feeding kinematics and nectar intake of the honey bee tongue
TRANSCRIPT

Feeding Kinematics and Nectar Intake of the HoneyBee Tongue
Rengao Zhu1,2& Hesong Lv2 & Tianye Liu2
&
Yunqiang Yang1 & Jianing Wu2& Shaoze Yan2
Abstract Most flower-visiting insects employ highly-evolved organs to feed them-selves rapidly and efficiently on the floral nectar. A honey bee drives its segmentedtongue (glossa) covered by dense hairs reciprocatingly to load nectar. A high-speedcamera system ameliorated by a microscope revealed morphological changes in glossalsurfaces during live honey bees’ nectar dipping and surface configurations through thestretching of postmortem honey bees’ glossae. Both the in vivo and postmortemobservations reveal that shortening and lengthening of the glossal segments performhigh concordance with the erection of glossal hairs, which aids in developing deform-able gaps between rows of glossal hairs during nectar trapping. A model was proposedto evaluate the nectar-intake volume considering the experimentally-measured averageerection angle and tongue elongation length during nectar feeding. The theoreticalresults fit the experimental data well and disclose that these two factors contribute to anaugmentation of nectar-intake observably. We also theoretically present that theextendible and deformable glossae have advantages for the polylectic feeding behavior.
Keywords Honey bee . nectar feeding . segmented structure . nectar-intake volume
DOI 10.1007/s10905-016-9561-5
Rengao Zhu and Hesong Lv contributed equally to this study.
Electronic supplementary material The online version of this article (doi:10.1007/s10905-016-9561-5)contains supplementary material, which is available to authorized users.
* Yunqiang [email protected]
* Shaoze [email protected]
1 School of Engineering and Technology, China University of Geosciences (Beijing),Beijing 100083, People’s Republic of China
2 Division of Intelligent and Biomechanical Systems, State Key Laboratory of Tribology,Department of Mechanical Engineering, Tsinghua University, Beijing 100084, People’s Republicof China
Revised: 6 May 2016 /Accepted: 24 May 2016 /Published online: 9 June 2016# Springer Science+Business Media New York 2016
J Insect Behav (2016) 29:325–339

Introduction
Liquid-feeding behavior plays an important role in bringing organisms water, sugar,amino acids, organic acids, proteins, fats, vitamins, minerals and other minor compo-nents (Corbet 2000; Bertazzini et al. 2010). There has an emphasis in investigating thedrinking patterns of animals that use tongues or tongue-like devices to load fluid(Karolyi et al. 2014; Krenn 2010; Kingsolver and Daniel 1983; Winter and vonHelversen 2003). Most tongues and tongue-like devices are considered just as rigidbodies with permanent surfaces during drinking process, such as the tongues ofbutterflies (Lee et al. 2014), shorebirds (Prakash et al. 2008), cats (Reis et al. 2010)and dogs (Crompton and Musinsky 2011). The butterfly proboscis is a complexhydrophobic–hydrophilic dichotomous and hierarchical system having nanopores toprovide strong capillarity and micropores to facilitate fluid transport, which can formliquid bridges even when the sucking pump is not activated (Monaenkova et al. 2012;Lehnert et al. 2013). In contrast to the fluid-uptake mechanics by butterflies, nectar-feeding bats employ a typical lingual dynamic surface during nectar feeding to improvefeeding efficiency (Harper et al. 2013). Blood vessels within the tongue tip ofG. soricina can engorge with blood and then the papillae on the tongue surface becomeerect while dipping nectar. With tumescence and papilla erection persisting throughouttongue retraction, the nectar trapped between rows of erected papillae is carried into themouth. Such dynamic surfaces equipped with erectable protrusions help increase nectarintake volume by increasing the surface area (Harper et al. 2013).
Nectar, an aqueous solution of sugar and some chemical ingredients, providesimportant nutrient sources for honey bees (Abou-Shaara 2014; Jensen et al. 2013;Martínez del Rio et al. 2001). The nectar drinking strategy has been studied extensivelyand the erection of glossal hairs has been revealed in several researches (Briant 1884;Cook 1880; Dade 2009; Snodgrass 1956; Snodgrass 1984). Especially in our previousstudy, the rhythmical erection pattern of honey bees’ glossal hairs in nectar feeding hasbeen reported, but the role of the deformable tongue structure in nectar feeding has notbeen investigated yet (Wu et al. 2015). In this work, we focused on the deformation ofthe tongue (glossa) surface of honey bees, especially the morphing of the segmentedstructures, and its influence on the nectar-intake efficiency.
