identification of immunogenic proteins from ovarian … · el grado de doctor en ciencia y...
TRANSCRIPT

“Identificación de nuevos antígenos con potencial inmunoprotector en tejidos de intestino y ovario de la garrapata del ganado Rhipicephalus
(Boophilus) microplus, a través de un enfoque inmunoproteómico”
TESIS
QUE PARA OBTENER
EL GRADO DE
DOCTOR EN CIENCIA Y TECNOLOGÍA
EN LA ESPECIALIDAD DE
BIOTECNOLOGÍA PRODUCTIVA
PRESENTA
M. en C. y T. Patricia Berenice Ramírez Rodríguez
GUADALAJARA, JAL. 02 de marzo 2017.
CENTRO DE INVESTIGACION Y ASISTENCIA EN TECNOLOGIA Y DISEÑO DEL ESTADO DE JALISCO, A.C.

I
DEDICATORIA
Quiero dedicar esta tesis de Doctorado a mi amada familia…
Mi Memo e hija María.
A mi madre que tanto amo, mi adorada hermana y a mis hermosos sobrinos.
Y finalmente a mi padre Antonio †

II
"El éxito puede estar tan plagado de peligros como el fracaso, porque abre más puertas y conlleva
más responsabilidades"
- "El sufrimiento no dura tanto como nuestros miedos quieren hacernos creer"
- "No creas arte después de convertirte en un artista. Te conviertes en artista al crear arte sin cesar"
- "El éxito consiste en el privilegio de trabajar más a profundidad, no de ganar"
-"La libertad no es la capacidad de hacer lo que te venga en gana. Es la voluntad de hacer lo que
quieres hacer"
-"Por desgracia, no existe una manera indolora de alcanzar los objetivos"
- "El fracaso es un acontecimiento, no una persona"
-"Tienes que salir ahí fuera para improvisar y tienes que superar el miedo a morir. Tienes que ser
capaz de arriesgarte a morir"
-"Tu mayor fracaso es aquello que soñabas hacer, pero que no hiciste porque no reuniste el valor
necesario"
* El engaño de Ícaro: ¿Qué tan alto quieres volar?
(The Icarus Deception: How High Will You Fly?,
Seth Godin, Portfolio Penguin, 2012)

III
AGRADECIMIENTOS
Este estudio fue apoyado por el Fondo Sectorial de Investigación para la Educación CB-
2008-01-104006, por el Fondo Institucional del CONACYT, Proyectos de Desarrollo
Científico para Atender Problemas Nacionales, 2013, PDCPN2013-01-216321 y por el
Fondo del CONACYT-Programa de Estancias Sabáticas Nacionales, Estancias al Extranjero
y Estancias Cortas para la Consolidación de Grupos de Investigación 2014-246310.
Agradezco al Consejo Nacional de Ciencia y Tecnología (CONACYT) por el financiamiento
de beca otorgado para sustentar mis estudios durante el Programa de Doctorado No. de apoyo
344858.
Dedico estas líneas a todas las personas que contribuyeron a que este trabajo se haya podido
realizar.
Comienzo por agradecer al doctor Moisés por su confianza, dedicación, apoyo y amistad. Y
por supuesto a mis asesores, con los cuales conté en todo momento y siempre estuvieron
exhortándome a mejorar y seguir adelante (Dr. Rodolfo, Dra. Sara y al Dr. Daniel).
De igual forma agradezco, al Dr. Eduardo, al Dr. Abel, al Dr. Emmanuel, a Rito, a mi amiga
Saray, a la familia Preciado que siempre nos recibió y atendió con amabilidad cuando se
realizaron los muestreos de garrapatas en Tapalpa.
A las personas de Inifap Morelos, que colaboraron en el proyecto. El Dr. Rodrigo, la Dra.
Delia, el Dr. Rodolfo y finalmente a Mtro. Edgar, gracias por todo el trabajo de campo y
además su afecto.
A mí familia por estar siempre conmigo y apoyarme en todas mis decisiones.
A los miembros del jurado: Dr. Moisés Martínez, al Dr. Eduardo Padilla, al Dr. Jorge Gaona,
al Dr. Emmanuel Díaz y finalmente a Dr. Miguel Vásquez. Les agradezco infinitamente su
tiempo dedicado.

IV
RESUMEN
A nivel global las garrapatas Rhipicephalus (Boophilus) microplus representan un grave
problema de salud para el ganado principalmente en zonas tropicales y subtropicales. Siendo
los vectores más importantes de algunas de las enfermedades trasmitidas al ganado y al
humano. Esto se traduce en grandes pérdidas económicas en la industria ganadera y el sector
salud. Por otra parte, el incremento en el desarrollo de la resistencia a los actuales acaricidas
y la subsecuente contaminación ambiental por el uso desmedido de ellos, han llevado a la
búsqueda de métodos alternativos de control, como la vacunación. A este respecto, las
vacunas son una estrategia viable de control de garrapatas, además de ofrecer la ventaja de
reducir el impacto ambiental y la disminución de los residuos tóxicos presentes en la carne
del ganado. Un ejemplo es la vacuna Gavac®, formulada con el antígeno recombinante
Bm86, una proteína presente en la membrana de las células del intestino en las garrapatas.
Esta vacuna se ha utilizado en varios programas de inmunización en el ganado, sin embargo,
los resultados obtenidos hasta el momento han mostrado diferentes niveles de eficacia en las
cepas de R. microplus, lo que podría atribuirse a variaciones antigénicas en la proteína diana.
Los intestinos y los ovarios de la garrapata desempeñan un papel clave en las funciones
fisiológicas y reproductivas del parásito, lo que los convierte en una fuente potencial para el
descubrimiento de antígenos protectores. Por esta razón, el objetivo del presente estudio fue
caracterizar proteínas inmunogénicas, a partir de tejido intestinal y ovárico, de garrapatas
hembras adultas ingurgitadas. Se aislaron proteínas totales de estos tejidos y se utilizaron
para inmunizar bovinos susceptibles, en condiciones controladas. El título de anticuerpos en
suero de los animales inmunizados se obtuvo mediante ensayos de ELISA; posteriormente,
las proteínas fueron analizadas por SDS-PAGE 1D y 2D y por la técnica de Western Blot.
Las proteínas reconocidas por los ensayos fueron identificadas mediante LC-MS/MS. Los
análisis obtenidos de las muestras dieron como resultado la identificación de diez proteínas
inmunogénicas, lo que representa una ventana de oportunidad para el descubrimiento de
nuevos candidatos a vacunas, de acción efectiva.
Palabras clave: Vacunas; Rhipicephalus (Boophilus) microplus; proteínas inmunogénicas.

V
ABSTRACT
Rhipicephalus (Boophilus) microplus ticks represent a serious health problem for livestock
globally, mainly in tropical and subtropical areas. Being the most important vectors of some
diseases transmitted to cattle and human. This results into large economic losses in the
livestock industry and the health sector. On the other hand, the increase in the development
of the resistance to current acaricides and the subsequent environmental contamination by
the excessive use of them, have led to the search for alternative control methods, such as
vaccination. In this regard, vaccines are a viable tick control strategy, as well as offering the
advantage of reducing the environmental impact and reducing the toxic residues present in
cattle meat. An example is the Gavac® vaccine, formulated with the recombinant Bm86
antigen, a protein present in tick’s midgut membrane cells. This vaccine has been used in
several livestock immunization programs; however, the results obtained so far have shown
different levels of efficacy for R. microplus strains, which could be attributed to antigenic
variations in the target protein. The intestines and ovaries of ticks play an important role in
the physiological and reproductive functions of the parasite, which makes them a potential
source for the discovery of protective antigens. For this reason, the objective of the present
study was to characterize immunogenic proteins, from intestinal and ovarian tissue, of
engorged adult female ticks. Total proteins were isolated from these tissues and used to
immunize susceptible cattle under controlled conditions. The serum antibody titer of the
immunized animals was obtained by ELISA assays. Subsequently, the proteins were
analyzed by SDS-PAGE 1D and 2D and by the Western Blot technique. The proteins
recognized by the assays were identified by LC-MS/MS. Tests obtained from samples
resulted in the identification of ten immunogenic proteins, representing a window of
opportunity for the discovery of new vaccine candidates with effective action.
Keywords: Vaccines; Rhipicephalus (Boophilus) microplus; Immunogenic proteins

2
Abreviaturas
CENID Centro Nacional de Investigación Disciplinaria CIATEJ Centro de Investigación y Asistencia en Tecnología y Diseño del
Estado de Jalisco CIR Centros de Investigación Regionales °C Centígrados Inifap Instituto Nacional de Investigaciones
Forestales, Agrícolas y Pecuarias cDNA Ácido desoxirribonucleico complementario DNA Ácido desoxirribonucleico DTT Ditiotreitol EDTA Ácido etilendiaminotetraacético F Factor de aclopamiento gi Identificador génico H3PO4 Ácido fosfórico HCl Ácido clorhídrico His Histidina HRP Peroxidasa de rábano picante IgG Inmunoglobulina G KH2PO4 Fosfato de potasio monobásico KCl Cloruro de potasio kDa Kilodaltones KH2PO4 Fosfato de potasio monobásico KOH Hidróxido de potasio LB Luria Bertani M Molar Mg Miligramo mL Mililitro Mm Milímetro mM Milimolar MPM Marcador de peso molecular µg Microgramo µL Microlitro µM Micromolar NaCl Cloruro de sodio Ng nanogramos PAGE Electroforesis en gel de poliacrilamida PAVET Parasitología Veterinaria Pb Pares de bases PBS Buffer salino de fosfatos PCR Reacción en cadena de la polimerasa RT-PCR Reacción en cadena de la polimerasa en tiempo real pI Punto isoeléctrico PSA Persulfato de amonio RNA Ácido ribonucleico iRNA Ácido ribonucleico de interferencia

3
Rpm Revoluciones por minuto SDS Dodecil sulfato de sodio Spp Sin especies específicas TAE Tris-acetato-EDTA TBS Tris buffer salino TEMED Tetrametiletilendiamina UV Ultravioleta Vit.Rm3conceptual Vitelogenina Rhipicephalus microplus 3 conceptual VitRm_NF1 Vitelogenina Rhipicephalus microplus_Amino Forwar 1 VitRm_NR2 Vitelogenina Rhipicephalus microplus_Amino Reverse2 VitRm_MF1 Vitelogenina Rhipicephalus microplus_Medio Forwar 1 VitRm_MR2 Vitelogenina Rhipicephalus microplus_Medio Reverse2 VitRm_CF2 Vitelogenina Rhipicephalus microplus_Carboxilo Forwar 1 VitRm_CR2 Vitelogenina Rhipicephalus microplus_Carboxilo Reverse2 Rm-Vg-1 Rhipicephalus microplus Vitelogenina 1 Rm-Vg-2 Rhipicephalus microplus Vitelogenina 2 Rm-Vg-3 Rhipicephalus microplus Vitelogenina 3 1D Una dimensión 2D Dos dimensiones

4
INDICE
DEDICATORIA I AGRADECIMIENTOS II RESUMEN III ABREVIATURAS IV ÍNDICE V ÍNDICE DE TABLAS ÍNDICE DE FIGURAS DEDICATORIA 1. INTRODUCCIÓN 10 2. ANTECEDENTES 13
2.1. Importancia de las garrapatas 13 2.2. Importancia económica 14 2.3. Ciclo de vida 15 2.4. Distribución geográfica 16 2.5. Control químico natural 17 2.6. Hospederos resistentes 18 2.7. Depredadores naturales 19 2.8. La rotación, descanso y quema de praderas 19 2.9. La composición y tipo de vegetación 20 2.10. Control biológico 20 2.11. Resistencia a ixodicidas 21 2.12. Mecanismos de resistencia a acaricidas en R. microplus 22 2.13. Control Inmunológico 26
2.13.1. Control de infestaciones de garrapatas mediante vacunación 26 2.13.2. Antígenos expuestos y ocultos 27 2.13.3. La vacuna Gavac 27 2.13.4. Identificación de nuevos antígenos 29
3. JUSTIFICACIÓN 33 4. HIPÓTESIS 34 5. OBJETIVOS 35
5.1. Objetivo general 35 5.2. Objetivos específicos 35
6. DISEÑO EXPERIMENTAL 36 7. MATERIAL Y MÉTODOS 37
7.1. Garrapatas 37 7.2. Disección de tejidos y extracción de proteínas 37 7.3. Electroforesis unidimensional 38 7.4. Bovinos 39 7.5. Inmunización 40 7.6. Prueba ELISA 41 7.7. Electroforesis bidimensional 42 7.8. Ensayos Western blot 43 7.9. Análisis de LC-MS/MS 43 7.10. Aislamiento de ARN y síntesis de cDNA 45 7.11. Síntesis de cDNA 45

