furthermore it appears from the diagram that the mass of degeneration is found in the lateral cerebellar nucleus, occupying to a great extent areas which in the experiments hitherto described were free of degeneration, viz., the ventrolateral parts of the nucleus. If we now com- pare with cat 12 (fig. 16) it seems reasonable to regard the degeneration in the dorsolateral parts of the nucleus (marked a in fig. 16) as a consequence of the injury in the lateral part

d
e
Fig. 16 Cat 13. The I ' ?' ' in the lateral cerebellar nucleus at level b indicates a defect in the series at this level. a, referred to in the text.
99"
Fig. 1 7 Cat 8. a, b , c referred t o in the text.

INTRINSIC FIBERS OF THE CBREBELLUM 293
of the ansiform lobule. Disregarding for the moment this degeneration we find then that the parafloccular lobe is con- nected with the ventrolateral parts of the lateral cerebellar nucleus. Of course, there is still the lesion of the floccular folia to be accounted for. Little difficulty is encountered, however, in distinguishing between the parafloccular and floccular projections since the fibers from the latter form a well defined layer along the ventral surface of the flocculus, well separated from the parafloccular fibers by a double cortical layer. The parafloccular fibers may be seen running in medial direction into the ventrolateral section of the lateral cerebellar nucleus, whereas the floccular fibers form a bundle which courses mediad between the nucleus and the roof of the IV ventricle.
Where the restiform body and the cerebellum fuse, acoustic fibers join the “floccular” bundle. The vast majority of fibers from both sources pass ventromediad toward the vestibular nuclei where they apparently end. Some fibers, however, leave the bundle and turn dorsad. The origin and ultimate destina- tion of these fibers remain obscure.
The present experiment seems to warrant the conclusion that the parafloccular cortex projects on the ventrolateral part of the lateral cerebellar nucleus. Furthermore i t is apparent from this series that the nuclear connections of the floccular cortex are largely-if not entirely-with the vestibular nuclei.
These conclusions are further substantiated in cat 31.
Cut 31. A superficial lesion was made in the right cerebellar hemisphere with thermocautery. From the microscopic ex- amination it appears that five or six folia of the paraflocculus were injured, together with parts of adjacent ansiform and floccular folia as indicated in the diagram (fig. 18). Here, as in cat 8 (fig. 17), the nuclear degeneration is confined to the lateral cerebellar nucleus and a comparison between the two cases clearly brings out the fundamental conformity in the distribution of the degeneration in both instances. The fibers issuing from the lesion in the ansiform lobule can be traced without difficulty to the rostra1 nuclear levels ( b - c - d ) where they end in the dorsolateral parts of the lateral nucleus, whereas the fibers in the caudal half of the nucleus are clearly derived from the paraflocculus. The floccular lesion is so small

294 J A N JANSEN A N D ALF BEODAL
as to render it impossible to trace the corticofugal fibers to their terminal nuclei with reasonable accuracy.
The series supports the conclusions drawn from the fore- going experiments, viz., that the ansiform lobule projects on the dorsolateral part of the lateral cerebellar nucleus, and the paraflocculus on the ventrolateral part.
Crus ZZ of ansi form lobzile
Having now determined the relations of the paraflocculus, we may return to the anso-paramedian lobe. It still remains to demonstrate the projections of the caudal par t of the crus I1 and the paramedian lobule. We may begin with cat 18.
Cat 18. The connections of practically all folia of the crus I1 were severed with a Graefe knife. In addition neighboring folia of crus I, paraflocculus and the paramedian lobule were injured, as indicated in the diagram (fig. 19). The distribution of the degeneration within the cerebellar nuclei may be seen in the same diagram. The degeneration is confined to the intermediate and lateral cerebellar nuclei, distributed in such a way that it is posssible to distinguish three different areas (a , b , c). Let us a t once call attention to the fundamental correspondence between area a and the similarly labelled de- generation in cat 12 (fig. 16). It is reasonable to conclude that we are here dealing with fibers from the lesion in the ansiform lobule. In fact the fibers may be traced from the lesion into the lateral nucleus. The more extensive degenera- tion a t level d of the diagram as compared with cat 12 is easily explained by the more extensive lesion of crus I1 in the present experiment. This might also be the explanation for the degeneration in area 0. I t appears, however, that fibers to this area may be traced from the paraflocculus, thus rendering it highly probable that this part of the lateral nucleus is related to the paraflocculus. The degeneration ( c ) at the nuclear levels f and g still remains to be accounted for. I t seems natural to conclude that the lesion of the para- median lobule is responsible for this degeneration, but, since the lesion affects several lobuli in this experiment, we admit that conclusions must be drawn with reservations. The de- ductions suggested above, however, are strongly supported by the findings in experiments to he described presently.

INTRINSIC FIBERS O F THE CEREBELLUM 295
Fig. 18 Cat 31. Fig. 19 Cat 18. a, 6, o referred to in the text.

296 J A N J A N S E N A N D ALF BRODAL
The next experiment makes i t possible to define more ac- curately the projection area of the caudomedial parts of the crus 11.
Cat 5 . The lesion is almost entirely confined to the crus I1 of the aiisiform lobule involving its caudomedial folia, only slightly affecting adjacent parts of the paramedian and para- floccular lobuli (fig. 20). The nuclear degeneration is found in the caudal half of the cerebellar nuclear mass. Unfortunately it is not possible to trace the degenerating fibers from the affected folia of the paramedian lobe and paraflocculus sepa- rately. In view of their comparatively small number, however, no great error is committed when we disregard these fibers for the moment and consider the nuclear degeneration in figure 20 as roughly indicating the area of projection of the injured folia of crus 11. This par t of the e m s I1 clearly pro- jects on the dorsolateral sections of the cerebellar nuclear mass.
Let us stop here for a moment and review the experiments bearing on the projection of the ansiform lobe. If the figures 15, 16, 29 and 20 are compared, i t is at once evident that the cortex of the ansiform lobule projects on the lateral half of the cerebellar nuclear mass. In figure 21 the areas of nuclear degeneration, which according to the analysis presented above are related to the lesions of the ansiform lobule, a re super- imposed in the same diagram.
I t appears that the projection of the ansiform lobule covers practically the entire lateral cerebellar nucleus, leaving only the veiitrolateral corner and a caudomedial section free. A glance at figure 15 makes clear that the rostromedial portion of the ansiform lobule (including the lobulus simplex) evi- dently is connected with the rostromedial section of the ansiform nuclear field, whereas the lateral folia (fig. 16) predominantly are related to the dorsolateral section and the posteromedial folia of crus 11 (fig. 20) to more caudomedial sections of the lateral nucleus.
Paramedian lobule The projection of the paramedian lobule still remains.
With this objective in mind we turn to cat 13.