Feeding in honey bees involves the process of nectar coating and nectar uploading(Kim et al. 2011). When the glossa inserts into nectar, the glossal surface of a honey beeis coated by nectar. Analogously to the model established for uptake of liquid from wetsurfaces by a brush-tipped proboscis, the nectar coating of honey bee’s glossal surfacein viscous fluid may be mainly caused by wetting and capillary (Lee and Lee 2014;Chen et al. 2015). For the nectar uploading process, whether all of the nectar betweenthe glossal hairs can be impelled in the viscous nectar is really important. The fluid flowaround and through rows of cylindrical hairs depends on the size, spacing, and speed ofthe hairs that is relative to the fluid (Koehl 2001; Loudon et al. 1994; Cheer and Koehl1987). The ‘leakiness’ of a gap between neighbouring hairs in an array is given byκ=U/Ux, where U is the volume of the fluid that flows through the gap in a unit of timeandUx is the volume of fluid that would flow at a free stream velocity through the spaceif there were no hairs (Koehl 2001). The appendage bearing arrays of hair-likestructures functions like a sieve or a paddle judging by the leakiness of a row of hairson a structure(Koehl 2001). The leakiness of pairs of hairs is related to the Reynolds
326 J Insect Behav (2016) 29:325–339

number Re=ρva/μ (the ratio of inertial to viscous forces) and hair spacing G/2a (G isdefined as the width of the gap). The ‘leakiness’ of hair gaps can be used to evaluate theuploaded volume in a nectar feeding cycle. As a preliminary, the glossa was modeled asa simple slender cylinder and the fluid was entrained by the outer surface of the glossa(Kim et al. 2011). Another model was presented in which the glossa was considered tobe a hairy cylinder under the condition of viscous dipping (Yang et al. 2014).The model developed by Yang et al. suggests the hairs contributes to the nectaruploading but the erection angles in their model remains 90 deg. In this study,we presented a model, in which the erection angle of their model was amendedand the influence of the elongation of the segments was added in considering theexperimental results.
Materials and Methods
Preparation of Honey Bee Specimens
The honey bees (Apis mellifera ligustica) were captured in Beijing, China(40.00°N, 116.33°E) and fed in the laboratory in a beehive, where the temper-ature and humidity were maintained at 25°C and 50 %, respectively. To mimicthe natural environment, the beehive was illuminated with a light-darknesscycle, following a cycle of 10-h day time and 14-h night time. The honeybees inside were fed with 35 % sucrose solution. We confirmed that no specificpermissions were required for these locations/activities, and the field studies didnot involve endangered or protected species.
Proboscis Morphology
To examine the external morphology of the worker honey bee’s mouthpart, ten glossaspecimens of the honey bees were observed under a microscope (Axiostar Plus, Zeiss,Germany). Another ten glossa specimens were made into 500 μm thin slices andimaged prior to toluidine blue staining. To obtain the microstructure of the surface,ten samples were soaked in 2.5 % glutaraldehyde solution for one night and cleaned by0.1 mol/L phosphate buffer (PH = 7) three times (10 min each), dehydrated by anethanol series (75, 80, 85, 90, 95, 100 %, 30 min each) prior to air drying. A graphiteadhesive tape was used to mount them on an SEM viewing stage. The samples werecoated with gold (~10 nm) and examined with a scanning electron microscope (SEM,FEI Quanta 200, Czech Republic).
Nectar Feeding Experiments
To obtain the experimental data referring to the volumetric intake rate of ahoney bee, two or three worker honey bees were captured from beehive andput into a glass bottle, prior to which a plastic dish was placed at the bottom ofthe bottle (Fig. 1b). Then the 20 μL droplets of 35 % (wt. /wt.) sucrosesolution were placed at the flat dish by a micropipette (Top Pipette20 ~ 200 μL, DragonLab, China ). We recorded the time from the beginning
J Insect Behav (2016) 29:325–339 327

of sucrose imbibing until a bee had completely consumed a droplet. In orderto observe the micrometer-length tongue covered by bushy hairs and its swiftmovements, we applied a high-speed camera (Olympus, iSpeed TR, Japan)equipped with a microscope (Axiostar Plus, Zeiss, Germany) to film thedrinking process (Fig. 1c,d). In addition to the high-speed camera system, wefabricated a special fixture for fixing honey bees, a glass nectar feeder and a3D-motion precision platform. The fixture clamping a honey bee in the thoraxwas controlled by the motion platform to adjust a honey bee to an appropriateposition where it could drink the artificial nectar. The artificial nectar wasprepared by the 35 % (wt./wt.) sucrose solution. The honey bee specimenswere fed in another isolated beehive with temperature of 25 °C and humidity of50 % and starved for 12 h before we did the nectar feeding experiment (Wuet al. 2015). Feeding process was recorded at 500 fps with the enlargementratio of 5X in vertical view.
Glossa Stretching Experiments
These experiments were conducted to demonstrate the relationship between thesegmental elongation length and the hair erection angle. The experiment setupwas comprised of a high-speed camera (Phantom M110, USA), a microscopetube (Keyence, VH-Z50L, Japan, up to the magnification of 50X), a specialfixture and a 3D motion platform (Fig. 1e, f). But the fixture and the platformwere different from the ones used in the nectar feeding experiments to clampthe dead honey bee and needed to withstand a certain stretching force invertical direction.