5
7.12. Análisis bioinformático 45 8. RESULTADOS 46
8.1. Cuantificación de las proteínas 46 8.2. Títulos de anticuerpos en bovinos vacunados 47 8.3. Detección de proteínas inmunogénicas mediante ensayos de
Western blot 49
8.4. Proteínas identificadas por LC-MS/MS 52 8.4.1. 1-Int-Sol: cadena pesada de la IgG 1 [Bos taurus] 52 8.4.2. 1-Int-Insoluble: Componente TF-II D del factor de iniciación
de la transcripción putativo 52
8.4.3. 2-Int-Insoluble: Subunidad alfa de la ATP sintasa tipo F0F1 putativa
52
8.5. 1-Ova-Soluble, 2-Ova-Soluble: Vitelogenina 53 8.5.1. 3-Ova-Soluble: Precursora de la Vitelogenina-2 53
8.6. 4a-Ova-Soluble, 4b-Ova-Soluble: Catepsina de yema 53 8.7. 1-Ova-Insoluble: Canal aniónico selectivo mitocondrial
dependiente de voltaje 53
8.8. 2-Ova-Insoluble: Serpina 54 8.9. 3-Ova-Insoluble: β-actina 54 8.10. Evidencia de múltiples vitelogeninas en R. microplus. 54 8.11. Productos de PCR´s en geles de agarosa al 1% de la especie
R. microplus en secuencias que codifican para las proteínas vitelogenias de ovario
60
9. DISCUSIÓN 62 9.1. Vitelogeninas en garrapatas 64
10. CONCLUSIÓN 66 11. PERSPECTIVAS 67 12. BIBLIOGRAFÍA 69 13. APENDICE 78
13.1. Electroforesis de Proteínas SDS-PAGE 1D 78 13.2. Protocolo para geles de separación y concentración de
proteínas en geles de poliacrilamida. 80
13.3. Marcadores de peso molecular para SDS-PAGE Tris-glicina 81 13.4. Carga de muestra apropiada para tinción con Coomassie y
plata para tiras IPG de 7cM 82
13.5. Isoelectroenfoque 82 13.6. Protocolo de corrida del isoelectroenfoque 83 13.7. Segunda Dimensión 84 13.8. Western Blot 84 13.9. Revelado de western blot con el método de peroxidasa 86 13.10. Soluciones 86
13.10.1. Solución amortiguadora de PBS pH 7.4 (Phosphate buffer saline)
86
13.10.2. Acrilamida al 30% /bisacrilamida al 0.8% 86 13.10.3. Tris-Cl/SDS 4X, pH 8.8 (1.5 M Tris-Cl, 0.4% SDS) 86

6
13.10.4. Tris-HCl/SDS 4X, pH 6.8 (0.5 M Tris-HCl, 0.4% SDS) 86 13.10.5. Persulfato de amonio al 10% 87 13.10.6. SDS-amortiguador 2X 87 13.10.7. Amortiguador de muestra 1X (BM 1X) (Muestras sólidas) 87 13.10.8. Amortiguador de muestra 2X (BM 2X) (Muestras líquidas) 87 13.10.9. Amortiguador de electroforesis 5X(Stock) 87 13.10.10. Amortiguador de electroforesis 1X. 87 13.10.11. Solución Colorante de proteínas (Azul brillante de
Coomassie) 88
13.10.12. Solución Decolorante de geles de poliacrilamida 88 13.10.13. Solución Equilibradora SDS de 2D 89 13.10.14. Amortiguador de lisis para 2D 89 13.10.15. Amortiguador de Transferencia 89
13.11. ELISA 90 13.12. Tablas con información acerca del material, reactivos,
equipos y kit utilizados en la tesis. 91
14. ANEXOS 95 14.1. Anexo 1. Tabla 1. Datos de animales con los que se
trabajaron en INIFAP, así como los grupos que se realizaron con estos mismos (tabla 2).
95
14.2. Anexo 2. De las proteínas que se identificaron mediante LC-MS/MS se realizó su análisis para llegar a su peso molecular, punto isoelétrico y nombre, los cuales se muestran a continuación.
95
14.2.1. Proteína 1-Int-Sol 95 14.2.2. Análisis in silico de la proteína de garrapata de 1-intestino
insoluble 96
14.2.3. Proteína 1-Int-Insol 96 14.3. Anexo 3. Análisis in silico de la proteína de garrapata de 2-
intestino insoluble 98
14.3.1. Proteína 2-Int-Insol 98 14.4. Anexo 4. Análisis in silico de la proteína de garrapata de 1-
ovario insoluble 101
14.4.1. Proteína 1-Ova-Insol 101 14.5. Anexo 5. Análisis in silico de la proteína de garrapata de 2-
ovario insoluble 103
14.6. Anexo 6. Análisis in silico de la proteína de garrapata de 3-ovario insoluble
105
14.7. Anexo 7. Análisis in silico de la proteína de garrapata de 4-ovario insoluble
107
14.8. Anexo 8. Proteínas de ovario solubles (tabla 3) 109

7
ÍNDICE DE TABLAS Tabla 1. Plantas que se han utilizado para el control de garrapatas. 18 Tabla 2. Ejemplos de algunos agentes biológicos que pueden ser usados para
el control de garrapatas. 20
Tabla 3. Características de la resistencia a acaricidas. 24 Tabla 4. Proteínas inmunogénicas reportadas. 31 Tabla 5. Grupos experimentales para el protocolo de inmunización. 39 Tabla 6. Muestras de proteínas y su concentración. 46 Tabla 7. Proteínas identificadas mediante análisis LC-MS/MS. 51 Tabla 8. Secuencia de péptidos para mandar a sintetizar. 67 Tabla 9. Programas que se utilizaron para analizar y diseñar los péptidos
sintéticos. 68

8
ÍNDICE DE FIGURAS
Figura 1. Fotografía de garrapata hembra adulta R. microplus parte dorsal. Aum.20x. Foto: Jaqueline Matías.
14
Figura 2. Esquema del ciclo biológico de la garrapata R. microplus. 16 Figura 3. Mapa de la distribución de la garrapata R. microplus en México,
según lo publicado en la campaña nacional para el control de la garrapata por SENASICA 2016.
17
Figura 4. A) Aplicación de acaricida mediante bomba de aspersión. Tapalpa, Jalisco 2014.
22
Figura 5. Esquema del desarrollo de la resistencia a los garrapaticidas. 23 Figura 6. Esquema de los factores involucrados en la respuesta inmune. 27 Figura 7. Inmunidad inducida por la vacuna del antígeno oculto Bm86. 29 Figura 8. A) Imagen de un bovino en donde se mantiene la cepa de la garrapata
de R. microplus.
37
Figura 9. A) Larvas de garrapata R. microplus. 38 Figura 10. A) Disección de garrapatas. 38 Figura 11. A) Garrapata diseccionada, en la cual se observan los tejidos de
intestino. 40
Figura 12. Esquema de inmunización e infestación de bovinos. 40 Figura 13. Esquema de sangrado de bovinos. 40 Figura 14. Esquema general de la prueba de ELISA con los sueros preinmunes
y de los bovinos inmunizados. 42
Figura 15. Determinación de los valores de absorbancia de la proteína BSA en diferentes concentraciones.
46
Figura 16. Patrón electroforético de proteínas de intestino e inmunodetección en 1D
47
Figura 17. Patrón electroforético de proteínas de ovario e inmunodetección en 1D.
48
Figura 18. Respuesta de anticuerpos en bovinos inmunizados. 48 Figura 19. Detección de proteínas inmunogénicas de R. microplus en 2D. (A)
Las proteínas solubles de intestino se sometieron a electroforesis bidimensional SDS-PAGE (12%, pH 3 a 10) y se tiñeron con azul de Coomassie.
49
Figura 20. Detección de proteínas inmunogénicas de R. microplus en 2D. (A) Las proteínas insolubles de intestino se sometieron a electroforesis bidimensional SDS-PAGE (12%, pH 3 a 10) y se tiñeron con azul de Coomassie.
50
Figura 21. Detección de proteínas inmunogénicas de R. microplus en 2D. (A) Las proteínas solubles de ovario se sometieron a electroforesis bidimensional SDS-PAGE (12%, pH 3 a 10) y se tiñeron con azul de Coomassie.
50
Figura 22. Detección de proteínas inmunogénicas de R. microplus en 2D. (A) Las proteínas insolubles de ovario se sometieron a electroforesis bidimensional SDS-PAGE (12%, pH 3 a 10) y se tiñeron con azul de Coomassie.
51

9
Figura 23. Alineación de la secuencia de aminoácidos deducida de Vitellogenin 1 (JAC58991.1), Vitellogenin 2 (Vg-3, JAC58954.1) y Vitellogenin 3 (Vg-2, JAC58956.1) de R. microplus.
58
Figura 24. Vitelogeninas en R. microplus. A) Comparación esquemática de las tres Vitelogeninas de R. microplus y sus dominios característicos.
59
Figura 25. Expresión de vitelogeninas en los ovarios de R. microplus. Se realizaron reacciones de RT-PCR a partir del RNA total de los ovarios de las garrapatas hembras adultas injurguitadas.
60
Figura 26. Esquema de predicción de péptidos antigénicos de la proteína vitelogenina 2 de R. microplus.
67

10
1. INTRODUCCIÓN
Las garrapatas del ganado Rhipicephalus (Boophilus) microplus (R. microplus) (Miller,
Davey y col., 2005), son ectoparásitos hematófagos obligados, clasificados en la clase Acari,
que se distribuyen en las regiones tropicales y subtropicales del mundo (De La Fuente y
Kocan 2006). Es el principal ectoparásito que afecta al ganado vacuno, ya que sirve como
vector para los protozoos Babesia bovis y Babesia bigemina, así como para la bacteria
Anaplasma marginale, que inducen enfermedades en el ganado.
La garrapata R. microplus se declaró erradicada de los EE.UU. en 1943 (Graham y Hourrigan
1977). Sin embargo, su reintroducción potencial por el ganado importado desde México
(Temeyer 2004) plantea una continua amenaza para la industria del ganado en los EE.UU.
La presencia de la garrapata ha aumentado recientemente, debido al hecho de que las
poblaciones mexicanas de garrapatas han desarrollado resistencia a los acaricidas que son
comúnmente utilizados para su control en el ganado de estas regiones (Temeyer 2004; Miller,
Davey y col., 2005). La infestación de los bovinos por garrapatas es una de las principales
causas de la baja productividad en la industria ganadera. Las hembras de R. microplus,
comienzan la ingestión rápida de sangre después de la fecundación, aumentan su volumen
corporal alrededor de 100 veces, lo cual puede inducir anemia en el bovino, así como
reducción en la producción de leche y carne, disminución de peso corporal, y eventualmente
una depreciación en el valor comercial de la piel (de La Fuente y Kocan 2006; Esteves,
Fogasa y col., 2009).
Las pérdidas económicas anuales, en la industria ganadera, debidas directa o indirectamente
a las enfermedades transmitidas por garrapatas se han estimado en más de 160 millones de
dólares en Australia, en aproximadamente 170 millones de dólares en Brasil y entre 2,000 a
3,000 millones de dólares en todo el mundo (FAO 2007). Por otro lado, se destinan más de
2 millones de dólares anualmente para el control de la garrapata en todo el mundo (Guerrero,
Nene y col., 2006). Debido a lo anterior es trascendental el control eficaz de las garrapatas,
para el continuo éxito de la industria ganadera tropical.

11
El control convencional que se utiliza para combatir a las garrapatas se basa en el uso de
acaricidas (Martins, Evans y col., 2002); sin embargo, el manejo indebido de estas moléculas
ha incrementado el desarrollo de garrapatas resistentes a los acaricidas (Crampton, Green y
col., 1999; Martins y Furlong 2001; Klafke, Sabatini y col., 2006; Parizi y col., 2009).
Además, este tipo de tratamientos químicos presenta severas limitaciones, ya que afecta la
inocuidad de los alimentos y agrava los problemas de contaminación ambiental. Debido a lo
cual, en las últimas décadas los investigadores han centrado su atención en la búsqueda de
nuevos métodos para el control de los ectoparásitos.
Como parte de esta búsqueda, se ha propuesto como una buena alternativa el método de
control inmunológico, el cual es capaz de inducir una respuesta tal en los animales
vacunados, que permite mantener controlados a los parásitos. Esta vía permitiría una
protección de mayor duración y está además prácticamente exenta de causar problemas de
índole ambiental, a diferencia del tratamiento con los acaricidas químicos (Rodríguez Valle
2000).
La vacuna comercial Gavac® (Cuba), se ha empleado con éxito para el control de la garrapata
R. microplus en ganado del sur de América. Ésta se compone de un antígeno recombinante
de la proteína Bm86, que es una proteína de la membrana del intestino de la garrapata (Rand,
Moore y col., 1989; Willadsen, Riding y col., 1989; Gough y Kemp 1993). Su protección
consiste en la inducción de anticuerpos que se unen a las células intestinales del ectoparásito
y provocan su muerte. Mediante el uso de esta vacuna se logra reducir hasta en un 60% la
aplicación de tratamientos contra la babesiosis, los cuales se reducen aún más (hasta un 82%)
con el adicional uso de acaricidas.
Para la transmisión de la Babesia, el intestino de las garrapatas juega un papel importante, ya
que es ahí donde, al beber la sangre como alimento, se ingieren los microbios, que entran así
en contacto con los tejidos internos de la garrapata (Untalan, Guerrero y col., 2005;
Rachinsky, Guerrero y col., 2008). Por otra parte, a partir de la digestión de la sangre la
garrapata obtiene los nutrientes necesarios para mantener activa la ovogénesis (Kongsuwan,
Josh y col., 2010). Por lo tanto, ya que tanto el intestino como los ovarios cumplen un papel
preponderante en la fisiología de la garrapata, si se logran identificar proteínas blanco tanto

12
de intestino como del ovario, que sean inmunogénicas, se podría bloquear o interrumpir la
producción de huevos de las garrapatas, para facilitar su control. Sin embargo, hasta la fecha
sólo un número limitado de proteínas de intestino de garrapata ha sido identificado. El
desarrollo de una vacuna recombinante a partir de proteínas inmunogénicas de intestino o de
ovario contra las garrapatas, representa en la actualidad una de las opciones más prominentes
para sustituir el control químico (Jonsson, Matschoss y col., 2000). Por esta razón, el objetivo
de este estudio fue caracterizar las proteínas inmunogénicas de tejidos de intestino y ovario
para identificar nuevas proteínas candidatas para la posterior elaboración de una vacuna
contra las garrapatas.