Fig.20 Cat 5. The broken line a t level e of the cerebel- lar nuclei marks the border of a defective impregnation in the upper right angle of the nucleus a t this level.’
~ -~
The most distal area of degeneration in figure 20, in Deiter’s nucleus, is not mentioned in the text and does not fit other experimental studies on the cerebellum. The attention of the authors was called to the discrepancy, but because of the tragic conditions in Norway no reply has been received. The figure is published a s submitted, with the editorial notation that the area should be questioned.
297

a
Fig. 2 1 In the left half of the diagram the total extent of the lesions in the experiments 011 the ansiforin lobule (cats 13, 12, 18 and 5 ) and the consequent nuclear degeneration are shown. The right half of the diagram demonstrates the total extension of all lesions and the ensuing nuclear degeneration found in t h e experiments on cats which are reported in this iiivrstigation.
298

I N T R I N S I C FIBERS OF T H E CEREBELLUM 299
Cat 13. I n this experiment the brain was injured with a Graefe knife. As indicated in the diagram (fig. 22) the lesion is partly located in the right paramedian lobule where the connections of the medial halves of 5 or 6 folia have been severed. Rostrally the lesion extends into the transitional zone between vermis on one side, crus I and the lateral division of the lobulus simplex on the other, destroying parts of three folia as shown in the diagram. The tips of some eight folia of the vermis a re cut medially.
The nuclear degeneration is located in the medial and inter- mediate parts of the nuclear mass. From experiments de- scribed above we know that the cortex of the vermis projects on the medial nucleus and i t is natural therefore to regard the degeneration in the medial nucleus as a consequence of the injury to the vermis. The degeneration found in the intermediate divisions of the nuclear mass, should conse- quently represent the nuclear fields of projection of the injured parts of the paramedian and ansiform cortex. In order to differentiate the nuclear fields of these two cortical areas we may trace the course of the degenerating fibers from the lesion.
It is then found that the fibers from the paramedian lobule generally occupy a more caudal and lateral position than the fibers from the rostra1 (ansiform) division of the lesion and the indications a re that the fibers maintain this topographic relation at their points of determination within the nuclei.
This being so it seems reasonable to conclude that the areas of degeneration which are labelled b in the diagram (fig. 22) are related to the ansiform cortex, whereas the areas labelled a are connected with the paramedian lobule. The cortex of the paramedian lobule, in other words, projects on the caudomedial parts of the fused lateral and intermediate cerebellar nuclei, a conclusion which finds further substantia- tion in the next experiment. Before describing the last experiment to be dealt with in this communication, i t should be mentioned that quite a few degenerating fibers in cat 13 join the fibrae perforantes and enter the medulla oblongata. It is not possible, however, to ascertain either their origin or termination, but the possibility remains that the paramedian lobule also sends fibers to the medulla oblongata.

300 J.4N J A N S E N A N D ALF RRODAL
Cat 3. A small cortical lesion was made on the left side, involving parts of 4 paramedian folia as well as two acl- jacent folia of the crus I1 (fig. 23). In conformity with the small lesion the nuclear degeneration is not so extensive as in the preceding experiment (fig. 22). The great majority of degenerating fibers terminate in the middle of the caudal half of the cerebellar nuclear mass. In addition some fibers, most of them apparently originating in the cortex of crus 11, pass rostrally along the outer surface of the lateral nucleus and then turn ventrally as it seems into the medulla oblongata. Their exact termination could not be ascertained.
Let us now compare the diagrams of cats 3 and 13 (figs. 23 and 22). As far as the lesions a re concerned they have the common feature that rostral parts of the paramedian cortex are injured. In cat 13, however, the lesion extends rostral- ward into the ansiform lobule, whereas in cat 3 there is a tiny lateral extension of the lesion into crus 11. Comparing the nuclear degenerations, it appears that both cases have in common the areas of degeneration which are labelled a. Thus the conclusion suggested above, viz., that these areas indicate the nuclear projection of the rostral folia of the paramedian lobule, seems justified. I n this connection we may also call attention to cat 18, figure 19. Here also, where part of the paramedian lobule is injured, we find nuclear degeneration within the same areas.
On the basis of the twelve experiments described above we have tried to make a chart of the cortico-nuclear connections. In doing so we are aware of the difficulties involved. Thus, although the diagrams were made as accurate as possible the boundaries between the different nuclear fields cannot be more than approximate, if onlp because the Marchi method does not permit exact determination of the areas where the fiber tracts end. On the other hand, we feel convinced that whatever inaccuracies may exist, none of them is of funda- mental significance. I n order to make clear to what extent our diagram is based on actual observations we have on the right side of the diagram (fig. 21) indicated the total extent of the twelve lesions in experiments on cat and the nuclear degenera-

INTRINSIC FIBERS OF THE CEREBELLUM 301
Fig. 22 the text.
Cat 13, a, b referred to in F i g . 2 3 Cat 3. The “ 8 ” in the vestibular nucleus indicates an un. certain cortico-vestibular connection. a referred to in the text.

302 J A N J A N S E N A N D ALF BRODAL
tion accompanying them. The findings in these fourteen series were in such good agreement that, in spite of the sources of possible errors pointed out above, we feel that the diagram presented in figure 24 gives a reliable picture of the funda- mental relations between cerebellar cortex and cerebellar nuclei.
DISCUSSION
Before discussing the observations made in this investiga- tion a few words should be devoted to the question of the sub- division of the cerebellar nuclei. From the diagrams it is apparent that we have not found sufficient reason to dis- tinguish a s separate parts a nucleus globosus and a nucleus emboliformis as Rasmussen ('32, '33) and some other authors do. In the cat we have considered the nucleus globusus of Rasmussen a s part of the nucleus fastigii and the cell group named nucleus emboliformis by him is included in our nucleus interpositus. Mussen ('27, '29) uses the same subdivision as Rasmussen. Allen ('24), on the other hand, working with guinea pigs, adopted Brunner's subdivision, as do Hohman and Dow. Clarke and Horsley do not consider the nuclei separately, but from their figure 2 i t appears that they use the name nucleus fastigii in the same sense as i t is employed by us. The nucleus globosus of these authors apparently cor- responds to nucleus interpositus. These differences in nomen- clature should be kept in mind when discussing the cerebellar nuclei and their connections since a great deal of confusion has been caused by the terminology.
A comparison between the results obtained in the cat and in the rabbit reveals a very good agreement in the principal arrangement of the cortico-nuclear projection. Thus in both animals the flocculus projects on the nucleus of Deiters, the lobulus c is related to the nucleus fastigii as well as the nucleus of Deiters and the paraflocculus is projected on the lateral par t of the nucleus dentatus. Our material does not give a clear cut picture of the projection of the lobulus ansi- formis in the rabbit, but the nuclear area presumably related to the ansiform cortex is considerably smaller than the cor-