Prior to the experiment, a honey bee was immersed into the ethyl alcoholand the heads of honey bee samples with their glossae extended were used forthe postmortem tests immediately. The distal portion of the glossa was stuck toa fixed capillary tube (1 mm diameter) with cyanoacrylate glue while theproximal portion with the body clamped by the fixture was controlled by themotion platform. During the experiment, the glossa was stretched with a step-by-step motion accuracy of 200 μm. The process was recorded at 500 fps witha magnification ratio of 5X.
Data Processing and Statistical Analyses
The high speed images of the nectar feeding and the glossa stretching exper-iment were analyzed by the software of Phantom Camera Control (PhantomM110, USA). When measuring the segmented structure length and the erectionangles of glossal hairs, we captured and selected 9 ~ 10 images in a completeerection process sequentially. We measured the length from the 10th to the 19thsegments (the first segment is initialized from the distal apex) and calculatedthe average length L. Moreover, in order to measure the glossa hairs’ erectionangles accurately, a vector method was introduced. We chose two hair strandsadhered to the same membrane but on separate sides. Then we named fourendpoints as: A1, A2, B1 and B2. In a coordinate system, the four endpoints hadcoordinates A1(x1,y1), A2(x2,y2), B1(x3,y3), and B2(x4,y4). Therefore these two hairs
328 J Insect Behav (2016) 29:325–339

could be illustrated as two vectors a= (x2− x1, y2− y1), b= (x4− x3, y4− y3). The angle θ
between two hairs was defined as θ ¼ arccosa:baj j: bj j (Wu et al. 2015). We used Origin
8.0 (OriginLab) to calculate the Pearson correlation coefficient between data sets of hairerection and segment elongation:
Correl L; θð Þ ¼X
L− L̅� �
θ− θ̅� �
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiXL− L̅
� �2Xθ− θ̅
� �2r ð1Þ
Results
Proboscis Morphology
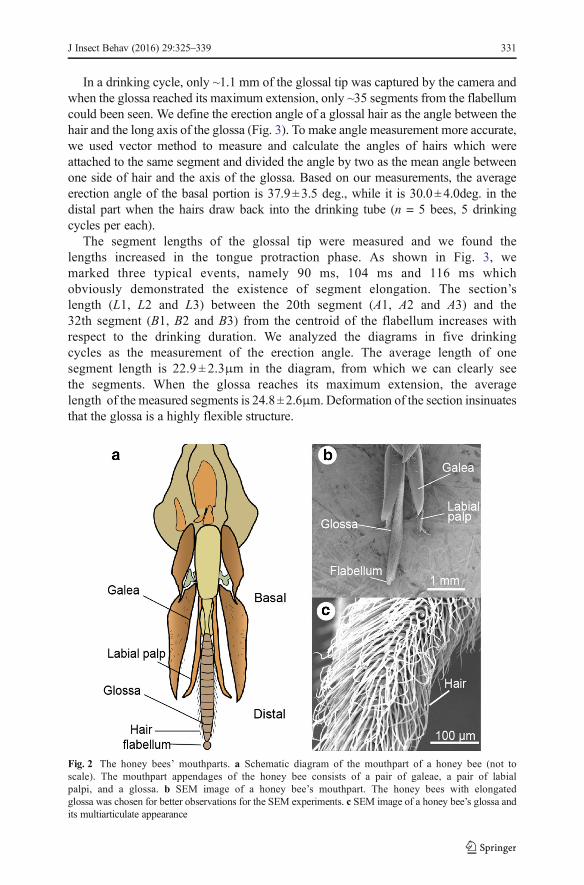
The honey bee mouthpart subassemblies for nectar feeding are composed of apair of galeae, a pair of labial palpi and a glossa with a flabellum in its distalapex (Fig. 2). The surface of a glossa is segmented and we regard one segmentas the portion from one hair base to the next one. The ring-like intersegmentalmembranes are the boundaries of each segment and the sites of insertion for thedense glossal hairs. Then the general appearance of a glossa is a slightlytapering brush-like organ densely covered with long hairs bearing on thesegmental membranes.