13
2. ANTECEDENTES
2.1. Importancia de las garrapatas
Las garrapatas son ácaros artrópodos, comprendidas en dos familias: Ixodidae o garrapatas
duras y Argasidae o garrapatas blandas (Barker y Murrell 2004). Debido a su gran capacidad
de adaptación y propagación, las garrapatas del género Rhipicephalus se extienden en
diversas áreas geográficas de todo el mundo (Estrada-Peña y Venzal 2006). De las cinco
especies que integran el género a nivel mundial, el Rhipicephalus (R. microplus) presenta
una mayor importancia por su amplia distribución en gran parte de América, África, Asia y
Australia (Nari y Fiel 1994, Rodríguez-Vivas y col., 2005, Taylor y col., 2007) (Fig. 1).
Por otra parte, las garrapatas R. microplus transmiten patógenos, los cuales provocan
encefalitis, en la enfermedad de Lyme, babesiosis (Babesia bovis y Babesia bigemina),
anaplasmosis (Anaplasma marginale) y teileriosis. Constituyen la principal plaga del ganado
en las regiones tropicales y subtropicales del mundo, lo cual impacta especialmente en la
producción del ganado, y afecta principalmente la ganancia de peso y la producción de leche
(Peter y col., 2005).
El control de las garrapatas con acaricidas químicos presenta graves inconvenientes, como el
desarrollo de la resistencia a los acaricidas en las garrapatas, su toxicidad, la contaminación
de los productos alimenticios, los riesgos para la salud humana y la contaminación ambiental
(Graf y col., 2004).

14
Garrapata del ganado
Clasificación científica
Reino: Animalia
Filo: Arthropoda
Clase: Arachnida
Subclase: Acari
Orden: Ixodida
Familia: Ixodidae
Género: Rhipicephalus
Subgénero:
Especie:
Boophilus
microplus
Figura 1. Fotografía de garrapata hembra adulta R. microplus parte dorsal. Aum.20x. Foto: Jaqueline Matías.
2.2. Importancia económica
R. microplus es responsable de pérdidas económicas directas, al alimentarse de sangre y
causar anemia e irritación de la piel, así como de pérdidas indirectas, al mantener y propagar
agentes patógenos, como los protozoarios Babesia bovis y Babesia bigemina, las rickettsias
Anaplasma marginale y Anaplasma centrale, y diversas enfermedades virales, todas ellas
causantes de mortalidad en bovinos (Avendaño y Correa, 2002; Baxter y Barker, 2002; Peter
y col., 2005; Walker, 2009; Francischetti y col., 2010; Kaufman, 2010).
La pérdida de peso de un bovino parasitado por garrapatas del género R. microplus se calcula
en 0.26 kg/garrapata/año y se ha observado que animales infestados con garrapatas reducen
su consumo de alimento (4.37 kg) en comparación con animales no expuestos a garrapatas
(5.66 kg). Estos efectos ocasionan pérdidas de varios miles de millones de dólares en la
economía pecuaria mundial (Rodríguez-Vivas y col., 2005, Jonsson 2006).

15
2.3. Ciclo de vida
El ciclo de vida de R. microplus se desarrolla en dos fases:
a) La fase parasitaria sobre el bovino
b) La fase de vida libre fuera del bovino
El periodo de vida libre comprende cuatro etapas: preoviposición, oviposición, incubación y
el periodo de sobrevivencia de las larvas sobre el pasto. El periodo de vida libre comprende
desde que una hembra ingurgitada se desprende del bovino y cae al suelo, hasta que las larvas
de su progenie logran parasitar un nuevo hospedero.
La fase parasitaria, pasa de larvas a metalarvas y de ninfas a metaninfas, posteriormente en
neóginas, partenóginas y teleóginas. Para el caso de los machos, pasan a meandros y a
gonandro. Los machos pueden permanecer sobre los bovinos durante periodos de hasta 90
días. Todas las etapas de la fase parasitaria se llevan a cabo en el mismo animal, por lo cual
se llama garrapata de un hospedero, para diferenciarla de otras garrapatas que necesitan 2 o
3 hospederos para completar su vida parasitaria, como ocurre con otras especies, como R.
bursa y Amblyomma.
Las dos especies del género Rhipicephalus spp., reportadas en México (R. annulatus y R.
microplus), así como Anocentor (Dermacentor spp.) nitens, la garrapata tropical de los
caballos, son ejemplos clásicos de garrapatas de un hospedero, es decir, pasan las tres fases
de su ciclo evolutivo parasitario (larva, ninfa y adulta) en la piel de un mismo animal.
1. La vida parasitaria de la garrapata Rhipicephalus spp. sobre el bovino es generalmente
de tres semanas, que incluyen dos mudas (de larva a ninfa y de ninfa a adulta). Las
hembras fecundadas y repletas de sangre se caen del animal hospedero (bovino) y
depositan entre 2,000 y 3,000 huevecillos, en lugares protegidos en el suelo, de los que,
dependiendo del clima, nace una nueva generación de larvas en un lapso de 6 a 8 semanas.
La hembra muere después de la oviposición.
2. Estas larvas, apenas perceptibles a simple vista, se mueven con sus 6 patas, trepan hierbas
y arbustos, y esperan a que pase algún animal que les sirva de hospedero.
3. Con sus órganos bucales se adhieren a la piel, la perforan, chupan sangre y líquido
corporal hasta repletarse para luego mudar al estadio de ninfa. La ninfa con cuatro pares

16
de patas vuelve a chupar sangre y pasa una segunda muda para convertirse en garrapata
adulta de sexo diferenciado.
4. Luego de la copulación, las hembras fecundadas y llenas de 0.3 a 0.5 ml de sangre se
caen del animal hospedero y así comienza el nuevo ciclo con la puesta de los huevos y la
muerte de la hembra (Figura 2).
Figura 2.-Esquema del ciclo biológico de la garrapata R. microplus. 1) Hembras repletas de sangre se dejan caer al suelo, 2) Puesta de huevos en el suelo, 3) Larva al salir del huevo, 4) Larva se adhiere al hospedero, 5) Larva repleta muda a ninfa en el hospedero, 6) Ninfa repleta muda a garrapata adulta sobre el hospedero y 7) Fecundación de las hembras antes de la succión de sangre.
2.4. Distribución geográfica
La garrapata común del ganado, R. microplus se encuentra distribuida en las regiones
tropicales y subtropicales del mundo. Estas garrapatas son endémicas de la India, Asia,
Australia, Madagascar, África, el Caribe y México (Estrada-Peña y col., 2006). En la
ganadería bovina mexicana las especies de garrapatas que tienen mayor importancia son: R.
microplus, R. annulatus, Amblyomma cajennense, Amblyomma maculatum, Amblyomma
americanum y Anocentor nitens. Sin embargo, las especies de mayor importancia para el
ganado bovino en México son R. microplus y Amblyomma cajennense. A. cajennense tiene
un área de distribución que cubre una superficie aproximada de 609, 857 Km2, lo que
representa el 31% del territorio nacional, mientras que R. microplus presenta un área de

17
distribución que abarca zonas tropicales, templadas y áridas, que cubren en conjunto 1,
043,772 Km2, lo que constituye el 53.0% del territorio nacional.
La distribución, desarrollo y reproducción de las garrapatas obedece a las condiciones
medioambientales, como la humedad relativa, la temperatura, la vegetación y la
disponibilidad de hospederos (Estrada-Peña y col., 2006). La Campaña Nacional contra la
garrapata Boophilus spp. establece 3 fases (Figura 3). La fase libre (verde) representa una
porción importante del norte del país y una pequeña área en el centro; comprende 94.4
millones de hectáreas que equivalen al 47.88% del territorio nacional. Las áreas de
erradicación cuentan con 1.1 millones de hectáreas; en estas regiones el parásito ha sido
eliminado por efectos de la Campaña y representan un 0.57% del territorio. Finalmente, las
áreas en Fase de Control alcanzan al momento una superficie total de 101.6 millones de
hectáreas y representan el 51.5% del país (www.senasica.gob.mx).
Figura 3. Mapa de la distribución de la garrapata R. microplus en México, según lo publicado en la campaña nacional para el control de la garrapata por SENASICA 2016.
2.5. Control químico natural Un método de control químico natural se basa en el uso de extractos de plantas (Sardá-Ribeiro
y col., 2007). Algunos estudios que se han realizado con extractos, como métodos de control
contra garrapatas, se muestran en la Tabla 1.

18
Tabla 1. Plantas que se han utilizado para el control de garrapatas.
Extracto de planta
Especie de garrapata
Estadio de la garrapata a la cual
afecta
Porcentaje de control
Autor
Calea serrata R. microplus y Rhipicephalus
sanguineus
Reducción en la oviposición. Mortalidad en larvas.
11 al 14 % 100%
Sardá-Ribeiro y col., 2007.
Nicotiana tabacum
R. sanguineus Mortalidad en larvas. En ninfas En ninfas
70% 61.79 % 64.9 %
Neira y col., 2009.
Azadirachta indica
Amblyomma
variegatum
Hyalomma anatolicum excavatum
Larvas Todos los estadios
No reportado No reportado
Pathak y col., 2004.
Azadirachta indica y Ocimum
R. appendiculatus Todos los estadios No reportado
Ndumu y col., 1999, Kaaya y Hassan 2000.
45 extractos metanólicos de plantas.
Petiveria
alliacea
R. microplus Larvas En adultas
5-99 % 86 % y 91%
Rosado-Aguilar y col., 2010 a,b.
Cuminum cyminum y Pimenta dioica
R. microplus Larvas 100 % Martínez-Velázquez y col., 2011a.
Lippia graveolens Ros-marinus officinalis
Allium sativum
R. microplus Larvas 85 a 100 % Martínez-Velázquez et col., 2011b.
Cuatro plantas tropicales ricas en taninos
R. microplus Larvas No reportado
Fernández-Salas y col., 2011.
2.6. Hospederos resistentes
Las razas de ganado Bos indicus son más resistentes a las garrapatas que las razas B. taurus.
Jonsson (2006) menciona que el ganado B. indicus presenta del 10 al 20 % menos garrapatas
que el ganado B. taurus. La resistencia a las garrapatas varía en relación al sexo, edad, estado
de gestación y lactación y temporada del año (Utech y col., 1978). La cría de ganado
resistente a las garrapatas se puede lograr por la selección de animales que presenten menos
garrapatas, y su posterior cruzamiento con otros animales con la misma característica, así

19
como introducir sangre cebú en el hato (Rodríguez-Vivas y col., 2005). En cruzas de ganado
B. indicus la heredabilidad del número de garrapatas R. microplus es alta, el total de la
proporción de la variación genotípica se debe al efecto aditivo de los genes (Mackinnon y
col., 1991).
2.7. Depredadores naturales
En México y en América Latina existen algunas garzas y pájaros que son depredadores
naturales de garrapatas. También existen algunas especies de hormigas con efecto depredador
en la población de garrapatas (Wilkison 1970, Verrisimo y Machado 1995, Ojeda-Chi y col.,
2011).
2.8. La rotación, descanso y quema de praderas
El sistema de rotación se basa en descansos obligados de las praderas, con la finalidad de
presionar a las garrapatas en su etapa de vida libre, al impedir o retardar que como larvas
activas encuentren a su hospedero, para que mueran por hambre y deshidratación (Wilkinson
1970). Furlong (1998) reportó que el tiempo de descanso para reducir el número de larvas
presentes en las praderas debe ser de 45-60 días. En Venezuela el empleo de la modelación
por computadora permitió predecir que al emplear una rotación de praderas de 36 días de
descanso en época de secas y 24 días en época de lluvias permite una reducción sustancial
de las infestaciones de garrapatas (FAO 2005). El fuego afecta directamente a las garrapatas
por la exposición que sufren a las altas temperaturas en los estadios de larvas, hembras
adultas y huevos (Rodríguez-Vivas y col., 2005). Indirectamente tiene un efecto por la
destrucción de la capa de vegetación que le sirve de protección a las garrapatas (FAO 2005).
Sin embargo, después de un estudio realizado en California, EUA para evaluar el efecto del
fuego sobre las poblaciones de garrapatas, los autores concluyeron que a pesar de las quemas
prescritas en los hábitats, esto no se tradujo en una disminución de la abundancia de adultos
ixódidos sobre la vegetación, y al parecer no afecta a las garrapatas argásidas e ixódidas que
se refugian en suelo (Padgett y col., 2009).

20
2.9. La composición y tipo de vegetación
Existen leguminosas con capacidad para atrapar larvas, mediante pelos y secreciones
glandulares viscosas presentes en sus hojas y que tienen la capacidad de inmovilizar entre un
12-27 % de larvas de R. microplus (Wilson y col. 1989). En México, se ha evaluado el efecto
de las gramíneas forrajeras: Melinis minutiflora y Andropogon gayanus, las cuales poseen la
capacidad de repeler, atrapar u obstaculizar a las garrapatas que buscan hospedero, y pueden
reducir el riesgo del encuentro garrapata-bovino (Cruz-Vazquez y Fernández-Ruvalcaba
2000, Fernández-Ruvalcaba y col., 2004).
2.10. Control biológico
Los agentes biológicos que potencialmente pueden ser usados para el control de garrapatas
se describen en la Tabla 2.
Tabla 2. Ejemplos de algunos agentes biológicos que pueden ser usados para el control de garrapatas. Hongos
entomopatógenos Bacterias Nemátodos Hormigas Estadio de la
garrapata al cual afecta
Autor
Metarhizium sp Beauveria sp
Cedecea -
lapagei
Heterorhabditi-dae
Solenopsis -germinata
Afectan los estadios de vida libre
Ojeda-Chi y col., 2011
Especie de garrapata
Bacterias Nemátodos Hormigas Estadio de la garrapata al cual afecta
Autor
R. microplus Escherichia
coli
Enterobacter
aglomerans
Steinernematidae S. saevissima
Camponotus rengira
Ectatomma
cuadridens
Afectan los estadios de vida libre
Fernandes y col., 2012
Todos estos agentes afectan principalmente los estadios de vida libre de las garrapatas
(Fernandes y col., 2012). En los últimos cuatro años se han desarrollado unos 171
micopesticidas alrededor del mundo, de los cuales únicamente tres productos contienen M.
anisopliae para el control de garrapatas (Fernandes y col., 2012).