INTRINSIC FIBERS O F THE CEREBELLUM 303
responding area in the cat, in harmony with the much higher development of the ansiform lobule in the latter animal. The primitive development of the ansiform lobule in the rabbit com- bined with a comparatively high development of the para- median lobule reasonably explains why the nuclear projection area of the latter lobule extends relatively farther rostralward in the rabbit (compare rabbit 13, fig. 5, with the diagram of the cat, fig. 24).
It may be noticed in the rabbit as well as in the cat that the nucleus interpositus has a dorsomedial crest-like extension (a t the levels f and g in the rabbit, d and e in the cat) . I n the rabbit this crest and its ventromedial extension is related to an unusually ~lrell defined bundle whose fibers originate in the cortex of the paramedian lobule. I n the cat the relations of this crest are not quite clear, but as f a r as our material goes it does not seem to be related to the cortex of the paramedian lobule. Thus these crests in spite of their morphological similarity in the two animals may not represent homologous structures .
Turning now to a comparison of our results with those obtained by previous investigators, we may begin with the projection of the vermis. Our own results a r e summarized in the diagram (fig. 24). The findings of previous investi- gators are assembled in table 1. The diagram shows that the nuclear projection field of the vermis is confined to the medial cerebellar nucleus and the nucleus of Deiters.
Let us consider the different lobules of the vermis indi- vidually, first the anterior lobe. Opinions are still divided as concerns the subdivision of the lobus anterior in a median vermis and lateral or “hemispheral” parts. On account of their different connections, however, we have deemed it ap- propriate to distinguish these two parts in the lobus anterior. The vermal region according to our determination extends as f a r laterally as the boundary between the vermis and the hemi- spheres behind the fissura prima. As is apparent from table 1 all authors who have investigated the question (Clarke and Horslep, Saito, Hohman, Bender) agree in finding fibers from

304 J A N JANSEN A N D ALF BRODAL
Fig. 81 Diagram of the eortico-nuclear projection. The niutunllj- interconnected divisions of cerebellar cortex and cerebellar nuclei are marked with identical symbols.

INTRINSIC FIBERS O F THE CEREBELLUM 305
the lobus anterior to the nucleus fastigii. Hohman, Bender and Saito, furthermore, like ourselves have traced fibers from the vermis of this lobus to the vestibular nuclei. When Clarke and Horsley as well as Hohman in addition describe fibers to the nucleus interpositus, the explanation is to be found in the fact that the lesions in some of their experiments extended so far laterally as to include the “heniispheral” part of the lobus anterior (see Hohmaii’s figs. 163 and 164). It is from this lateral part of the anterior 1obe.that fibers pass to the intermediate nucleus as shown in our own experiments. Thus in reality full agreement exists between our results and those of previous workers.
However, Clarke and Horsley as well as Saito also mention fibers which pass from the lateral parts of the anterior lobe to the nucleus dentatus. The rabbit and cat material employed in the present investigation is inadequate with regard to the connections of the extreme lateral parts of the anterior lobe. In monkey material, however, which is now in preparation it is clearly seen that the lobus anterior on the basis of its efferent connections can be divided in three longitudinal zones, viz., a median (vermis) projecting on the nucleus fastigii, an intermediate related to the nucleus interpositus and an ex- treme lateral zone which is connected with the dentate nu- cleus. The fibers observed by Clarke and Horsley and Saito presumably are those originating from the extreme lateral zone of the anterior lobe. Whether this extreme lateral division is a constant part of all mammalian cerebella is a question which calls for further investigations. Studies of the olivo-cerebellar connections by Brodal ( ’40 a ) give some information concerning this point as will be mentioned below.
As regards the lobulus c (Bolk) all investigators agree (see table 1) that it projects on the nucleus fastigii. Some authors also find fibers to the nucleus interpositus and even to the nucleus dentatus, e.g., Clarke and Horsley. However, in those experiments where these authors describe such fibers, the lesion was never confined to the vermis, but other regions were affected simultaneously, their cortex being at least

306 J A N JANSEN AND ALF BRODAL
“bruised” (eg. , cat 11). Where the vermis alone was in- jured the efferent fibers were followed to the nucleus fastigii alone (cats 10, 13, 14, monkey 12). Similar objections may apparently be made to Bender’s findings. Dow (’36) states that the fibers which he has traced to the nucleus interpositus have their origin in the lateral part of the pyramis, in a region which seems to belong to the copula pyramidis. Saito too describes fibers from “vermis” to the nucleus globosus, but they are less numerous than the fibers to the nucleus fastigii, and from his very schematic drawings it cannot be definitely made out whether his nucleus globosus is really the nucleus interpositus. As fa r as can be seen, no findings in the litera- ture conclusively demonstrate any connection from tlie lobulus c to the nucleus interpositus.
As regards connections to the vestibular nuclei the evi- dence is not equally clear cut, but our material, as far as it goes, seems to indicate that the entire vermis besides project- ing on the nucleus fastigii, is connected with the vestibular nucleus (Deiters).
All authors agree that the lobulus ansiformis sends ef- ferents to the nucleus lateralis or dentatus, as also found by us, our findings a re in complete accordance with Hohman’s results. Some autliors, however, describe fibers from the lobulus ansiformis also to other parts of the nuclear complex-. Thus Clarke ancl Horsley find fibers from crus I to the nucleus fastigii. However, in those espei-iments where such fibers are described and tlie verrnis is not simuItaneouslp injured, the authors speak of “few, fine fibei-S” (cats 1 and 4), and the lesions in these experiments nearly reached the vermis. Their finding, therefore, cannot be accepted as conclusi~e. Saito who reports similar findings has 110 series where the lesion is confined to the lobulus ansiformis, and it is scarcely possible from liis paper to get an impression as to the validity of his statements. Dow (’36) describes fibers from “the more ante- rior portion of Crus I and lobulus simplex” in the cat to the nucleus interpositus. This observation is in harmony with our own findings provided the lesion was located medially