Nectar-Feeding Behavior
We manufactured a glass feeder filled with artificial sucrose solution (35 %, wt./wt.)for honey bees to dip (see Supplementary Material movie S1) and analyzed thezoomed-in high-speed images to investigate the nectar-loading behavior of a honeybee’s glossa during feeding process. Before drinking cycles, the mandibles of ahoney bee open and the previously contracted proboscis unfold to the feedingposition and the galeae and labial palpi align together to form a temporary tubewhich holds the glossa inside. Then the tube remains relatively motionless while thehoney bee trap the nectar by extending and retracting their glossa periodically (Wuet al. 2015). A feeding cycle is categorized into the phases of tongue protraction andtongue retraction based on the motion properties of the tongue body. In the tongueprotraction phase, the glossa extends out of the tube with its bushy hairs attached tothe glossal rod. When the glossa nearly reaches its maximum displacement, theglossal hairs begin to swiftly deploy and the nectar fills the gaps structured by rowsof glossal hairs, then the glossa extends to the limit instantly. In the retraction phase,the glossa retreats from the liquid, and the deployable structures form a cone-like shapewhile the glossal hairs continue to deploy until they reached the maximum erection andkeep stationary to load nectar into the proboscis (Wu et al. 2015). The glossal hairsflatten instantaneously to the same configuration as that of the initialization when thewhole glossa retracts into the tube. The mean duration of a single dipping cycle, from
J Insect Behav (2016) 29:325–339 329

the start of glossa protrusion to the beginning of next protrusion, is 0.20±0.03 s(mean ± S.D., n = 5 bees, 5 drinking cycles per each). After drinking, the mouthpartsubassemblies retract back to the natural rest position. Figure 3 depicts 12 typical high-speed images of the temporal variation in erection angle of the glossal hairs in a liquid-feeding cycle.
Fig. 1 Honey bee feeding nectar and the experimental setups. a In-feeding honey bees. b Image of two honeybees feeding nectar. c The setup of nectar feeding experiments. In the upper right-hand corner, the enlargefigure shows that the honey bee is right in front of the glass feeder with its proboscis stretching into the sucrosesolution. d Nectar feeding system. The camera was connected with a computer by a 1000 MEthernet cable (the yellow cable in this subfigure). e The setup of glossa stretching experiments.The glossa was stretched with one side fixed to a platform and the other side moving toward the oppositedirection. f Glossa stretching system
330 J Insect Behav (2016) 29:325–339
In a drinking cycle, only ~1.1 mm of the glossal tip was captured by the camera andwhen the glossa reached its maximum extension, only ~35 segments from the flabellumcould been seen. We define the erection angle of a glossal hair as the angle between thehair and the long axis of the glossa (Fig. 3). To make angle measurement more accurate,we used vector method to measure and calculate the angles of hairs which wereattached to the same segment and divided the angle by two as the mean angle betweenone side of hair and the axis of the glossa. Based on our measurements, the averageerection angle of the basal portion is 37.9±3.5 deg., while it is 30.0±4.0deg. in thedistal part when the hairs draw back into the drinking tube (n = 5 bees, 5 drinkingcycles per each).
The segment lengths of the glossal tip were measured and we found thelengths increased in the tongue protraction phase. As shown in Fig. 3, wemarked three typical events, namely 90 ms, 104 ms and 116 ms whichobviously demonstrated the existence of segment elongation. The section’slength (L1, L2 and L3) between the 20th segment (A1, A2 and A3) and the32th segment (B1, B2 and B3) from the centroid of the flabellum increases withrespect to the drinking duration. We analyzed the diagrams in five drinkingcycles as the measurement of the erection angle. The average length of onesegment length is 22.9 ± 2.3μm in the diagram, from which we can clearly seethe segments. When the glossa reaches its maximum extension, the averagelength of the measured segments is 24.8±2.6μm. Deformation of the section insinuatesthat the glossa is a highly flexible structure.
Fig. 2 The honey bees’ mouthparts. a Schematic diagram of the mouthpart of a honey bee (not toscale). The mouthpart appendages of the honey bee consists of a pair of galeae, a pair of labialpalpi, and a glossa. b SEM image of a honey bee’s mouthpart. The honey bees with elongatedglossa was chosen for better observations for the SEM experiments. c SEM image of a honey bee’s glossa andits multiarticulate appearance
J Insect Behav (2016) 29:325–339 331

Coordination of Hair Erection and Segment Elongation
Comparing hair erection and segment elongation, we found a high consistencyof their kinematics during the drinking process (Fig. 4). In a dipping cycle,when the average erection angle of whole hairs increases from 20 deg. to38 deg., the average length of one glossal segment increases from 22.9 ± 1.6μm to 24.7 ± 2.2 μm. The Correl function can verify potential correlationbetween these variables. The absolute valueof the Correl function is closer to1, the two variables are strongly correlated. The concordance measure is equalto 0.99 in our in vivo observation experiments, which shows that the averageelongation length of a glossal segment is closely correlated to the averageerection angle of hairs. To further demonstrate the relationship between them,we conducted supplementary experiments in which we stretched the glossa of10 postmortem honey bees. In these experiments, the hairs of a dead honeybee’s glossa erect only when the glossa is stretched by an external force, whilethey could not erect by inserting the glossa into the fluid without intervention,thus it is the stretching force that causes the hair erection and segmentelongation. The average erection angle of hairs increases from 23 deg. to57 deg., and the average length of one glossal segment increases from34.7 ± 2.8 μm to 37.7 ± 3.1 μm accordingly. The concordance value is 0.96,which is close to the nectar feeding experiment (0.99). Though the maximum erectionangle of glossal hairs and final elongation length of one segment in the postmortem testsis bigger than in vivo observations (one-tailed paired t test: hair erection angle t24 = 18.3,p < 0.001; segment elongation length t24 = 1.93, p = 0.032), the hair erection andsegment elongation occurs in synchrony in both experiments. This high coordinatedmotion indicates that the glossal hairs are hinged in the intersegmental membranes,which could deploy and fold synchronously.