21
2.11. Resistencia a ixodicidas
La continua aplicación de pesticidas en un intento por controlar las altas infestaciones de
garrapatas R. microplus ha resultado en el desarrollo de resistencia a los químicos en esta
especie de garrapata (Benavides, 2008). Durante las últimas cuatro décadas, el desarrollo de
productos garrapaticidas de gran eficacia y poder residual permitió al ganadero disponer de
una herramienta de control práctica y adaptable a diferentes sistemas de producción. Estas
características, sumadas a una disminución de la toxicidad hacia los humanos en los más
modernos grupos químicos, crearon un falso sentido de seguridad en el productor pecuario,
quien sustituyó el diagnóstico y el asesoramiento profesional por la casi exclusiva utilización
de fármacos. Sin embargo, el desarrollo paulatino de la resistencia parasitaria en el ámbito
mundial ha demostrado que los antiparasitarios son un recurso necesario, pero no renovable,
en la medida que la resistencia se extiende y persiste en las poblaciones parasitarias
(Organización de las Naciones Unidas para la Agricultura y la Alimentación [FAO], 2003;
Nari, 2011).
Los químicos ampliamente utilizados para el control de garrapatas han sido, arsenicales,
organoclorados, organofosforados, carbamatos, formamidinas, piretroides, lactonas
macrocíclicas y fenilpirazolonas (Peter y col., 2005, Baffi y col., 2008; Rodríguez-Vivas y
col., 2010, Prullage y col., 2011). La estrategia mundial más utilizada consiste en la
aplicación de ixodicidas sobre el cuerpo de los animales infestados, con intervalos de tiempo
determinados por la región ecológica, por las especies a combatir y por la eficacia residual
del ixodicida (Rodríguez-Vivas y col., 2008). Bajo esta estrategia, el método más utilizado
para aplicar los acaricidas es el de aspersión con bomba de espalda (Benavides y Romero,
2001; Consejo Nacional de Ciencia y Tecnología-Fondo Sectorial de Investigación en
Materia Agrícola, Pecuaria, Acuacultura, Agrobiotecnología y Recursos Fitogenéticos
[Conacyt-Sagarpa], 2005; Baffi y col., 2007), los cuales tienen como efecto común en la
garrapata alterar la transmisión del impulso nervioso en la unión sináptica mediante
diferentes mecanismos de acción que afectan los neurotransmisores o los canales iónicos que
intervienen en ella (Figura 4).

22
A B
Figura 4. A) Aplicación de acaricida mediante bomba de aspersión. Tapalpa, Jalisco 2014, fotografía tomada por Patricia Berenice Ramírez Rodríguez B) Esquema del mecanismo de transmisión del impulso nervioso en garrapatas, tomado de Díaz (2012).
2.12. Mecanismos de resistencia a acaricidas en R. microplus
Resistencia se define como la habilidad de una población de parásitos para tolerar dosis de
tóxicos que serían letales para la mayoría de individuos en una población normal
(susceptible) de la misma especie, es un mecanismo defensivo del parásito a nivel molecular
(Errecalde y col., 2003; Conacyt-Sagarpa, 2005; Badii y Garza, 2007). Es una modificación
genética que confiere a las poblaciones de artrópodos la capacidad de adaptarse a ambientes
tóxicos, una condición preadaptativa promovida naturalmente por la selección de genes
asociados a hidrólisis de componentes alelopáticos o artificialmente por la aplicación de
productos químicos (Rosario-Cruz y col., 2009) (Figura 5).
La resistencia a pesticidas es una característica hereditaria que implica cambios en el DNA,
generada por mutaciones en genes del sitio blanco de los acaricidas, como son los genes de
la acetilcolinesterasa en el caso de los organofosforados, del canal de sodio para los
piretroides, del receptor de octopamina para las formamidinas y del receptor GABA para las
avermectinas, o por mutaciones en los genes de enzimas multifuncionales tipo oxidasas,
hidrolasas y transferasas, que dan lugar a manifestaciones metabólicas relacionadas con sus
niveles de concentración o su actividad detoxificante (Heminway y col., 2004; Peter y col.,
2005). Inicialmente, los genes de resistencia o mutantes son raros en la población, pero por
selección continua aumentan su proporción en la medida que aumenta la población de

23
parásitos resistentes (Sangster, 2001). Esta situación se origina por el uso frecuente de
acaricidas con un mismo mecanismo de acción, lo cual lleva a que los individuos que no
presentan el alelo resistente hacia ese fármaco sean eliminados, mientras los que sí lo
presentan sobreviven, lo que permite transmitir este genotipo a su descendencia, con lo cual,
a través de generaciones posteriores, aumentan su frecuencia en la población.
Figura 5. Esquema del desarrollo de la resistencia a los garrapaticidas. Tomado del 1er. Simposium de Salud y Producción de Bovinos de Carne en la Zona Norte-Centro de México. Aguascalientes, Ags. Mayo 2010.
La resistencia a la penetración es una modificación del exoesqueleto para inhibir o retardar
la penetración del químico, y tiene que ver con la concentración de lípidos que facilitan o
retardan la penetración del pesticida a través de esta estructura (Alonso-Díaz y col., 2006).
El metabolismo incrementado se caracteriza por la detoxificación enzimática del
garrapaticida que es degradado a consecuencia de la acción de enzimas que participan en los
procesos de detoxificación, como son las oxidasas de función mixta, hidrolasas (esterasas,
carboxilesterasas) y glutatión S-transferasas (Conacyt-Sagarpa, 2005; Shahein y col., 2008).
La sensibilidad reducida se basa en la modificación que hace el ácaro del sitio blanco del
ixodicida, a fin de contrarrestar la toxicidad del principio activo del producto (He y col.,
1999). En R. microplus, las alteraciones en el metabolismo de degradación del principio
activo y los cambios en la estructura del sitio blanco parecen ser las respuestas principales
(Jamroz y col., 2000; Conacyt-Sagarpa, 2005; Chevillon y col., 2007b; Rosario-Cruz y col.,
2009) (Tabla 3).

24
Tabla 3. Características de la resistencia a acaricidas. Compuestos
Toxicidad Sitios blanco Referencias
Organofosforados (OP) Solubilidad: En agua, son miscibles en solventes orgánicos. Se encuentran como: Fosfatos, fosfonatos, fosforotioatos y fosforoditioatos.
Aguda Efectos SNC (cadena nerviosa ganglionar ventral) donde se ubican prácticamente todas las sinapsis colinérgicas, conducen a su parálisis o muerte.
Enzima acetilcolinesterasa, es fosforilada sobre el grupo hidroxilo de una serina en el sitio activo de la enzima. La acetilcolinesterasa inactiva bloquea la degradación del neurotransmisor acetilcolina, cuyas concentraciones sinápticas aumentan y generan hiperexcitación del SNC.
Cossio-Bayugar y col., 2002; Bloomquist, 2003; Li y col., 2003; Hawkes y col., 2005; Liming y col., 2006.
Carbamatos Solubilidad: En solventes orgánicos. Algunos carbamatos de naturaleza alifática pueden ser mezclables en agua
Toxicidad Aguda Efectos Hiperexcitabilidad, parálisis y muerte en artrópodos.
Actúan sobre el SNC y causan una reacción de carbamilación del grupo hidroxilo en una serina de la acetilcolinesterasa, que genera un grupo hidroxilado que migra e inactiva la enzima.
Bloomquist, 2003; Hawkes y col., 2005; Liming y col., 2006.
Piretroides sintéticos (PS) Solubilidad: Tienen un alto grado de lipofilia. Se encuentran como: Ésteres del ácido crisantémico.
Toxicidad Aguda Efectos Causan principalmente ataxia, convulsiones, descoordinación y, finalmente, la caída de la garrapata.
Canales de sodio voltaje dependientes, estructuras que comprenden cuatro dominios (I-IV) conformados por seis hélices transmembranales (S1–S6), donde los PS actúan para impedir o demorar su cierre. Esto retrasa el mecanismo normal de inactivación del impulso nervioso.
He y col., 1999; Bloomquist, 2003; Hemingway y col., 2004, Peter y col., 2005
Formamidinas Solubilidad:
Toxicidad Moderada Efectos La sobre estimulación ejercida por el amitraz sobre los receptores de
Estos compuestos imitan la acción de la octopamina, neurotransmisor que regula el comportamiento de excitación dentro del SNC, actúa también como una neurohormona sobre tejidos periféricos que induce la movilidad de lípidos y carbohidratos y como un neuromodulador central y periférico que actúa sobre
Bloomquist, 2003; Chen y col., 2007; Chevillon y col., 2007b; Prullage y col., 2011

25
Ligeramente solubles en agua, pero muy solubles en solventes orgánicos. Se encuentran como: Amitraz
octopamina origina su efecto tóxico en el SNC, e impide la acción reguladora de la octopamina, tanto del SNC como del periférico. Genera sobre estimulación de las sinapsis octopaminérgicas, y da lugar a temblores, convulsiones, anorexia, desprendimientos e interrupción de la reproducción.
los músculos, la córpora cardíaca y la córpora allata en los artrópodos; media su actividad a través de tres clases de receptores acoplados a proteínas G vinculadas a la adenilato ciclasa.
Lactonas macrocíclicas (LM) Solubilidad: Insolubles en agua pero altamente liposolubles. Se encuentran como: Avermectinas ivermectina, doramectina, abamectina y eprinomectina y las milbemicinas como la moxidectina y milbemicina oxima.
Toxicidad Acción endectocida, es decir, con efecto tóxico sobre ecto- y endoparásitos. Efectos Origina una parálisis flácida con el desprendimiento y muerte del parásito.
Se unen a canales de cloro regulados por glutamato, con lo que se impide el cierre de los canales y se aumenta la permeabilidad de este ion, lo que desencadena la hiperpolarización de la membrana y cesa el estímulo nervioso.
Bloomquist, 2003; Rodríguez-Vivas y col., 2010.
Fenilpirazoles Solubilidad: agua Se encuentran como: Fipronil
Toxicidad Posee alta toxicidad en artrópodos. Efectos Parálisis y posterior muerte del artrópodo.
Receptores del ácido gama-amino butírico (GABA). Se basa en el bloqueo de los canales de cloro que son controlados por GABA, a los cuales el fipronil se une en las membranas celulares de las neuronas de los invertebrados, e impide la acción inhibitoria del neurotransmisor y permite que continúe la entrada de iones cloruro.
Castro-Janer y col., 2010; Prullage y col., 2011.

26
2.13. Control Inmunológico
2.13.1. Control de infestaciones de garrapatas mediante vacunación
La vacunación se considera una alternativa ambientalmente segura y económicamente
sostenible para el control de los ectoparásitos. En un esfuerzo por minimizar las cantidades
de acaricidas actualmente utilizados, el desarrollo de vacunas contra la garrapata ha sido
priorizado en todo el mundo (Willadsen, 2004).
Los beneficios del uso de vacunas contra la garrapata, como parte de un programa de control
integrado, incluyen una reducción en el uso de acaricidas, lo cual prolonga la vida útil de los
acaricidas y retrasa el inicio de la resistencia, esto a la vez disminuye la incidencia de
enfermedades transmitidas por R. microplus y reduce los costos de producción (De La Fuente
y col., 2007; Rodríguez y col 1995a, b; De La Fuente y col., 1998).
La viabilidad de utilizar las proteínas de la garrapata como antígenos para la vacunación,
como método de control de garrapatas, se ha demostrado desde la introducción de dos
productos a mediados de la década de 1990; TickGARD, de Australia (Willadsen y col.,
1995) y Gavac ©, de Cuba (Canales y col., 1997). Ambos fueron desarrollados al utilizar la
glicoproteína Bm86 de los intestinos de las garrapatas R. microplus como antígeno
inmunorreactivo (de La Fuente y col., 2000; Willadsen y Kemp, 1988). TickGARD ya no
está comercialmente disponible pero Gavac © aún se maneja hasta la fecha, principalmente
regiones del sur de latinoamerica (Figura 6).

27
Figura 6. Esquema de los factores involucrados en la respuesta inmune. 1) Factores asociados con la inmunidad del hospedero. 2) Proteínas del parásito y el antígeno vacunal. 3) Vacunación con los antígenos, abreviatura: APC, Célula presentadora de antígenos (Figura tomada de Maritz-Olivier y col., 2012).
2.13.2. Antígenos expuestos y ocultos
Se han explorado dos tipos distintos de diana antigénica para el desarrollo de la vacuna. El
primero son los antígenos convencionales secretados en la saliva durante la fijación de la
garrapata y la alimentación en el hospedero, los llamados antígenos expuestos. Por lo general,
son proteínas o péptidos sintetizados en las glándulas salivales (Khalil y Wikel 1979;
Larregina y Falo 2005). Los antígenos ocultos están normalmente ocultos de los mecanismos
inmunes del hospedero (Willadsen y Kemp 1988). Regularmente se encuentran en la pared
del intestino de las garrapatas e interactúan con inmunoglobulinas específicas tomadas en la
comida de sangre. Sin embargo, cualquier antígeno que no se presenta normalmente al
hospedero es un antígeno oculto.