INTRINSIC FIBERS OF THE CEREBELLUM 307
in the ansiform lobule where our experiments have disclosed a “paravermian” zone, the cortex of which is projected on the intermediate cerebellar nucleus (fig. 24). Hohman describes fibers from the lobulus simplex to the nucleus fastigii. His experiment cat 5 leaves no doubt that these fibers originate in the vermis portion of the lobule (a par t of lobulus c2).
Our results concerning the efferent fibers from the lobulus paramedianus also are in conformity with those of Clarke and Horsley and Hohman. However, Clarke and Horsley have no experiment in which the lobulus paramedianus alone was injured, but in those cases where this lobulus was affected together with the vermis, they describe fibers to the nucleus “globosus,” i.e., nucleus interpositus. The same is the case with some of Saito’s experiments. Dow ( ’38) describes in the monkey, besides the connection with nucleus interpositus, also fibers to the nucleus dentatus. Since in his experiment there is also a lesion of crus I1 we are inclined to regard the de- generation within the dentate as a result of the latter lesion. The discrepancies may also, however, be due to slight differ- ences in the delimitation of the nuclei.
Concerning the projection of the parafloccular cortex our findings a re in complete accordance with Hohman who states that “the paraflocculus efferents terminate on the ventro- lateral border of the dentate.” Dow (’36) in addition men- tions fibers to the lateral extremity of the nucleus interpositus, where the nucleus is continuous with the dentatus. The dis- crepancy on this point may be entirely due to differences in the delimitation of the two nuclei which, as mentioned above, to some extent must be a matter of opinion. As concerns the other investigators who have studied the connections of the paraflocculus, the objection may be raised against all of them, that they have not properly distinguished between the flocculus and the paraflocculus. The statement by Clarke and Horsley, that the paraflocculus is in connection with the nucleus fastigii is hardly valid. In their experiment cat 4, where half of the paraflocculus was damaged (besides other parts) the authors describe O I A ~ ; ~ “very few fine fibers” to the nucleus fastigii.

308 J A N JANSEN A N D ALF BRODAL
In another experiment (cat 1) the fibers were also few, in the third experiment with lesion of the paraflocculus together with other parts (cat 3), “fine and coarse fibers” to the ventral surface of the nucleus fastigii a re mentioned. How- ever, the lesion in this case consisted in only “slight bruise” of the paraflocculus and the flocculus, and part of the crus 11, and it is astonishing that no fibers were traced to the nucleus dentatus. The somewhat conflicting evidence in Clarke and Horsley’s own material makes it highly improbable that the fibers to the nucleus fastigii originate in the paraflocculus.
That part of the cerebellum of the rabbit which Brouwer and Coenen label flocculus, obviously is the paraflocculus as appears from their figure 1. Very probably also the flocculus was involved in their experiments. From lesions of this type the authors find fibers which probably end in the nucleus fastigii. That they did not find any fibers to the nucleus dentatus in such cases is explained by the extent of the lesion, which also affects the lateral par t of the nucleus dentatus. Similar objections can be raised against the investigations of Lowy. The fibers which Lowy traces to the nucleus of Bechterew probably do not have their origin in the para- floccuIus, possibly they come from the flocculus. If this be the case, it is in full accordance with the results of DOW, who has found fibers from the flocculus to the nuclei of Deiters and Bechterew. Our own material is scanty on this point, but as f a r as can be seen our series corroborate the findings of DOW, as fibers can be followed from the flocculus to the vestibu- lar nuclei.
I t was stated above that extensive connections exist be- tween the vermis of the anterior lobe and the vestibular nuclei. Dow has demonstrated similar relations for the lobuli a and b and our own material leaves little doubt that the same holds for the lobulus G . But great uncertainty prevails when it comes to the question of cortico-vestibular connections from the remainder of the cerebellum. I n several of our experiments involving “extravermian” areas we observed fibers which seemed to traverse the nuclei and join the fibrae perforantes.

INTRINSIC FIBERS OF THE CEREBELLUM 309
Since the fibrae perforantes, however, apparently is a favorite place for pseudodegeneration it is difficult to make certain that we are in these experiments dealing with true degenerated cortico-vestibular fibers. On this account we prefer not to draw any definite conclusion on this point, but leave the question open for further investigation. As is apparent from table 1 Saito also describes such fibers. Clarke and Horsley saw them only in more extensive lesions, and do not regard theni as being of importance.
Before finishing this discussion of the literature we may briefly deal with the question whether cortico-nuclear fibers cross the mesial plane. Clarke and Horsley could find no satisfactory evidence of heterolateral projection and Hohman emphasizes the ipsilateral character of the projection. Saito, Mussen and Rasmussen on the other hand maintain that the cortico-nuclear fibers in part also pass to the heterolateral nuclei. Our own findings are in accordance with the coii- elusions drawn by Clarke and Horsley and Hohman. The cerebellar cortico-nuclear projections seem to be strictly ipsilateral.
Excepting the vestibular connections, an analysis of the data available from the literature shows on practically all points a very good conformity between the experiments of previous investigators and those made by us. The existing discrepancies are more apparent than real, due partly to differences in nomenclature or a different delimitation of lobuli and nuclei. Sometimes pseudo-degeneration seems to have been interpreted as real degenerated fibers. Upon the whole it thus seems appropriate from our investigations and from those of previous authors to draw the following con- clusion : There exists within the cortico-nuclear connections of the cerebellum a definite topographical localizatory corre- lation, every part of the cerebellar cortex being projected on a certain circumscribed portion of the ipsilateral nuclear complex. This cortico-nuclear localization is diagrammatically represented in figure 24.4
' A s concerns the floceulonodular lobe and the lobulus b the results of Dow are ineluded in order to make the diagram complete.