Fig. 3 Glossa movements in a feeding process captured from a high-speed video. The glossal hairs stay flatagainst the glossal surface at the beginning of a drinking cycle. Just before the glossa approaches the maximumextension, the glossal hairs erect and the distal segments elongate. Then the configuration of glossal surfaceholds throughout the glossa retraction stage and the solution is trapped between the rows of hairs until it isingested by the cibarial pump. The parameter α is defined as the erection angle of glossal hairs
332 J Insect Behav (2016) 29:325–339

Liquid-Feeding Model of Honey Bees
According to our observations, in the whole process of a drinking cycle, the erectionand flattening movements of glossal hairs occur during the forward and backwardmovements of the glossa without bending. Thus the glossal hairs can be regarded asrigid structures in the dipping process. For a honey bee drinking a 35 % (w./w.) sucrosesolution in our lab environment, the Reynolds number Re<10−3 and hair spacing G/2a=7±2. The model of Cheer and Koehl suggests that the leakiness of a honey bee’sglossa is less than 0.05 (Loudon et al. 1994). So a honey bee can employ its thick hairyglossa propelling the fluid between hairs toward the drinking tube even though it is fullof gaps between neighbouring hairs (Fig. 5). There are no air bubbles formed on glossalsurface during feeding, thus the space between hairs are filled with nectar when theglossa retracts.
Based on the above results, we present a drinking model of honey bee’sfeeding behavior in which the glossa is considered as a flexible multisegmental
Fig. 4 Hair erection and segment elongation of the glossa. Nine videos, each of which eight frames before themoment that glossa reaches its maximum extension were used for analysis of the hair erection and segmentelongation. a The result of nectar drinking experiment. b The result of glossa stretching experiment. From bothof the experiments, we found the high coordination between the hair erection and segment elongation
J Insect Behav (2016) 29:325–339 333

structure with thick hairs. As shown in Fig. 6, ri and β are radius and taperangle of the glossa in nectar dipping, respectively. hi represents the length ofthe erected hairs and xi is the axial length of one segment. The angle betweenthe two opposite hairs are defined as θi= 2αi, in which αi is the erection angleof glossal hairs. The thickness of the adhered nectar layer is ei= hi sin (αi+β)/cosβ. Then we can present the nectar-intake volume V of a honey bee in thefollowing form:
V ¼ πXn
i¼1
2eiri þ ei2−2sa2hi=3
� �xi ð2Þ
where s is the density of hair distribution.According to our experimental results and the theoretical model, we specu-
late that the erection pattern of hairs and the increase of the segmental lengthare effective for a honey bee to get better nectar rewards. We calculated thenectar intake volume which relied on viscous entrainment by the permanentsurface of the tongue. When the glossa inserts into the nectar, the surface iscoated with fluid. The intake volume could be written in the following waybased on Kim’s model (Kim et al. 2011):
V ¼ π 2e0iri þ e
0i
2� �
L ð3Þ
in which L is the glossal length. The thickness of the nectar layer coated on thepermanent glossal surface in the fluid of viscosity μ and surface tension σ ise0i~Ca
2/3ri, where Ca=μv/σ is the Capillary number (Quéré 1999). To calculatethe intake volume of a drinking cycle, proximate equations are given based onequations (2) and (3):
Fig. 5 Conceptual hypothesis for glossa movements during the drinking process. The hairs erect coincidentlywith the elongation of segments, and when the erection angles come to its maximum, the glossa retracts withthe space between hairs filled with nectar
334 J Insect Behav (2016) 29:325–339

Qd ¼ π 2e r0−X e
2tanβ
� �þ e2−2sa2h=3
Le f
Qp ¼ π 2e0r0−
X e
2tanβ
� �þ e
02
Lf
8>><>>:
ð4Þ
The volumetric intake rateQp was ~0.016 μL/s with a permanent glossal surface andthe volumetric intake rate Qd was ~0.965 μL/s with dynamic surface that the glossalhairs erected and glossal segments elongated in nectar-dipping cycles, which indicatedthat the dynamic deformation could increase the dipping volume by ~60 times. Valuesused to calculate the volumetric intake rate of a honey bee are listed in Table 1. Wefilmed the imbibing process of 20 μL droplets by honey bees (see SupplementaryMaterial movie S2) and found that it took a honey bee 22.0±3.3 s to ingest it. Thus theaverage volumetric consuming rateQe of a honey bee is 0.91 μL/s. However, the honeybees cannot immerse their whole glossa into the nectar as the nectar droplets arecontinuously consumed leading to the reduction of employed glossa length, so theexperimentally-average intake rate is relatively lower than the theoretical predictions.