28
2.13.3. La vacuna Gavac
La vacuna Gavac fue lanzada en México en 1997 por Revetmex S.A. de C.V. (Ciudad de
México, México). Los ensayos de campo en México proporcionaron pruebas del efecto de la
vacunación con Bm86 recombinante para el control de infestaciones de R. microplus y R.
annulatus (Fragoso y col., 1998, de la Fuente y col., 1999, Redondo y col., 1999). Bm86
reduce el número de garrapatas hembras ingurgitadas, su peso y capacidad reproductiva. Por
lo tanto, el mayor efecto de la vacuna fue la reducción de infestaciones larvales en las
generaciones siguientes (Cobon y col., 1995, Rodríguez y col., 1995a, b; Willadsen y col.,
1995, Canales y col., 1997, de la Fuente y col., 1998, 1999, Redondo y col., 1999, Jonsson y
col., 2000a).
El antígeno Bm86 se expresa en el intestino de las garrapatas adultas hembras y los estudios
de silenciamiento de su expresión génica revelan que el Bm86 juega un papel importante
durante el período de alimentación y digestión de la sangre, en garrapatas hembras de R.
microplus repletas, alimentadas de bovinos infectados con Babesia bovis (Bastos y col.,
2010). Los anticuerpos anti-Bm86 aparentemente se unen a la superficie de las células
epiteliales del intestino de la garrapata e interrumpen la endocitosis, lo que causa la lisis de
las células y reduce la capacidad reproductiva de las hembras repletas (Willadsen 1997)
(Figura 7).
Entre los diversos efectos de la vacuna sobre la garrapata R. microplus se incluyen: la
reducción de la capacidad reproductiva (50-90 %), la reducción del número de garrapatas
repletas (20-30 %), así como del peso de las garrapatas (30 %) y del peso de los huevos (60-
80 %), no obstante, no produce mortalidad (Jonsson y col. 2000; De la Fuente y col., 2007).
También se ha demostrado que es eficaz contra otros géneros y especies de garrapatas tales
como B. annulatus, B. decoloratus, H. anatolicum y H. dromedarii (De la Fuente y col.,
2007). Se ha sugerido que el efecto de la vacuna Bm86 se podría incrementar con la inclusión
de otros antígenos efectivos o por el uso de adyuvantes. Sin embargo, las vacunas basadas en
Bm86 tienen una eficacia variable frente a diferentes cepas geográficas de R. microplus y no
afectan la capacidad vectorial de la garrapata (de la fuente y col., 1998).

29
Figura 7. Inmunidad inducida por la vacuna del antígeno oculto Bm86. Los elementos del sistema
inmunológico presentes en la sangre, penetran a través del capítulo y la boca de la garrapata hasta llegar
al intestino donde reaccionan con las moléculas blanco y provocan la inhibición de la función biológica de
1) Linfocitos; 2) anticuerpos anti-Bm86; 3) proteínas del complemento; 4) eritrocitos; 5) capítulo y boca; 6)
intestino; 7) acción conjunta de anticuerpos anti-Bm86 y complemento sobre las células del intestino de la
garrapata (Daniel Sojka y col., 2013; Carlos Bautista 1987).
2.13.4. Identificación de nuevos antígenos
Las vacunas contra la garrapata son una alternativa potencial a los métodos de control
químico, pero hasta ahora el pequeño número de antígenos de protección contra garrapatas
que se ha identificado, ha sido un paso limitante en el desarrollo de nuevas vacunas (de la
Fuente y Kocan, 2006). El avance de las vacunas ha sido lento. Muchos autores coinciden en
que el paso limitante es la identificación de nuevos antígenos con una función biológica
importante dentro del artrópodo (Mulenga y col., 2000; Willadsen y col., 2001).
Actualmente, existe una fuerte tendencia hacia el uso de la vacunología inversa para
identificar nuevos antígenos en ensayos de vacunación (Guerrero y col., 2012). La
disponibilidad del genoma de Ixodes scapularis y del genoma casi completo de R. microplus,

30
en combinación con la rápida expansión de los datos de transcriptómica, permite a los
científicos aventurarse en una era posgenómica para el diseño de vacunas contra la garrapata.
Otros intentos incluyen la identificación de nuevos antígenos, de otras especies de garrapatas,
que se espera contribuyan al control de garrapatas mediante el desarrollo de vacunas más
eficaces o vacunas polivalentes eficaces contra numerosas especies de garrapatas (Willadsen
y col., 2001, Rajput y col., 2006). Recientemente, se ha recurrido a la aplicación de métodos
de biología molecular, que incluyen transcriptómica y proteómica, y a la caracterización de
interacciones entre garrapatas (Rhipicephalus spp.) y patógenos (A. marginale o B.
bigemina), con análisis funcionales mediante RNA de interferencia (iRNA), para la
identificación de proteínas de garrapatas con un posible papel en la infección y en la
transmisión de los patógenos (Almazán y col., 2010; Antunes y col., 2012).
A partir del trabajo de varios grupos en todo el mundo, se ha identificado un número
considerable de vacunas candidatas (Parizi y col., 2012; Seixas y col., 2012). Sin embargo,
una baja antigenicidad y una protección limitada contra especies de garrapatas no
relacionadas, han mostrado la importancia del uso de antígenos adicionales.
También se han utilizado nuevos antígenos identificados en los ensayos de vacunación. Los
resultados de algunos de estos ensayos han sido decepcionantes (Parizi y col., 2009). Otros
estudios han mostrado candidatos atractivos para futuras evaluaciones (Almazan y col., 2012;
Hajdusek y col., 2010; Hajdusek y col., 2009; Parizi y col., 2011; Trimnell y col., 2005). Sin
embargo, se requieren análisis adicionales para caracterizar el potencial de estas proteínas
como antígenos vacunales, para el control de infestaciones por garrapatas e infección por
patógenos (de la Fuente 2013). Algunos de los antígenos que han sido utilizados como
candidatos para posibles vacunas contra la garrapata se muestran en la Tabla 4.
El avance real vendrá cuando estos desarrollos tecnológicos puedan desentrañar las
complejas interacciones entre las respuestas inmunes del hospedero y las garrapatas y
patógenos que transmiten (Nutall y col., 2006).

31
Tabla 4. Proteínas inmunogénicas reportadas.
Antígeno Especie de garrapata Comentarios Referencia
* Bm86 B. microplus Vacunas comerciales TickGARD/Gavac. Willadsen y col., 1995.
* Bm91 B. microplus Eficacia mayor que Bm86. Willadsen y col., 1996.
* Calreticulina A. americanum
D. variabilis
R. microplus
Baja inmunogenicidad en ganado. Jaworski y col., 1995. Ferreira y col., 2002.
IGBMC Proteína de adherencia a imunoglobulina
R. appendiculatus Proteína IGBMC de saliva específica para machos: Ligero efecto sobre la alimentación de las hembras en conejillos de indias.
Wang H y col., 1998.
Proteína de adherencia a Histamina
R. appendiculatus Reacción adversa en el modelo de conejillo de indias. Paesen GC y col., 1999.
P29 H. longicornis Tipo Colágeno; efectiva en contra de todas las garrapatas. Mulenga A y col., 1999.
HL 34 H. longicornis Proteína de la saliva; efecto adverso sobre la alimentación de las garrapatas en conejos.
Tsuda A y col., 2001.
RIM 36 R. appendiculatus Proteína de adherencia; muy antigénica pero no protege al ganado. Bishop R y col., 2002.
* 64TRPs glándula salival de garrapata
R. appendiculatus Acción dual mediante la reacción cruzada con antígenos ocultos y potencialmente con efectos universales sobre un gran número de especies de garrapatas.
Trimnell AR y col., 2002. Trimnell AR y col., 2005.
Péptidos sintéticos de Bm86
R. microplus Disminución de la población. Patarroyo y col., 2002.
Follistina de Ovario H. longicornis Disminución de ovogenesis y embriogénesis, reduce la tasa de reproducción.
Zhou y col., 2006.
* SBm7462 R. microplus Se busca la región conservada de 20 cepas de Brasil, Argentina, Colombia y Uruguay.
Peconick y col., 2008.
* FER2 transportador de hierro en la hemolinfa de garrapata
I. ricinus Se expresa en todas las etapas de crecimiento de la garrapata: silenciarlo por iRNA afecta la adquisición de sangre y reproducción.
Hajdysek y col., 2009.
* Bm95 H. longicornis La vacuna protegió a los animales de los retos de larvas, ninfas y adultas de garrapatas con una eficacia del 98,7%, 84,6% y 78,9%, respectivamente.
Sugumar y col., 2011.
* SUB-MSP1 R. microplus
R. annulatus
La vacuna mostró 60% de eficacia en la protección de ganado contra garrapatas.
Almazan y col., 2012.

32
* Serina proteasa R. microplus Reduce el número de la hembra adulta 83% en conejos en comparación al control.
Jittapalapong y col., 2012.
* Acuaporina (RmAQP1) R. microplus La proteína recombinante proporcionó una eficacia del 75% y 68% en la reducción del número de garrapatas femeninas adultas muestra en las infestaciones de R. microplus en bovinos.
Guerrero y col., 2014
* P0 proteína ribosomal R. sanguineus
R. microplus
Efecto adverso contra hembras adultas, durante la oviposición y número de huevos eclosionados en conejos.
Rodríguez y col., 2015.
Nota: * Proteínas de antígenos ocultos

33
3. JUSTIFICACIÓN
R. microplus es considerado el parásito más importante del ganado bovino, ya que reduce la
producción ganadera, produce pérdida de peso y disminución de la producción de leche en
los animales. Su mayor importancia radica en ser vector de enfermedades limitantes en el
trópico como la babesiosis y anaplasmosis. El control convencional que se utiliza para
combatir a las garrapatas se basa en el uso de acaricidas químicos. Sin embargo, el uso
indebido de estos ixodicidas ha incrementado la incidencia en la selección de garrapatas
resistentes y en la contaminación ambiental. Todo esto multiplica las dificultades de lucha
contra las garrapatas y muestra un futuro poco alentador en el uso de productos químicos
para la erradicación de los ectoparásitos. Las vacunas han demostrado ser un método factible
de control de las garrapatas, que ofrecen una solución rentable, y constituyen una alternativa
ecológica al control químico. Tienen la ventaja de ser específicas de especie, reducen la
incidencia de babesiosis y resultan inocuas para el hospedero bovino, el ser humano y el
medio ambiente, además que son fáciles de administrar. El desarrollo notable de la
inmunología molecular y la biotecnología, nos lleva a proponer la búsqueda de nuevos
antígenos con potencial vacunal, a partir de proteínas de intestino y de ovario de la garrapata
R. microplus.

34
4. HIPÓTESIS
Los bovinos vacunados con proteínas de tejidos de intestino y ovario de la garrapata R.
microplus desarrollan anticuerpos contra las mismas, lo que permite la identificación de
proteínas inmunogénicas con potencial inmunoprotector.

35
5. OBJETIVOS
5.1. Objetivo general
� Identificar nuevos antígenos con potencial inmunoprotector en tejidos de intestino y
ovario de la garrapata del ganado R. microplus mediante un enfoque
inmunoproteómico.
5.2. Objetivos específicos
� Evaluar la capacidad inmunogénica de los extractos de tejidos de intestino y ovario
de R. microplus en bovinos.
� Realizar la detección de proteínas inmunogénicas por medio de Western-Blot en 1D
y 2D con los sueros inmunes.
� Realizar la identificación de las proteínas inmunogénicas por medio de cromatografía
de líquidos/espectrometría de masas.
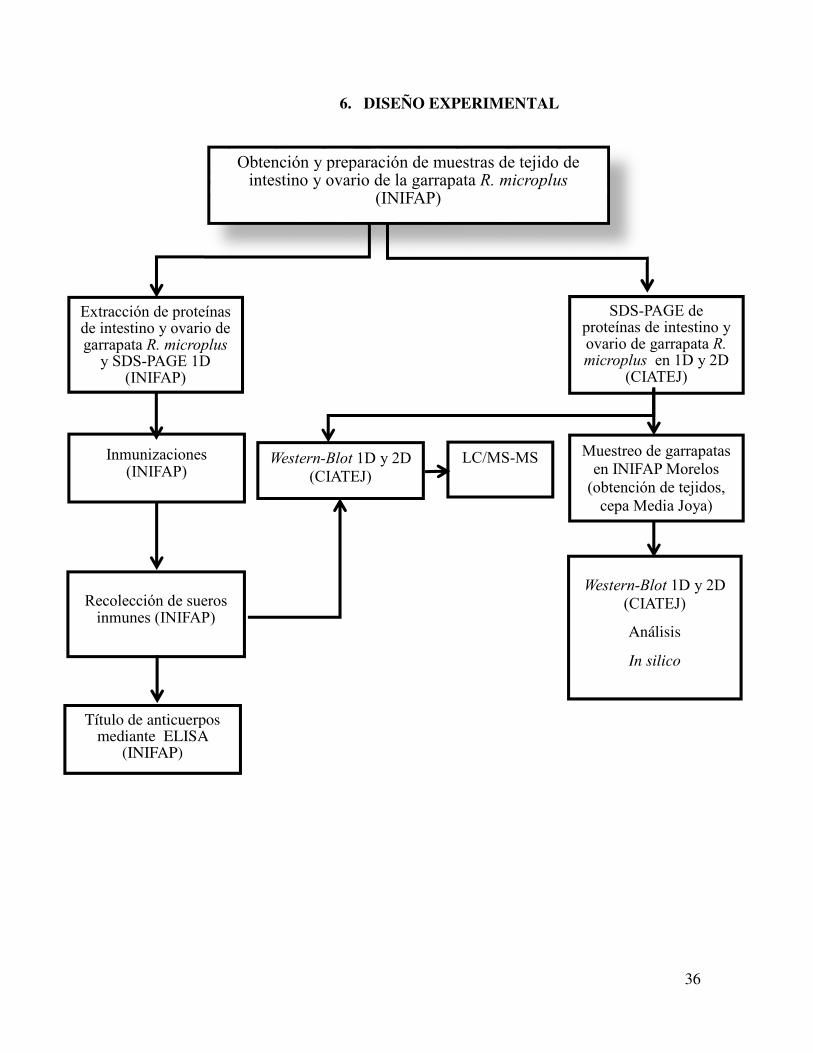
36
6. DISEÑO EXPERIMENTAL
Extracción de proteínas de intestino y ovario de garrapata R. microplus
y SDS-PAGE 1D (INIFAP)
Recolección de sueros inmunes (INIFAP)
Título de anticuerpos mediante ELISA
(INIFAP)
Western-Blot 1D y 2D
(CIATEJ)
Análisis
In silico
LC/MS-MS
Inmunizaciones (INIFAP)
Muestreo de garrapatas
en INIFAP Morelos
(obtención de tejidos,
cepa Media Joya)
SDS-PAGE de proteínas de intestino y ovario de garrapata R. microplus en 1D y 2D
(CIATEJ)
Western-Blot 1D y 2D
(CIATEJ)
Obtención y preparación de muestras de tejido de intestino y ovario de la garrapata R. microplus
(INIFAP)