310 JAN JANSEN AND ALF BBODAL
As fa r as we know a detailed demonstration of this localiza- tion has not been given hitherto, although several authors have pointed out that the different parts of the cerebellar cortex are connected with the par t of the nuclear complex lying immediately beneath the respective lobule. This state- ment obviously holds good in principle.
I t is a matter of considerable interest to see now to what extent our results regarding the cortico-nuclear projection fit iiito the general coiiception of the cerebellum and its connect ions.
I n general it may be said that we are quite well informed concerning the comparative anatomy of the cerebellum, its embryology and its fiber connections. As regards the cerebellar iiuclei, however, our knowledge is still rather incomplete. Nevertheless it is in several instances possible to discover a distinct correlation between the phylogenetic and onto- genetic age of the various parts of the cerebellar cortex and the respective nuclear regions.
Dow ('36, '38) has been able to demonstrate such a eor- relation in regard to the flocculonodular lobe. This most primitive par t of the cerebellum (Herrick, Larsell) both receives direct vestibular fibers (Iiigvar, '18 ; Dov7, '36) and sends i ts efferents to the vestibular nuclei. The lobulus n in addition discharges fibers to the fastigial nucleus (Dow, '36). The fact that the lobulus b (uvula) as well as tlie vermis of the anterior lobe, both being primitive parts of the corpus cerebelli, also project on the vestibular nuclei (mainly the riucleus of Deiters) also is in accordance with the same principle. Besides, these parts also send efferents to the iiucleus fastigii and in greater numbers than to the vestihular nuclei. Turning to the newest part of the vermis, the lobulus G (especially the lobulus e n ) , its efferent connections a re pre- dominantly with the fastigial nucleus. However, some fibers also evidently pass to tlie vestibular nuclei, but these are certainly not so prominent.
Generally, the nucleus dentatus is considered a younger part of the nuclear complex than the nucleus interpositus,

INTRINSIC FIBERS OF THE CEREBELLUM 311
as especially the first undergoes a progressive development in mammals. Embryological investigations (Dowd, '29) corro- borate this view. Apparently, however, this opinion is to a considerable extent based on results from studies of human material, where several facts indicate that the nucleus denta- tus consists of two parts, a ventrolateral (neo-dentatum) and a dorsomedial (palaeo-dentatum). The two parts differ in regard to the cell types found (Gans, '24; Demol6, '27), in regard to the iron reaction (Gans, '24), embryologically (Vogt and Astwazaturow, '12; Bruii, '17; Demol6, '27; et al.) myelogenetically (v. Valkenburg, '12) and under pathological conditions (Rrouwer, '13 ; Brun, '17 ; Koster, '26). Some authors use the name "palaeo-dentatum" synonymously with "nucleus interpositus" in mammals, thus presuming for all mammals the existence of similar relations between the nu- cleus interpositus and the nucleus lateralis, as has been established between the two parts of the dentate in man. Whether this assumption is strictly correct can at present scarcely be settled. Concerning one point, however, viz., the ventrolateral extremity of the dentate, it is obviously wrong as will be shown below. Yet in general the more medial parts of the complex probably are older than the more lateral regions.
Our present knowledge of the efferent connections of the cerebellar nuclei gives no information as to the relative age of the nucleus interpositus and the dentate nucleus, as both these nuclei send their efferent fibers in the brachiurn con- junctivum (Allen, '24). The conflicting statement by Rasmus- sen ('33) is due to his designating part of the nucleus fastigii (in the sense of Brunner, Allen, et al.) as nucleus globosus. Furthermore, fibers probably homologous to the brachium con- junctivum are to be found in Petromyzon (Hausman, '29), likewise such fibers are present in Amphibia (Herrick, '14 ; Larsell, '23, '31, '32 a) , in reptiles (Larsell, '26) and in birds (Schimazono, '12).
With the conception outlined above, viz., that the nucleus dentatus represents a more recent part of the nuclear com-

312 J.4N JANSEN A N D ALF BRODAL
plex than the nucleus interpositus, the cortico-nuclear pro- jection is to a great extent in complete accordance. The lobulus paramedianus, which is considered a relatively old part of the cerebellar hemispheres, showing but small varia- tions, is projected on the intermediate nuclei. The same holds good for the intermediate parts of the anterior lobe (vide infra). The projection area of the lobulus ansiformis, how- ever, is situated farther laterally, in conformity with the fact that this is the part of the hemispheres showing the greatest degree of differentiation and progressive enlarge- ment in the mammalian forms. The projection of the para- flocculus on the extreme ventrolateral part of the dentate riucleus apparently is in contradiction to the conception out- lined above, as the paraflocculus, present also in birds, must be regarded as a rather ancient cerebellar lobule. If accordance between the cerebellar cortex and the cerebellar nuclei can be assumed also on this point, the consequence is that tlie most lateral par t of the dentate nucleus is considerably older than the middle portion of this nucleus. On a closer analysis this contradiction to the generally accepted views seems highly probable. Only if this is the case, can the development of the cerebellar nuclei be said to fit into the general plan of de- velopment as known from other regions of the nervous system, namely that the growth within an “equipotential system” ( Coghill, ’33) takes place through differentiation and pro- liferation from the center and not thi*ougli appositional growth in the periphery. The objection may be raised that investi- gations of the development of the nucleus dentatus in man have failed to demonstrate this. However, this is no wonder, as the paraflocculus in man is only rudimentary and accord- ingly its correlated area in the dentate nucleus supposedly is extremely small. We consider the human paraflocculus as homologous with the paraflocculus in mammals, as most authors hold. Furthermore, i t is worthy of notice, that this conception of the age of the paraflocculus and the correlated lateral par t of the nucleus dentatus is substantiated by the fact that the paraflocculus is projected on the lateral and

INTRINSIC FIBERS OF THE CEREBELLUM 313
caudal part of the principal olive (for details see Brodal, ’40 a).
The projection of the “hemispheral” parts of the anterior lobe must be considered separately. As shown we have been able to demonstrate that in the anterior lobe of the cat the “vermal” region is projected on the fastigial nucleus and the vestibular nuclei, whereas the rest of the anterior lobe sends its efferents to that part of the nuclear complex which by us and other authors is called the nucleus interpositus. It has already been mentioned that in monkeys, however, fibers from the “hemispheral” part of the anterior lobe can also he traced to more lateral parts of the nuclei belonging without doubt to tlie dentate (p. 305), and the assumption was made that in the “hemispheral” parts of the anterior lobe a dis- tinction should be made between an intermediate and a lateral division. Also Clarke and Horsley (’05) find fibers to the dentate nucleus in the monkey, and Saito (’22a, ’22b) de- scribes similar fibers in the rabbit. The apparently conflicting results obtained in cats as compared with monkeys (as well as rabbits) call for a closer analysis.
Granted, in regard to the anterior lobe, a correlation in age between the cerebellar cortex and the corresponding part of the nuclei, the intermediate parts of the anterior lobe must be considered as relatively old divisions. This assumption is in good accordance with the conclusion reached by Larsell (’32b), viz., that the lateral parts of the lobus anterior in mammals are foreshadowed by the pars lateralis in the cere- bellum of reptiles. Furthermore the projection of this part on the dorsal accessory olive (Brodal, ’40 a ) points in the same direction, the dorsal accessory olive being a rather old part of the olivary complex. On the other hand the connections with the dentate nucleus point to the lateral parts of the anterior lobe as being relatively new, a fact in complete ac- cordance with the considerable progressive development of this part of the cerebellum in higher mammals and man. The absence of fibers from the anterior lobe to the dentate nucleus in the cat, however, is surprising. This finding may indicate