Discussion
In this study, the morphology of the honey bees’ mouthparts and their liquid-feedingprocesses were investigated experimentally. We observed the glossal surface morphol-ogy and analyzed the dynamics of glossal hairs, together with the multi-segmented
Fig. 6 Schematic diagram of the physical model. The glossa is simplified as a flexible taper which bearsarrays of hairs functioning like a paddle propelling the fluid between hairs toward the drinking tube. Eachsegment has a length xi while the hairs in it have an average length hi and an average erection angle αi
J Insect Behav (2016) 29:325–339 335

structures. We found high degree of rhythmicity in the movement of glossal surface andstrong synchronicity between the motion of glossal hairs and segments during thenectar-dipping process. Furthermore, we demonstrated the high accordance betweenthe hair erection and segment elongation by an additional experiment. Finally,we established a nectar drinking model of the honey bee and compared theintake volume to the previously-proposed models, confirming that the hair erection andsegment elongation can remarkably contribute to augmenting the nectar-intake volumeof honey bees.
Maximizing net meal energy is a general demand of many animals, but how it isachieved may vary depending on the usage of different foraging techniques(Hainsworth et al. 1991; Paul and Roces 2003). Tongues of a variety of animals, whichhave dynamic surfaces, could reconfigure their surface profiles to improve the nectar-uptake volume (Harper et al. 2013; Howell and Hodgkin 1976; Kim et al. 2012; Rico-Guevara and Rubega 2010). Erection of hair-like papillae in bats driven by blood flowproduces rapid changes in tongue surface area during nectar lapping (Harperet al. 2013). Hummingbird tongues driven by release of surface tension whenthe tongue enters the nectar and elastic recoil of the keratinous lamellae alsoundergo rapid changes in tongue surface area during lapping (Rico-Guevara andRubega 2010). In this paper, the glossal surface forms a natural volume-variablecontainer to load nectar, which is structured by the deformable hair-equippedsegments. The dynamic surface not only changes its surface configuration byunfolding the glossal hairs, but also elongates the segmented structures alongthe long axis of the tongue, which can further enhance the volume for nectarfeeding. In this way, the tongue of a honey bee could derive the maximumamount of nutrients while being withdrawn, like a sponge that is alternately allowed toexpand to take up water and squeezed to expel it (Simpson and Riedel 1964). Accordingto the theoretical analysis, deformation of glossal surface can increase the thickness ofadhered nectar through the combined action of viscosity and capillarity, which willbenefit the honey bees’ feeding efficiency.
To achieve efficient energy rewards, elongated mouthpart appendages evolvedconvergently on numerous occasions in flower-visiting insects as nectarivorous animalsneed to protrude their mouthparts into the base of corolla tube when dipping nectar in anatural environment (Harder 1986; Nilsson 1988; Krenn et al. 2005; Wilhelmi andKrenn 2012). When honey bees forage on the flowers, they need to immerse theirglossa as deep as possible to trap more nectar as the nectaries are located at the bottom
Table 1 Parameters forcalculating the volumetric intakerate of a honey bee
Parameter Description Value
α Glossa erection angle 40 deg.
β Glossa tilt angle 3 deg.
r0 Glossa radius 100 μm
a Hair radius 3 μm
s Hair distribution density 0.8 /μm
Xe Glossa length 2500 μm
f Dipping frequency 5 /s
h Hair length 130 μm
336 J Insect Behav (2016) 29:325–339

of the corolla tube, while suctorial foragers just need to immerse their proboscis tipsinto the nectar (Krenn et al. 2005; Wilhelmi and Krenn 2012). Thus the length of honeybees’ glossae is crucial for their foraging efficiency. But the movements in insects withextremely long proboscis during feeding are slower than the movements in insects withaverage-sized proboscis, which will produce exceptionally augmentation of functionalcosts and reduction of foraging efficiency (Bauder et al. 2011). A relatively longproboscis insect also need more space for unfolding of the proboscis and thus someof the extremely long-tongued animals still hover while they are foraging, so extendiblemouthparts have advantages for the feeding performance of insects (Krenn et al. 2002).In most pollination systems, a given pollinator often visits a wide variety of flowerspecies and Apis mellifera is one of the polylectic bee species (Aronne et al. 2012;Shimizu et al. 2014; Hilgert-Moreira et al. 2014). The honey bees’ lissome segmentedglossal structures can increase the flexibility of the glossa, as the movements ofmulti-segmented glossa exhibit a higher degree of freedom and allow them formore adequate responses to flower corollas of varying width and depth. Inaddition, a flexible tongue makes it easier to sweep the corolla tube in whichsmall amount of nectar are available in the target flowers. Thus the extensible mouthpartcan reduce the effect of different drinking conditions on flower handling efficiency(Harder 1986).