37
7. MATERIAL Y MÉTODOS
7.1. Garrapatas
Las garrapatas adultas de R. microplus fueron obtenidas de una cepa susceptible, mantenida
en condiciones de laboratorio en el CENID-PAVET-INIFAP, ubicado en Jiutepec, Morelos,
México (Figura 8).
A B C
Figura 8. A) Imagen de un bovino en donde se mantiene la cepa de la garrapata de R. microplus. B) Lavado de garrapatas R. microplus. C) Acomodo de garrapatas R. microplus en cajas Petri con buffer PBS para su disección.
7.2. Disección de tejidos y extracción de proteínas
Se preparó un lisado de proteínas totales a partir de tejido de intestino y ovario diseccionado
de 15-20 garrapatas hembras adultas ingurgitadas, recolectadas al día siguiente del
desprendimiento espontáneo de los bovinos (aproximadamente 21 días después de la
infestación con larvas) (Figura 9), con base en el protocolo descrito por Rachinsky y col.,
(2008) con modificaciones. Se pesaron 100 mg de tejido de intestino y de ovario, se colocaron
en un tubo Eppendorf y se añadió 1 ml de PBS. Se rompió el tejido con un mazo cónico de
Teflon® y luego se pasó la suspensión a través de una jeringa (tamaño de aguja de 1 ml, 29
Gauge x ½ ") 10 veces, para ayudar a la fragmentación del tejido, hasta que se desintegró
completamente. La suspensión se incubó a 70 °C durante 5 min e inmediatamente después,
a 26 °C durante 5 min, repitiendo el procedimiento tres veces. A continuación, el lisado se
sonicó durante 5 min y se centrifugó a 13,000 rpm durante 15 min a 4ºC; después se separó

38
el sobrenadante. Las muestras de proteínas se cuantificaron mediante el ensayo de proteínas
DC (Bio-Rad, EUA) (Lowry y col., 1951). La absorbancia se leyó a 750 nm mediante un
Espectrofotómetro de Microplaca (XMarkTM, Bio-Rad, USA) (Figura 10).
A B C
Figura 9. A) Larvas de garrapata R. microplus. B) Infestación de bovinos con larvas de garrapata. C) Se muestra en la imagen cómo es sellada la abertura de la manga de los bovinos.
A B C
Figura 10. A) Disección de garrapatas. B) Maceración de tejidos de garrapata. C) Sonicación de tejidos de garrapata para obtener proteínas.
7.3. Electroforesis unidimensional
El análisis de las proteínas de intestino y ovario se llevó a cabo mediante geles de
electroforesis unidimensionales (1D) para verificar los patrones de expresión. Brevemente,
se hicieron geles de poliacrilamida de 7 cm de ancho x 8 cm de largo y 1 mm de espesor
(12%). Se llevó a cabo la electroforesis en gel de poliacrilamida-dodecil sulfato de sodio
(SDS-PAGE con el Sistema de Electroforesis Tetra Mini-PROTEAN® (Bio-Rad). Se
agregaron 10 µg de proteínas de intestino y 10 µg de proteínas de ovario en tampón de
muestra de SDS-PAGE y se calentaron a 100 °C durante 5 min. Los marcadores de peso

39
molecular (Precision Plus Protein Standards, Bio-Rad) se diluyeron 1:19 en tampón de
muestra SDS-PAGE y se calentaron a 100 °C durante 5 min. Los geles se corrieron a 120 V
durante 80 minutos en tampón de electroforesis (glicina 192 mM, Tris 25 mM, SDS al 0,1%),
se tiñeron con azul de Coomassie y se visualizaron y fotografiaron con un Sistema de
Documentación Gel-Doc XR (Bio-Rad).
7.4. Bovinos
Se utilizaron 10 animales de seis meses de edad, mestizos europeos con un peso promedio
de 140 kg. Se mantuvieron en las instalaciones experimentales de animales en el CENID-
PAVET-INIFAP. Los animales se alojaron individualmente en corrales de aislamiento y se
alimentaron con forraje y agua ad libitum. Los animales fueron asignados aleatoriamente a
cinco grupos experimentales. El primer grupo (dos animales) se inmunizó con proteínas de
intestino solubles, el segundo grupo (dos animales) con proteínas de intestino insolubles, el
tercer grupo (dos animales) con proteínas de ovario solubles, el cuarto grupo (dos animales)
con proteínas de ovario insolubles, mientras que el quinto grupo (dos animales) sólo con PBS
más adyuvante como control (Tabla 5).
Tabla 5. Grupos experimentales para el protocolo de inmunización.
Primera y segunda dosis
En 2ml de PBS 100 µg proteína/adyuvante 1:1 (1 ml proteína +��������� )
Proteínas de
intestino solubles
No 50, No 51
Proteínas de
intestino insolubles
No 44, No 47
Proteínas de ovario
solubles
No 46, No 42
Proteínas de ovario
insolubles
No 52, No 43
Testigo
No 48, No 45

40
7.5. Inmunización
El protocolo experimental fue aprobado por el Comité Institucional de Experimentación
Animal del CENID-PAVET-INIFAP (Anexo 1). El ganado se inyectó subcutáneamente en
el cuello con 2 ml/dosis (100 µg de proteínas de intestino o de ovario, solubles e insolubles)
con una jeringa de 5 ml y una aguja 18G. La emulsión se hizo con el lisado de proteínas de
intestino y ovario de las fracciones solubles e insolubles, en PBS más adyuvante Montanide
ISA 50V (anhidromannitoleteroctodecenoato en aceite mineral) (Seppic, París, Francia) a
razón de volumen 1:1. Se aplicaron dos inmunizaciones; la primera inmunización en el
primer día y la inmunización de refuerzo después de 15 días (Figuras 11, 12 y 13). Todos los
animales fueron revisados regularmente para detectar cualquier signo de reacción local o
anormalidades clínicas después de la inmunización.
Figura 11. A) Garrapata diseccionada, en la cual se observan los tejidos de intestino. B) Tejido de ovario de garrapata. C) Fotografía representativa de proteínas extraídas de garrapata. D) Emulsión de proteínas con adyuvante para inmunizar bovinos.
Figura 12. Esquema de inmunización e infestación de bovinos.
Figura 13. Esquema de sangrado de bovinos.

41
7.6. Prueba de ELISA
Se preparó un lisado de proteínas totales de larvas, de acuerdo con el método de Shahein y
col., (2008). Se homogenizó un gramo de larvas de 6 días en solución salina tamponada con
fosfato, enfriada con hielo (PBS); la solución resultante se filtró, se sonicó y se centrifugó a
13,000 rpm durante 20 min a 4 °C. Se separó el sobrenadante y se cuantificó la concentración
de proteína mediante el ensayo de proteína DC (Bio-Rad), como se mencionó anteriormente.
Se diluyó 1 µg de proteínas totales en 100 µl de tampón de carbonato de sodio (Na2CO3 50
mM) y se usó para recubrir cada pozo de una microplaca de 96 pozos; posteriormente se
incubó durante toda la noche a 4ºC. La placa se lavó cuatro veces con 300 µl de tampón de
lavado (PBS/Tween 20 al 0.5%) y después se bloqueó con tampón de bloqueo (PBS/Tween
20 al 0.5% y leche baja en grasa al 5%) durante una hora a temperatura ambiente (25ºC). A
continuación, la placa de incubación se lavó con tampón de lavado, como se mencionó
anteriormente. Los sueros de los bovinos inmunizados y no inmunizados fueron añadidos por
triplicado a los pozos, en dilución de 1:100. La placa se lavó como se ha descrito
anteriormente. Se utilizó un anticuerpo secundario (anticuerpo anti IgG acoplado a fosfatasa)
con una relación de dilución 1:2000. La placa se lavó una vez más y después se añadieron
200 µl de sustrato paranitrofenil phosfato (PNN) (Sigma 104). La placa se incubó durante 30
min y se leyó a 405 nm en un espectrofotómetro de microplaca (XMark™, Bio-Rad, USA)
(Figura 14).

42
Figura 14. Esquema general de la prueba de ELISA con los sueros preinmunes y de los bovinos inmunizados.
7.7. Electroforesis bidimensional
Para la electroforesis en 2D, se rehidrataron en gel durante una noche geles de tiras secas de
inmovilina con pH lineal 3-10 de 7 cm (Bio-Rad) con 12 µg de proteínas de intestino y ovario
(solubles e insolubles) en 125 µl de tampón de rehidratación (urea 7 M, tiourea 2 M, CHAPS),
DTT 1% y Tampón IPG 20 mM (3-10). La primera separación dimensional se realizó con un
sistema de Isoelectroenfoque (IEF) Ettan IPGphor ™ 3 (GE Healthcare); El programa se
ejecutó con cuatro pasos: Paso 1: (Paso) 300 V/2:30 h; Etapa 2 (gradiente) 1000 V/0:30 h;
Etapa 3 (Gradiente) 5000 V/1:30 h, y Etapa 4: (Etapa) 5000 V/0:35 h. Después, se
inmovilizaron las tiras de gel que contenían las proteínas de intestino y ovario;
posteriormente se equilibraron las tiras en dos etapas de incubación: con DTT al 1% (p/v) y
con yodoacetamida al 2.5% (p/v) durante 20 min, respectivamente. Una vez inmovilizadas,
las tiras de gel se transfirieron sobre un gel de poliacrilamida al 12% y la separación 2D se
llevó a cabo bajo las mismas condiciones de funcionamiento, como se mencionó
anteriormente para la electroforesis unidimensional. Las proteínas separadas se visualizaron
mediante tinción con azul de Coomassie. Se capturaron imágenes de electroforesis
bidimensional con el Sistema de Documentación Gel-Doc XR (Bio-Rad) y se analizaron por

43
Sigma Gel Software para calcular el peso molecular de las proteínas. La electroforesis 2D se
realizó en quintuplicado por muestra.
7.8. Ensayos de Western blot
Se realizaron inmunotransferencias en 1D y 2D, de acuerdo con el protocolo desarrollado por
Towbin y col., (1979) y Hawkes y col., (1982) con algunas modificaciones. En resumen, las
proteínas de intestino y ovario (solubles e insolubles) se separaron en geles SDS-PAGE al
12% y se transfirieron a membranas de nitrocelulosa mediante un aparato Mini Trans-Blot
(Bio-Rad) en tampón de transferencia (Tris 25 mM, glicina 192 mM, SDS al 0.1% Metanol)
a 110 V durante 60 min. Después de la transferencia, la membrana se bloqueó en solución
salina tamponada con Tris (TBS) que contenía Tween 20 al 0.1% (v/v) (TBS-T) y leche con
bajo contenido de grasa al 5%, durante 4 h a temperatura ambiente. La membrana se lavó
brevemente en TBS-T y después se incubó con los sueros de los animales control, preinmunes
o inmunizados en una dilución 1:50 en TBS-T durante toda la noche a 4°C. Después de la
incubación, la membrana se lavó 5 x 5 min con TBS-T y se incubó durante 4 h a temperatura
ambiente con una dilución 1:2000 de IgG de cabra anti-bovino acoplado a peroxidasa de
rábano (Jackson Immuno Research Laboratories, Inc., PA, USA). La membrana se lavó
entonces tres veces con TBS-T. La detección se llevó a cabo con el reactivo 4-cloro-1-naftol
(4CN, Bio-Rad).
7.9. Análisis de LC-MS/MS
Las proteínas reconocidas por el suero bovino en la membrana de nitrocelulosa, se obtuvieron
a partir del gel réplica correspondiente y se analizaron mediante LC-MS/MS. Las muestras
de gel se transfirieron a tubos previamente digeridos y se añadieron 100 µl de DTT 0.01
M/Tris 0.1 M, pH 8.5. El tubo se colocó en un bloque de calentamiento a 55ºC durante 1-2
h. Después de enfriar el tubo a temperatura ambiente, se retiró el líquido y se reemplazó con
100 µl de yodoacetamida 0.03 M/Tris 0.1 M, pH 8.5. Se dejó reaccionar durante 30 minutos

44
en la oscuridad, después de lo cual se retiró el líquido y se lavó el gel como se describe a
continuación.
Se prepararon piezas de gel para la digestión, lavando dos veces con 200 µl de Tris 0.05 M,
pH 8.5/acetonitrilo al 30% durante 20 min con agitación y una vez con 100 µl de acetonitrilo
durante varios minutos hasta que el gel era blanco opaco. Después de retirar el acetonitrilo,
las piezas de gel se secaron durante 20-30 min en un concentrador Speed-Vac.
Las piezas de gel se digirieron con 0.10 µg de tripsina modificada (grado de secuenciación,
Roche Molecular Biochemicals, Indianapolis, IN) en 50 µl de Tris 0.025 M, pH 8.5, o
volumen suficiente para hidratar completamente el gel. Los tubos se colocaron en un bloque
de calentamiento a 32ºC y se dejaron durante la noche.
Los péptidos se extrajeron con 2 x 50 µl al 50% de acetonitrilo/2% TFA y los extractos
combinados se redujeron en volumen en un concentrador Speed-Vac y se llevaron hasta 20
µl con ácido fórmico al 0.1%. Se analizaron alícuotas de 5 µl mediante LC-MS/MS.
El análisis de LC-MS/MS se realizó en un espectrómetro de masas de cuadruplo/tiempo de
vuelo, híbrido Waters Q-Tof Ultima con una fuente de nanoelectrospray. El eluyente se
introdujo en la fuente a través de una punta PicoTip New Objective de 15 µm de diámetro
interno. El voltaje capilar se fijó en 1.8 kV y la tensión del cono 32 V. La energía de colisión
se fijó según la masa y la carga del ion, de 14eV a 50eV. La cromatografía se realizó en un
HPLC LC Packings con una columna PepMap C18 mediante un gradiente lineal de
acetonitrilo con una velocidad de flujo de 200 nl/min.
Los archivos de los datos crudos se procesaron con el software MassLynx ProteinLynx v.4.1
y los archivos .pkl fueron enviados para búsqueda en www.matrixscience.com mediante el
algoritmo Mascot. La secuenciación de novo se realizó mediante inspección manual, ya sea
de los espectros crudos de MS/MS o los espectros procesados con el algoritmo MaxEnt 3
contenido en el paquete de software Waters MassLynx 4.1.