314 JAN JANSEN AND ALF BRODAL
that the lateral part of the anterior lobe is wanting in the cat, only the intermediate par t being present. Indeed, this rather astonishing assumption finds strong support in the conditions prevailing within the olivo-cerebellar connections of the anterior lobe. Brodal ( ’40a) has been able to show that whereas the entire ‘ ‘ hemispheral” portion of the anterior lobe in the cat is projected on the medial part of the contra- lateral dorsal accessory olive, this in the rabbit is only true of the “intermediate” part. The lateral part in this animal, on the other hand, is projected on the rostral pole of the principal olive. I n man also the lateral parts of the anterior lobe a re in connection with the rostral part of tbe principal olive (Koster, ’26). Based on these findings, and on an analysis of the morphology of the lateral parts of the anterior lobe in the different species, Brodal ventures the following hypothesis : “Within the lateral parts of the lobus anterior two portions must be differentiated; one older medial portion, better called intermediate, which is projected on the dorsal accessory olive and is present in all mammals, and a newer, lateral portion which is projected on the rostral pole of the principal olive and which is not present in all mammals, but in monkey and man is especially well developed. ”
This hypothesis would be in complete accordance with the fact that the cortico-nuclear fibers from the intermediate portion of the anterior lobe pass to the nucleus interpositus, whereas the fibers from the lateral portion (where such a part is developed) are distribnted to the dentate nucleus. Further, the absence of fibers to the dentate in the cat thus would be accounted for by the “hemispheral” part of the anterior lobe in this animal consisting only of the inter- mediate part.
As a matter of fact i t can be said that the cortico-nuclear localization as determined above fits very well into our knowledge of the cerebellum. It is further of interest that this localizatorp system reveals several features which are in accord with the principles found in the system of the olivo-

INTRINSIC FIBERS O F THE CEREBELLUM 315
cerebellar localization. Fo r details concerning this point tho reader is referred to the paper by Brodal ( ’40 a).
Whether the results obtained in this investigation permit any determination of definite boundaries between the different parts of the nuclear complex is a matter of some importance. Only concerning the most medial part of the complex can a conclusion be drawn: There seems to be no reason for sub- dividing this par t into two nuclei, a nucleus fastigii and a nucleus globosus as some authors do. On the contrary, the cortico-nuclear projection in the cat clearly indicates that the medial mass constitutes an entity, forming the nucleus medialis or fastigii in the sense of Brunner, Allen, e t al., a delimitation also used by us. As appears from the experiments on the rabbit the area of the nucleus interpositus extends somewhat into the nucleus fastigii, as determined on a purely topographical basis. However this deviation from the rela- tions in the cat should not be over emphasized, since the nuclei at this level in the rabbit a re continuous. Concerning the rest of the nuclear complex our investigations give no clue as to the possibility of fixing a n exact limit between the nucleus interpositus and the dentate nucleus. I n fact, the results lend support to the conception that in rabbit and cat, a t any rate, no such boundaries exist. This conception is in accordance both with the findings of other authors in regard to the efferent nuclear connections and our ideas of a funda- mental mutual similarity of the divisions of the “hemispheres ” as compared with the vermis of the corpus cerebelli and the flocculonodular lobe.
We do not intend to consider the question of cerebellar localization in detail. Some points, however, should be empha- sized. The existence of a definite cortico-nuclear localization as determined in this investigation leads one to assume that a similar distinct topographical correlation may exist between the cerebellar nuclei and the parts of the central nervous system to which the efferent fibers of these nuclei a re dis- tributed. This question, as f a r as we know, has up to the present not been investigated and calls for further research.

316 J A N J A N S E N AND ALF BRODAL
We should finally like to call attention to the striking con- formity which exists in the arrangement of the cortico-nuclear and olivo-cerebellar projections, both systems apparently being arranged according to localizatory principles essentially dif- ferent from those prevailing within the spino-cerebellar and vestibulo-cerebellar projections. (MacNalty and Horsley, '09 ; IngT-ar, '18; Beck, '27; Jansea, '31, et al.). Whereas within the former systems the fibers a re so arranged as to favor the conception of a point to point type of localization, MacNaIty and Horsley have demonstrated for the spino-cerebellar system that fibers from different levels of the spinal cord intermingle diffusely within the area of termination in the vermis.
Thus there seem to be two types of anatomical localization within the cerebellum. It is further worthy of notice that the one type, represented by tlie spino-cerebellar and the vestibulo- cerebellar systems (probably also the fibers from the lateral and posterior funicular nuclei belong here) form systems lead- ing afferent impulses from the periphery, whereas the other type, represented by the olivo-cerebellar and the cortico- nuclear (arid probably also the ponto-cerebellar ?) fibers, con- stitute systems forming links in tlie efferent pathways of impulses from the cerebrum via the cerebellum. It must be left to future investigations to decide whether this rather tempting schematization will prove correct.
S T h l X A R Y
After a brief survey of the literature conceriiiiig the cortico- nuclear connections of the cerebellum, the authors report the results of their own investigations on rabbits and cats. Small circumscribed lesions were made in different regions of the cerebellar cortex and the consequent degeneration was followed in Marchi preparations. A number of selected experiments are described and illustrated diagrammatically. The investi- gations have shown that there exists within the cortico-nuclear projection of the cerebellum a definite topographical localiza- tion. This conclusion is chiefly based on the findings made in the experiments on cats, but the resnlts obtained in the more