Precise coordination of distributed parts in organs aids animals accomplishingcomplex functions during physiological activities. The close consistency between theerection of glossal hairs and elongation of glossal segments indicates that the glossa ofa honey bee is a highly-evolved and subtle mechanism. Driven by the muscles and anelastic glossal rod, the glossa can be elongated and compacted, with the glossalhairs erected and flattened synchronously, and the tiny membrane-structuredglossal segments create possibilities for linking these two types of motionstogether (Simpson and Riedel 1964). The muscles slowly load the spring-likeintersegmental membranes producing the erection of glossal hairs and elonga-tion of segments and later the release of potential energy causes the flatteningof hairs immediately. The active movements of glossal muscles can preciselycontrol the dynamic motion of glossal surfaces. To confirm this, a simple testcan be conducted by stimulating the antenna with nectar, the glossa do notprotract with hair erection every time (Goodman 2003). Future attention shouldfocus on the anatomy of the honey bee’s glossa and the biomechanics of thestructure morphing to find out how the honey bee coordinates different structures withinthe tongue to control the erection of the hairs accurately.
Acknowledgments We thank the Centre of Biomedical Analysis of Tsinghua University for their assistancewith the specimen processing and SEM image capturing. This study was funded by the National NaturalScience Funding of China (Grant no. 51475258) and a Research Project of the State Key Laboratory ofTribology under Contract no. SKLT11B03.
Author Contributions R. Z., H. L. and T. L. planned and performed the experiments. S. Y., Y. Y. and J. W.conceived the project. T. L. and H. L. analyzed the data, and J. W., S. Y., R. Z. wrote the paper.
Compliance with Ethical Standards
Competing Interests The authors declare no competing financial interests.
J Insect Behav (2016) 29:325–339 337

References
Abou-Shaara HF (2014) The foraging behavior of honey bees, Apis mellifera: a review. Vet Med 59:1–10Aronne G, Giovanetti M, Guarracino MR, Micco V (2012) Foraging rules of flower selection applied by
colonies of Apis mellifera: ranking and associations of floral sources. Funct Ecol 26:1186–1196Bauder JA, Lieskonig NR, Krenn HW (2011) The extremely long-tongued Neotropical butterfly Eurybia
lycisca (Riodinidae): proboscis morphology and flower handling. Arthropod struct dev 40:122–127Bertazzini M, Medrzycki P, Bortolotti L, Maistrello L, Forlani G (2010) Amino acid content and nectar choice
by forager honeybees (Apis mellifera L.). Amino Acids 39:315–318Briant TJ (1884) On the anatomy and functions of the tongue of the honeybee (worker). J Linnean Soc Lond,
Zool 17:408–417Cheer AYL, Koehl MAR (1987) Paddles and rakes: fluid flow through bristled appendages of small
organisms. J Theor Biol 129:17–39Chen J, Wu J, Yan S (2015) Switchable wettability of the honeybee’s tongue surface regulated by erectable
glossal hairs. J Insect Sci 15:164Cook AJ (1880) The tongue of the honey bee. Am Nat 14:271–280Corbet SA (2000) Butterfly nectaring flowers: butterfly morphology and flower form. Entomologia
Experimentalis et Applicata 96:289–298Crompton AW, Musinsky C (2011) How dogs lap: ingestion and intraoral transport in Canis familiaris. Biol
Lett 7:882–884Dade, H. A. (2009). Anatomy and dissection of the honeybee. pp. 17–22. Cardiff: International Bee Research
AssociationGoodman, L. D2003]. Form and function in the honey bee. International Bee Research Association.Hainsworth FR, Precup E, Hamill T (1991) Feeding, energy processing rates and egg production in painted
lady butterflies. J Exp Biol 156:249–265Harder LD (1986) Effects of nectar concentration and flower depth on flower handling efficiency of bumble
bees. Oecologia 69:309–315Harper CJ, Swartz SM, Brainerd EL (2013) Specialized bat tongue is a hemodynamic nectar mop. Proc Natl
Acad Sci 110:8852–8857Hilgert-Moreira SB, Nascher CA, Callegari-Jacques SM, Blochtein B (2014) Pollen resources and trophic
niche breadth of Apis mellifera andMelipona obscurior (hymenoptera, Apidae) in a subtropical climate inthe Atlantic rain forest of southern Brazil. Apidologie 45:129–141