45
7.10. Aislamiento de RNA y síntesis de cDNA
Se diseccionaron 100 mg de tejido ovárico, se pesaron y se pusieron en un tubo Eppendorf,
con 1 ml del reactivo TRIZOL (Invitrogen, Carlsbad, CA, EUA) para extraer el RNA total.
El cDNA se sintetizó a partir de 5 µg de RNA, con el sistema de síntesis de primera cadena
Superscript® III (Invitrogen, Carlsbad, CA, EUA) de acuerdo con las instrucciones del
fabricante.
7.11. Síntesis de cDNA
Reactivo Condiciones de corrida en termociclador
• 1µl de oligo dT
• 1µl de buffer de alineación
• 8µl RNA
• 10µl (2X First-Strand Buffer)
• 2µl enzima mix
• 95 °C 5 min calentar tapa
• 50 °C 50 min oligo dT
• 50 °C 50 min random hexamer primer
• 85 °C 5 min
Almacenar a -20°C
7.12. Análisis bioinformático
Todos los cebadores fueron diseñados con el software público Primer3 v.4.0.0
(http://primer3.wi.mit.edu/; (Rozen y Skaletsky., 2000). El análisis filogenético se realizó
con el software CLC Main Workbench 6.0 (CLC bio, Cambridge, MA, USA), con 1,000
réplicas boostrap.

46
RESULTADOS
7.13. Cuantificación de las proteínas
En este trabajo se identificaron proteínas de intestinos y ovarios como una fuente potencial
de nuevos candidatos vacunales para combatir las garrapatas del ganado. Para esto, a partir
de las garrapatas disectadas, se realizó la cuantificación de proteínas totales obtenidas de
intestino y ovario, para ajustar la concentración a 100 µg/ml para la elaboración de la vacuna
con adyuvante. El rango lineal de este ensayo va de 50-500 µg/ml.
Los ensayos se realizaron por duplicado. En la Tabla 6 se presentan las muestras de estudio,
su concentración y el volumen que se utilizó de cada una. Para determinar la concentración
de proteínas de cada muestra se realizó una curva con el estándar albúmina de suero bovino
(BSA) (Figura 15).
Tabla 6. Muestras de proteínas y su concentración.
Muestras Concentración µg/µl Volumen µg/µl
Intestino soluble 1620.13 32.4026
Intestino insoluble 170.45 3.4091
Ovario soluble 420.45 8.4100
Ovario insoluble 68.17 1.3400
A B
Figura 15. Determinación de los valores de absorbancia de la proteína BSA en diferentes concentraciones.

47
7.14. Títulos de anticuerpos en bovinos vacunados
Las proteínas del intestino y del ovario (Figuras 16 y 17) se complementaron con el
adyuvante Montanide ISA 50V y se utilizaron para inmunizar el ganado vacuno. Las
proteínas de intestino y ovario generaron una fuerte respuesta inmune humoral en bovinos
vacunados. Los títulos de anticuerpos aumentaron después de cada inmunización, alcanzaron
su máximo en la cuarta semana y permanecieron altos hasta el final del experimento (Figura
18). Las muestras de animales control no mostraron cambios significativos en los niveles de
IgGs. Debido a que los sueros con los niveles más altos de IgGs se encontraron en la cuarta
semana, se eligieron para los ensayos de Western blot en 1D y 2D.
A B C D
Figura 16. Patrón electroforético de proteínas de intestino e inmunodetección en 1D. (A) Las proteínas solubles e insolubles de intestinos se sometieron a electroforesis unidimensional SDS-PAGE (12%) y se tiñeron con azul de Coomassie. Las réplicas de gel se transfirieron a membranas de nitrocelulosa y se incubaron separadamente con sueros control (B), preinmunes (C) e inmunizados (D), y se desarrollaron ensayos de Western blot. (1) Marcador de peso molecular (MPM); (2) Proteínas de intestino solubles; (3) Proteínas de intestino insolubles. Se muestra una inmunotransferencia 1D representativa. Para la inmunotransferencia se muestran (1) Proteínas de intestino solubles; (2) Proteínas de intestino insolubles; (3) MPM. Las flechas indican las bandas reactivas.
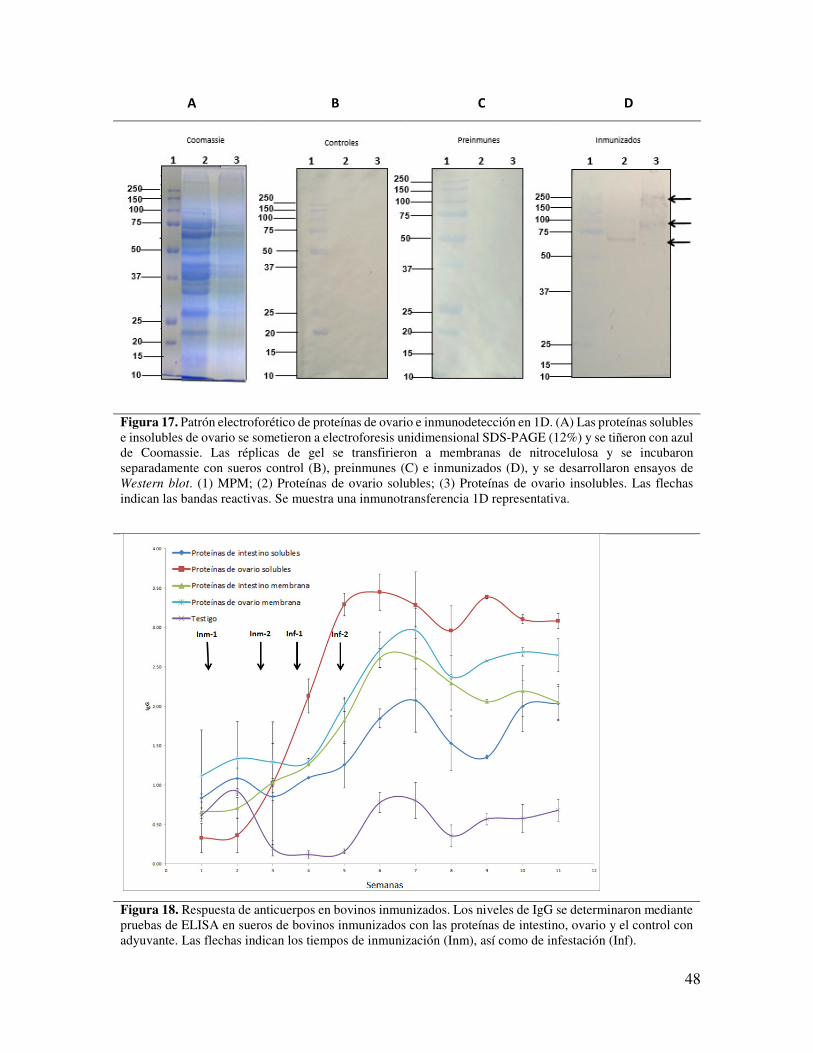
48
A B C D
Figura 17. Patrón electroforético de proteínas de ovario e inmunodetección en 1D. (A) Las proteínas solubles e insolubles de ovario se sometieron a electroforesis unidimensional SDS-PAGE (12%) y se tiñeron con azul de Coomassie. Las réplicas de gel se transfirieron a membranas de nitrocelulosa y se incubaron separadamente con sueros control (B), preinmunes (C) e inmunizados (D), y se desarrollaron ensayos de Western blot. (1) MPM; (2) Proteínas de ovario solubles; (3) Proteínas de ovario insolubles. Las flechas indican las bandas reactivas. Se muestra una inmunotransferencia 1D representativa.
Figura 18. Respuesta de anticuerpos en bovinos inmunizados. Los niveles de IgG se determinaron mediante pruebas de ELISA en sueros de bovinos inmunizados con las proteínas de intestino, ovario y el control con adyuvante. Las flechas indican los tiempos de inmunización (Inm), así como de infestación (Inf).

49
7.15. Detección de proteínas inmunogénicas mediante ensayos de Western blot
Los sueros recolectados en la cuarta semana de inmunización se utilizaron como primer
anticuerpo, contra las proteínas de intestino y ovario, en experimentos de 1D SDS-PAGE e
inmunotransferencias. Algunas proteínas fueron reconocidas con sueros de bovinos
inmunizados, pero no con sueros control ni sueros preinmunes (Figura 16 B, C y D; Figura
17 B, C y D). Con el fin de obtener una mayor resolución sobre el reconocimiento de
proteínas inmunogénicas, se realizaron ensayos de Western blot en 2D. Se encontró que los
sueros de bovinos inmunizados detectaron varias proteínas de intestino soluble, intestino
insoluble, ovario soluble y ovario insoluble (Figura 19 A y B; Figura 20 A y B; Figura 21 A
y B y Figura 22 A y B). Los puntos reactivos observados en las membranas de nitrocelulosa,
fueron identificados en los geles réplicas correspondientes. Las manchas de proteínas fueron
cortadas manualmente con los extremos de puntas de micropipeta nuevas y estériles, y
depositadas en tubos Eppendorf estériles. Las muestras fueron digeridas con tripsina y
analizadas por LC-MS/MS. Las muestras fueron nombradas en el orden en el que fueron
cortadas del gel (Tabla 7).
Figura 19. Detección de proteínas inmunogénicas de R. microplus en 2D. (A) Las proteínas solubles de intestino se sometieron a electroforesis bidimensional SDS-PAGE (12%, pH 3 a 10) y se tiñeron con azul de Coomassie. Las flechas indican las muestras cortadas del gel. (B) La réplica de gel se transfirió a una membrana de nitrocelulosa y ésta se incubó con sueros de bovinos inmunizados con proteínas de intestino, que se usaron como el primer anticuerpo para el ensayo de Western blot. Las flechas indican las bandas que fueron reconocidas por los anticuerpos. Se muestra una inmunotransferencia 2D representativa.
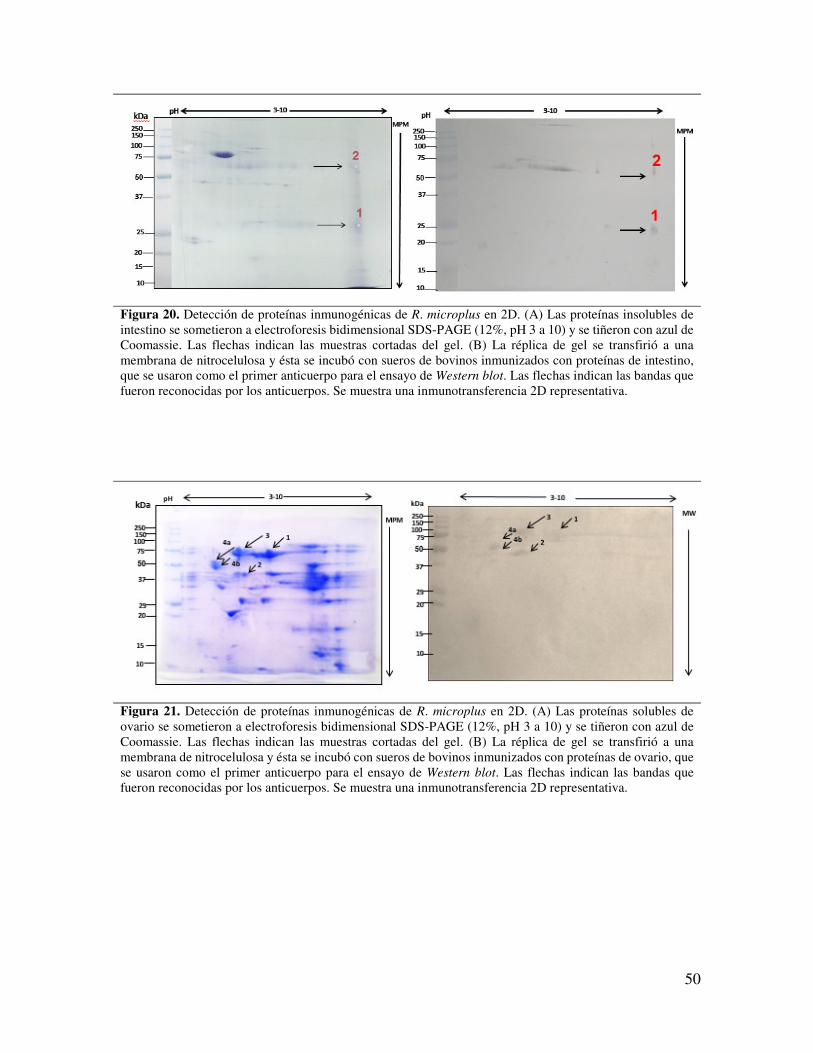
50
Figura 20. Detección de proteínas inmunogénicas de R. microplus en 2D. (A) Las proteínas insolubles de intestino se sometieron a electroforesis bidimensional SDS-PAGE (12%, pH 3 a 10) y se tiñeron con azul de Coomassie. Las flechas indican las muestras cortadas del gel. (B) La réplica de gel se transfirió a una membrana de nitrocelulosa y ésta se incubó con sueros de bovinos inmunizados con proteínas de intestino, que se usaron como el primer anticuerpo para el ensayo de Western blot. Las flechas indican las bandas que fueron reconocidas por los anticuerpos. Se muestra una inmunotransferencia 2D representativa.
Figura 21. Detección de proteínas inmunogénicas de R. microplus en 2D. (A) Las proteínas solubles de ovario se sometieron a electroforesis bidimensional SDS-PAGE (12%, pH 3 a 10) y se tiñeron con azul de Coomassie. Las flechas indican las muestras cortadas del gel. (B) La réplica de gel se transfirió a una membrana de nitrocelulosa y ésta se incubó con sueros de bovinos inmunizados con proteínas de ovario, que se usaron como el primer anticuerpo para el ensayo de Western blot. Las flechas indican las bandas que fueron reconocidas por los anticuerpos. Se muestra una inmunotransferencia 2D representativa.