INTRINSIC FIBERS O F THE CEREBELLUM 31 7
incomplete rabbit material indicate that similar conditions prevail in this animal. The localization in the cat is given diagrammatically in figure 24, where the different lobuli of the cerebellum and the corresponding nuclear regions are marked with identical symbols. For details we refer to the text.
A comparison with the results obtained in this investigation and those of previous authors apparently reveals many con- flicting opinions. However, on a closer analysis, the diver- gencies are seen to be due to a considerable extent to a different nomenclature for the divisions of the nuclear complex, or to the authors’ treatment of entirely different parts of the cerebellum as entities. Discrepancies are also certainly due in part to the interpretation of pseudo-degeneration as real degeneration. When these circumstances are taken into ac- count there is in reality a very good accordance between the results of previous authors and those of the present investi- gation. The importanee of an exact interpretation of the nomenclature used in such studies is emphasized.
The subdivisions used by the present authors is indicated in figures 1 and 2. Attention is called to the fact that any delimitation on a purely morphological basis to a great extent must be arbitrary. In the present study the boundaries be- tween the different components of the nuclear complex have been determined partly on morphological features, partly on the cortico-nuclear relations demonstrated in this investi- gation. An analysis of the localization found in these ex- periments reveals a very good phylogenetic agreement between the different parts of the cerebellar cortex and the corre- sponding part of the nuclear complex.
On the basis of this analysis some conclusions can be drawn. The extreme lateral part of the nucleus lateralis (dentatus) is not the youngest part of this nucleus in mammals as it is in man. The difference is accounted for by the rudimentary development of the paraflocculus in man, the paraflocculus being projected on this part of the nucleus lateralis and being a relatively old part of the cerebellum. I n the hemispheral

318 JAN J A N S E N AND ALF BRODAL
parts of the lobus anterior a distinction should be made be- tween an intermediate zone and a lateral zone, differing with regard to efferent connections as well as in their relation to the inferior olive, as experiments have shown. In the cat the lateral zone of the anterior lobe apparently is not de- veloped. Also in the lobulus ansiformis the efferent projection of the medial and lateral par ts is different, the cortex of the former being projected on the intermediate nucleus whereas the latter projects on the lateral nucleus.
Finally the authors call attention to the similarity of or- ganization within the cortico-nuclear and the olivo-cerebellai. projections, differing in principle from the type of terminal distribution shown by the peripheral afferent connections, the spino- and vestibulo-cerebellar systems. There seem to exist within the cerebellum two different principles of anatomical localization.
LITERATURE CITED
ALLES, WILLIAM F. Distribution of the fibers originating from the different basal cerebellar nuclei. J. Comp. Neur., vol. 36, pp. 399-439.
BECK, G. M. 1927 The cerebellar terminations of the spino-cerebellar fibers of the lower lumbar and sacral segments of the cat. Brain, vol. 50,
Corticofugal and association fibers arising from the cortex of
19%
pp. 60-98. BENDER, L. 1932
BOLK, LOUIS 1906 Das Cerebellum der Saugetiere. Harlem u. Jena. BRADLEY, 0. CHARNOCK 1903
the vermis of the cerebellum. Arch. of Neur., vol. 38, pp. 1-25.
On the development and homology of the mam- malian cerebellar fissures. J. Anat. and Physiol., vol. 37, pp. 112-130 and 221-240.
RRODAL, ALP 1940 a Experiinentelle Untersuchungen iiber die oliro-cerebellare Lok:tlisation. Z . Neur. (In press.)
1940 b The cerebellum of the rabbit. A topographical atlas of the folia as revealed in transverse sections. J. Comp. Neur., t o l . 72, pp. 63-81.
BROUWER, B. 1913 i h e r Hemiatrophia neocerebellaris. -4rch. f . Psychiatr., Bd.
BROUWER, B., AND L. COENEN 1921 Untersuchungen iiber das Kleinhirn. Psych. en New. Blaaden, no. 3 en 4.
BRUN, R. 1917-1918 Zur Kenntiiis der Bildungsfehler des Kleinhirns. Schweiz. ,4rch. f. Neur. u. Psychiatr., Bd. 1, S. 61-123, Bd. 2, S. 48-105, Bd. 3, S. 13-88.
1919 Die zentralen Kleinhirnkerne bei den Saugeticrcn. Arb. Neur. Inst. Wien., Bd. 22, S. 200-277.
51, 8. 539-577.
RRDNNER, H.

INTRINSIC FIBERS O F T H E CEREBELLUM 319
CLARKE, R. H., AND V. HORSLEY On the intrinsic fibers of the cerebellum, its nuclei and its efferent tracts. Brain, vol. 28, pp. 13-29.
COGHILL, G. E. 1933 Growth of a localized functional center in a relatively equipotential nervous organ. Arch. Neur. and Psychiat., vol. 30,
DENOI.~, V. Structure et connexion des noyaux dentCl6s du cervelet. Schweiz. Arch. f . Neur. u. Psychiatr., Bd. 20, 8. 271-294, Bd. 21, S. 73-110.
Dow, ROBERT S. 1936 The fiber connectioiis of the posterior parts of the cerebellum in the cat and rat. J. Comp. Neur., vol. 63, pp. 527-548.
1938 Efferent connections of the flocculonodular lobe in Macaca mulatta. J. Comp. Neur., vol. 68, pp. 297-306.
DOWD, LESLIE W. 1929 The development of the dentate nucleus i n the pig. J. Comp. Neur., vol. 48, pp. 471-499.
GANS, A. 1924 Beitrag zur Kenntnis des Aufbaues des Nucleus dentatus aus zwei Teilen, nameiitlich auf Grund von Untersuchungen mit der Eisen- reaktion. 2. Neur. u. Psych., Bd. 93, S. 750-755.
HAUSNAN, LOUIS 1929 The comparative niorphology of the cerebellar vermis, the cerebellar nuclei and the vestibular mass. Research publications of Association for Research in Nervous and Mental Disease, vol. VI , The Cerebellum, pp. 193-237. Baltimore.
H A Y ~ ~ H I , M. 1924 Einige wiehtige Tatsaehen aus der ontogenetischen Ent - wicklung des inenschlichen Kleinhirns. Deutsche Z. f . Nervenheilk.,
HERRICK, C. JUDSON 1914 The cerebellum of Necturus and other urodele Amphibia. J. Comp. Neur., vol. 24, pp. 1-29.
1924 Origin and evolution of the cerebellum. Arch. Neur. and Psychiat., vol. 11, pp. 621-652.
HOHUAN, LESLIE B. 1929 The efferent connections of the cerebellar cortex; investigations based upon experimental extirpations in the cat. Re- search publications of Association for Research in Nervous and Mental Disease, vol. VI. The Cerebellum, pp. 445-460. Baltimore.
INGVAR, SVEN 1918 Zur Phylo- und Ontogenese des Kleinhirns nebst einem Versuche zu einheitlicher Erklarung der zerebellaren Funktion und Lokalisation. Folia Neurobiol., Bd. 11, S. 205-495.
JANSEN, JAN 1931 Om cerebellum og cerebellar lokalisation. Norsk. Mag. f. Laegevidensk., S. 789-805.
Experimental studies on the intrinsic fibers of the cerebellum. I. The arcuate fibers. J. Comp. Neur., vol. 57, pp. 369-400.
KAPPERS, C. U. ARIENS, G. CARL HUBER AND ELIZABETH C. CROSBY The Comparative Anatomy of the Nervous System of Vertebrates, including Man. New York.
KOSTER, S. 1926 Two cases of hypoplasia ponto-neocerebellaris. Acta Psychia- trica et Neurologica, vol. 1, pp. 47-83.
KUZUNE, G. 1926 Experimentell-anatoniische Untersuchungen iiber die inneren und ausseren Verbindungen des Flocculus und der Kleinhirnkerne (hauptsachlirh des Dachkerns). Fol. . b a t . Jap., vol. 4, pp. 75-110.
1905
pp. 1086-1091. 1927
Bd. 81, S. 74-82.
1933
1936