Howell DJ, Hodgkin N (1976) Feeding adaptations in the hairs and tongues of nectar-feeding bats. J Morphol148:329–336
Jensen KH, Kim W, Holbrook NM, Bush JW (2013) Optimal concentrations in transport systems. J R SocInterface 10:20130138
Karolyi F, Colville JF, Handschuh S, Metscher BD, Krenn HW (2014) One proboscis, two tasks: Adaptationsto blood-feeding and nectar-extracting in long-proboscid horse flies (Tabanidae, Philoliche). ArthropodStruct Dev 43:403–413
Kim W, Gilet T, Bush JWM (2011) Optimal concentrations in nectar feeding. Proc Natl Acad Sci108:16618–16621
Kim W, Peaudecerf F, Baldwin MW, Bush JW (2012) The hummingbird’s tongue: a self-assembling capillarysyphon. Proc Natl Acad Sci 279:4990–4996
Kingsolver JG, Daniel TL (1983) Mechanical determinants of nectar feeding strategy in hummingbirds:energetics, tongue morphology, and licking behavior. Oecologia 60:214–226
Koehl MAR (2001) Transitions in function at low Reynolds number: hair-bearing animal appendages. Mathmethods applied sci 24:1523–1532
Krenn HW, Mauss V, Plant J (2002) Evolution of the suctorial proboscis in pollen wasps (Masarinae,Vespidae). Arthropod struct dev 31:103–120
Krenn HW, Plant JD, Szucsich NU (2005) Mouthparts of flower-visiting insects. Arthropod StructDev 34:1–40
Krenn HW (2010) Feeding mechanisms of adult Lepidoptera: structure, function, and evolution of themouthparts. Annu Rev Entomol 55:307
Lee SC, Lee SJ (2014) Uptake of liquid from wet surfaces by the brush-tipped proboscis of a butterfly. SciReport 4:6934
Lee SJ, Lee SC, Kim BH (2014) Liquid-intake flow around the tip of butterfly proboscis. J TheorBiol 348:113–121
338 J Insect Behav (2016) 29:325–339

Lehnert MS, Monaenkova D, Andrukh T, Beard CE, Adler PH, Kornev KG (2013) Hydrophobic–hydrophilicdichotomy of the butterfly proboscis. J R Soc Interface 10:20130336
Loudon C, Best B, Koehl M (1994) When does motion relative to neighboring surfaces alter the flow througharrays of hairs? J Exp Biol 193:233–254
Martínez del Rio C, Schondube JE, McWhorter TJ, Herrera LG (2001) Intake responses in nectar feedingbirds: digestive and metabolic causes,osmoregulatory consequences, and coevolutionary effects. Am Zool41:902–915
Monaenkova D, Lehnert MS, Andrukh T, Beard CE, Rubin B, Tokarev A, Lee W, Adler PH, Kornev KG(2012) Butterfly proboscis: combining a drinking straw with a nanosponge facilitated diversification offeeding habits. J R Soc Interface 9:720–726
Nilsson LA (1988) The evolution of flowers with deep corolla tubes. Nature 334:147–149Paul J, Roces F (2003) Fluid intake rates in ants correlate with their feeding habits. J Insect Physiol
49:347–357Prakash M, Quéré D, Bush JW (2008) Surface tension transport of prey by feeding shorebirds: the capillary
ratchet. Science 320:931–934Quéré D (1999) Fluid coating on a fiber. Annu Rev Fluid Mech 31:347–384Reis PM, Jung S, Aristoff JM, Stocker R (2010) How cats lap: water uptake by Felis catus.
Science 330:1231–1234Rico-Guevara A, Rubega MA (2010) The hummingbird tongue is a fluid trap, not a capillary tube. Proc Natl
Acad Sci 108:9356–9360Shimizu A, Dohzono I, Nakaji M, Roff DA, Miller DG III, Osato S, Yajima T, Niitsu S, Utsugi N, Sugawara
T, Yoshimura J (2014) Fine-tuned bee-flower coevolutionary state hidden within multiple pollinationinteractions. Sci Report 4:3988
Simpson J, Riedel I (1964) Discharge and manipulation of labial gland secretion by workers ofApis mellifera (L.)(hymenoptera: Apidae). Proceedings of the Royal Entomological Society ofLondon. Series A 39:76–82
Snodgrass, R. E. (1956). Anatomy of the Honey Bee. New York: Comestock Publishing Association, Ithaca.pp. 59–93
Snodgrass, R. E. (1984). Anatomy of the honey bee. New York: Cornell University Press. pp. 43–50Wilhelmi AP, Krenn HW (2012) Elongated mouthparts of nectar-feeding Meloidae (Coleoptera).
Zoomorphology 131:325–337Winter Y, von Helversen O (2003) Operational tongue length in phyllostomid nectar-feeding bats. J Mammal
84:886–896Wu, J., Zhu, R., Yan, S. and Yang, Y. (2015). Erection pattern and section-wise wettability of a honeybee's
glossal hairs in nectar feeding. The Journal of experimental biology jeb-111013Yang H, Wu J, Yan S (2014) Effects of erectable glossal hairs on a honeybee’s nectar-drinking strategy. Appl
Phys Lett 104:263701–263704
J Insect Behav (2016) 29:325–339 339