320 J A N J A N S E N A N D A L F BRODAL
LARSELL, 0. 1920 The cerebellum of Amblystonia. J. Comp. Neur., rol. 31,
The cerebellum of the frog. J. Comp. Neur., vol. 36, pp. 89-112. The development of the cerebellum in the frog (Hyla regilla)
in relation to the vestibular and lateral-line systems. J. Comp. Neur., vol. 39, pp. 249-289.
1926 The cerebellum of reptiles: lizards and snake. J. Colnp.
1931 The cerebellum of Triturus torosus. J. Comp. Neur., 801. 53,
1932 a The development of the cerebellum in Amblystoma. J.
1932 b The cerebellum of reptiles: chelonians and alligator. J.
1934 Morphogenesis and evolution of the cerebellum. Arch. New.
1935 The development and morphology of the cerebellum in the opossum. 11. Later development and adult. J. Comp. Neur., rol. 63.
____ 1936 Cerebellum and vorpus pontobulbare of the bat (Myotis). J. Comp. Neur., vol. 64, pp. 275-302.
1937 The cerebellum. A review and interpretation. Arch. Neur. and Psychiat., vol. 38, pp. 580-607.
LARSELL, OMF, AND ROBERT STONE Dow 1935 The development of the cere- bellum in the bat (Corynorhinus sp.) and certain other mammals. J. Comp. Neur., vol. 62, pp. 443-468.
Lowr, ROBERT 1916 ober die Faseranatomie und Physiologie drr Forniatio vermicularis cerebelli. Arb. Neur. Inst. Wien., Bd. 21, S. 359-382.
MACNALTY, S., AND V. HORSLEY On the cervical spinobulbar and spino- cerebellar tracts and on the question of topographical representation in the cerebellum. Brain, vol. 32, pp. 237-255.
MUSSEN, AUBREY T. 1927 Experimental investigations on the cerebellum. Brain,
1929 Experimental investigations of the cerebellum. Research publications of Association for Research in Nervous and Mental Disease, vol. 6, pp. 381-423, Williams and Wilkins, Baltimore.
RASMUSSEN, A. T. 1932 Secondary vestibular tracts in the cat. J. Comp. Neur., vol. 54, pp. 143-171.
1933 Origin and course of the fasciculus nncinatus (Russell) in the cat with observations on other fiber tracts arising from the cerebellar nuclei. J. Comp. Neur., vol. 57, pp. 165-197.
RILEY, HENRY ASLOP 1929 The mammalian cerebellum. A comparative study of the arbor vitae and folial pattern. Research publications of Association for Research in Nervous and Mental Disease, vol. 6, The Cerebellum, pp. 37-192. Baltimore.
pp. 259-282. 1923 1925
Neur., vol. 41, pp. 59-94.
pp. 1-54.
a m p . Neur., vol. 54, pp. 357-435.
Comp. Neur., vol. 56, pp. 299-345.
and Psychiat., vol. 31, pp. 373-395.
pp. 251-292.
1909
v01. 50, pp. 313-349.

INTRINSIC FIBERS OF T H E CEREBELLUM 321
SAITO, MAKOTO 1922 a Experimentelle Untersuchungen iiber die inneren Ver- bindungen der Kleinhirnrinde nnd deren Beziehungen zu Pons und Medulla oblongata. Arb. Neur. Inst. Wirn., Bd. 23, S. 74-106.
1922 b Weitere Untersuehungen iiber die inneren Verbindungen der Kleinhirnrinde. Der Lobus anterior. Arb. Neur. Inst. Wien., Bd. 24, S. 77-84.
SCHINAZONA, J. 1912 Das Kleinhirn der Vogel. Arch. f . mikrosk. Anat., Bd. 80, 5. 397-449.
SNITH, G. ELLIOT 1902 The primary subdivision of the mammalian cerebellum. J. Anat. and Physiol., vol. 36, pp. 381-385.
1903 Further observations on the natural mode of subdivision of the mammalian cerebellum. Anat. Anz., Bd. 23, S. 368-384.
Bijdrage tot de Kennis eener Localisatie in de inenschelijke klcine Hersenen. Nedeil. Tijdschrift voor Genceskunde,
VOGT, H., AND M. ASTWAZATUROW 1912 Ueber angeborene Kleinhirnerkrank- ungen mit Beitrage zur Entwicklnngsgeschichte des Kleinhirns. Arch. f . Psychiatr., Bd. 49, S. 75-203.
VOGT, 0. 1905 Die myclogenetische Gliederung des Cortex Cercbelli. J. Psychol. und Neur., Bd. 5, S. 235-250.
WEIDENREICH, F. 1899 Zur Anatomie der zcntralen Kleinhirnkerne der Sauger 2. Morph. und Anthrop., Bd. 1, S. 259-312. (Cited after Brunner.)
VAN VALKENBURG, C. T. l O l Z
TO^. 1, pp. 6-24.