51
Figura 22. Detección de proteínas inmunogénicas de R. microplus en 2D. (A) Las proteínas insolubles de ovario se sometieron a electroforesis bidimensional SDS-PAGE (12%, pH 3 a 10) y se tiñeron con azul de Coomassie. Las flechas indican las muestras cortadas del gel. (B) La réplica de gel se transfirió a una membrana de nitrocelulosa y ésta se incubó con sueros de bovinos inmunizados con proteínas de ovario, que se usaron como el primer anticuerpo para el ensayo de Western blot. Las flechas indican las bandas que fueron reconocidas por los anticuerpos. Se muestra una inmunotransferencia 2D representativa.
Tabla 7. Proteínas identificadas mediante análisis LC-MS/MS. Muestra Nombre gi Proteína
identificada
Masa pI Secuencia
1 1-Int-Sol 91982959 Cadena pesada de la IgG 1 [Bos
taurus]
52 6.5
1 1-Int-Insoluble L7M0M0 Componente TF-II D del factor de iniciación de la transcripción putativo
30 9.5 AVDVDDVR
2 2-Int-Insoluble L7MAA0 Subunidad alfa de la ATP sintasa tipo F0F1 putativa
59.9 8.9 VLSIGDG
1 1-Ova-Soluble 82878755 vitelogenina 85 6.0 FVLSAGK
2 2-Ova-Soluble 82878755 vitelogenina 65 4.8 FVLSAGK
3 3-Ova-Soluble 523713961 Precursor de la vitelogenina -2
75 5.5 YTGTLLGLCGD
4a 4a-Ova-Soluble 62199768 Catepsina de yema
60 4.8 VYTIFK
4b 4b-Ova-Soluble 62199768 Catepsina de yema
50 4.8 VYTIFK
1 1-Ova-Insoluble E7CF11 Canal aniónico selectivo mitocondrial dependiente de voltaje
29.5 8.9 NMNN
2 2-Ova-Insoluble Q7Z021 Serpina 43 5.5 NMWNMM
3 3-Ova-Insoluble Q6R128 β-actina 41 5.78 NCMCMNMMQM
4 4-Ova-Insoluble L7LW49 Proteína hipotética
34 5.2 SMMM

52
7.16. Proteínas identificadas por LC-MS/MS
7.16.1. 1-Int-Sol: cadena pesada de la IgG 1 [Bos taurus]
La proteína 1-Int-Sol fue identificada como la región constante de la cadena pesada de la
inmunoglobulina gamma 1 de Bos taurus, con un peso molecular de 52 kDa y un pI de 6.5
(gi: 91982959). Ésta fue obtenida de los intestinos macerados de garrapata, por lo que muy
probablemente haya sido ingerida a través de la sangre del bovino hacia el intestino de la
garrapata.
7.16.2. 1-Int-Insoluble: Componente TF-II D del factor de iniciación de la
transcripción putativo
El factor de transcripción TF-II D es uno de los diferentes factores de transcripción generales
que conforman el complejo de pre-iniciación de la RNA polimerasa II. La proteína TF-II D
(30 kDa, pI 9.5) en garrapatas, sólo ha sido identificada en la especie Rhipicephalus
pulchellus (Tan y col., 2012).
7.16.3. 2-Int-Insoluble: Subunidad alfa de la ATP sintasa tipo F0F1 putativa
La subunidad alfa de la ATP sintasa tipo F0F1 (F=factor de acoplamiento) es una enzima
transmembranal que cataliza la síntesis de ATP a partir de ADP, un grupo fosfato y la energía
suministrada por un flujo de protones (H+). Durante la respiración celular, la síntesis de ATP
se denomina fosforilación oxidativa y el flujo de protones tiene lugar entre el espacio
intermembranal y la matriz mitocondrial. La subunidad alfa de la ATP sintasa tipo F0F1
putativa, de 59.9 kDa y un pI de 9.5 en garrapatas, sólo ha sido identificada en la especie
Rhipicephalus pulchellus (Tan y col., 2012).

53
7.17. 1-Ova-Soluble, 2-Ova-Soluble: Vitelogenina
7.17.1. 3-Ova-Soluble: Precursora de la Vitelogenina-2
Las muestras 1-Ova-Soluble (masa 85 kDa, pI 6.0) y 2-Ova-Soluble (masa 65 kDa, pI 4.8)
fueron identificadas como vitelogeninas, mientras que la muestra 3-Ova-Soluble (masa 75
kDa, pI 5.5) fue identificada como el precursor de la vitelogenina-2. La vitelogenina es el
precursor de la vitelina, principal proteína de la yema de los huevos en animales, tanto
vertebrados como invertebrados, que comparten la misma descendencia (Chen y col., 1997).
En las garrapatas, la vitelogenina es sintetizada en el cuerpo graso y el intestino medio, es
secretada hacia la hemolinfa y se une a receptores de vitelogenina, localizados en la superficie
de los ovocitos, y se internaliza a los mismos. Dentro de los ovocitos, las subunidades de
vitelogenina son recombinadas y convertidas a vitelina, la proteína de almacenamiento del
huevo (Thompson y col., 2007).
7.18. 4a-Ova-Soluble, 4b-Ova-Soluble: Catepsina de yema
Las muestras 4a-Ova-Soluble (masa 60 kDa, pI 4.8) y 4b-Ova-Soluble (masa 50 kDa, pI 4.8)
fueron identificadas como Catepsina de yema. Esta proteína está implicada en la hidrólisis
de la vitelina en huevos de R. microplus (Logullo et al., 1998).
7.19. 1-Ova-Insoluble: Canal aniónico selectivo mitocondrial
dependiente de voltaje
La muestra 1-Ova-Insoluble (masa 29.5 kDa, pI 8.9) fue identificada como un canal aniónico
mitocondrial dependiente de voltaje. Los canales aniónicos dependientes de voltaje son una
familia de porinas eucarióticas dependientes de voltaje, que forman canales acuosos, y son
un componente central de la maquinaria apoptótica (Shimizu y col., 1999) ya que
desempeñan un papel esencial en la permeabilidad de la membrana celular.

54
7.20. 2-Ova-Insoluble: Serpina
La muestra 2-Ova-Insoluble (masa 43 kDa, pI 5.5) fue identificada como una serpina. Las
proteínas pertenecientes a la superfamilia de inhibidores de serina proteasa (serpina)
desempeñan un papel esencial en muchos organismos. En los artrópodos, estas proteínas
están implicadas en el sistema inmune innato, la morfogénesis y el desarrollo. Por lo tanto,
se plantea la hipótesis de que las garrapatas usen serpinas para evadir la defensa del
hospedero, lo que facilita el parasitismo (Tirloni y col., 2014). No está probada como
inmunógeno.
7.21. 3-Ova-Insoluble: β-actina
La muestra 3-Ova-Insoluble (masa 41 kDa, pI 5.78) fue identificada como la proteína β-
actina. La β-actina es una de las seis diferentes isoformas de actina que han sido identificadas.
Éstas están muy conservadas y están implicadas en la motilidad, estructura e integridad
celular. Debido a su conservación, el gen que codifica esta proteína ha sido utilizado como
un control y normalizador de la medición de DNA, cuantificado mediante PCR y RT-PCR.
Por otro lado, se ha identificado que algunos patógenos reclutan actina y proteínas del
citoesqueleto del hospedero hacia sus superficies y activan el ensamblado de una cola cometa
que les permite actuar sobre el hospedero (Goldberg, 2001, Gouin y col., 2005, Cossart y
Toledo-Arana, 2008).
7.22. Evidencia de múltiples vitelogeninas en R. microplus.
Recientemente Tirloni y col. (2014) reportaron las secuencias que codifican para la
vitelogenina-1, vitelogenina-2 y vitelogenina-3 de la cepa Porto Alegre de la garrapata R.
microplus. Estos genes fueron aparentemente numerados de manera errónea. Por lo tanto, en
el presente estudio se denominó a la vitelogenina-1 como vitelogenina-1, a la vitelogenina-3
como vitelogenina-2 y a la vitelogenina-2 como vitelogenina 3, con respecto a las secuencias
reportadas por Tirloni y col. (2014). En el presente trabajo se identificó a la vitelogenina-1 y

55
al precursor de la vitelogenina-2 como proteínas inmunogénicas, en el ganado vacunado con
proteínas de ovario. A partir de las secuencias reportadas para los genes de vitelogenina de
R. microplus, es posible deducir que la vitelogenina-1 es un polipéptido de 1783
aminoácidos, con una masa molecular de 201.58 kDa y un pI de 7.96. La proteína tiene un
péptido señal de 18 aminoácidos, dos sitios de corte de propéptidos, predichos en los
aminoácidos 464-467 y 663-666, que generarían diferentes subunidades con peso molecular
de 51 a 150 kDa, un dominio N-terminal de lipoproteína (LPD_N) el cual está entre los
aminoácidos 33-720, un dominio DUF1943 (aa 752-973) y un dominio C-terminal del factor
von Willebrand tipo D (vWD) de los aminoácidos 1464-1642 Figura 37). Los resultados
obtenidos en el presente trabajo sugieren que las muestras 1-Ova-Soluble y 2-Ova-Soluble,
podrían representar fragmentos diferencialmente procesados del extremo N-terminal de la
vitelogenina-1.
Con respecto a la vitelogenina-2 (vitelogenina-3 del estudio de Tirloni), es una proteína de
1923 aminoácidos, la cual tiene un peso molecular calculado de 217 kDa y un pI de 7.56.
Esta proteína tiene un péptido señal de 16 aminoácidos, cuatro sitios de corte de propéptido,
predichos en los amino ácidos 855-858, 884-887, 1156-1159 y 1562-1565, que podrían
generar diferentes subunidades con peso molecular de 40 a 176 kDa, un dominio LPD-N, el
cual se encuentra en los aminoácidos 28-735, un dominio DUF1943 (aa 771-1069) y un
dominio C-terminal vWD en los aminoácidos 1587-1755 (Figura 37). Dado su peso
molecular, la muestra 3-Ova-Soluble podría corresponder a un fragmento N-terminal de la
vitelogenina-2. Adicionalmente, una vitelogenina-3 putativa (vitelogenina-2 del estudio de
Tirloni) en R. microplus ha sido predicha. La proteína deducida sería de 1679 aminoácidos
sin péptido señal, un peso molecular de 189.89 kDa y un pI de 6.08. La molécula tiene
tentativamente ocho sitios de corte de propéptido que podrían generar subunidades de 20 a
165 kDa y un dominio C-terminal vWD en los aminoácidos 1100-1254 (Figura 23). La
secuencia de aminoácidos en las tres vitelogeninas identificadas es muy diferente y las
identidades entre estas proteínas son de menos del 25% (anexo 12). Se desarrolló un árbol
filogenético a partir de las secuencias completas de aminoácidos de vitelogeninas de
diferentes especies de garrapatas (Figura 24). En este análisis, la vitelogenina-1 de D.
variabilis, R. microplus y A. hebraeum formaron un mismo grupo. La vitelogenina-2

56
(vitelogenina-3 del estudio de Tirloni) formó otro grupo con la vitelogenina-2 de D. variabilis
y A. hebraeum, mientras que la vitelogenina-3 de R. microplus (vitelogenina-2 del estudio de
Tirloni) formó otro grupo en una rama separada. Adicionalmente, se confirmó la expresión
del mRNA de estas tres vitelogeninas en ovarios de garrapatas repletas de R. microplus
(Figura 25). Como se puede observar, la vitelogenina 1 y la vitelogenina 2 se expresan en
altos niveles, mientras que no se encontró expresión de la vitelogenina 3.
Rm-Vg-1 MRVLCGLPLLLLAAAANGVLFDP--VPPTQSGGVVYKVRATVTLKSPELSVTEGAGLQYA 58
Rm-Vg-2 MRVLAALGLLVAAVAALDVPYSVPKLVFEPRQEYLYKYRTTVALSVP-QKATHATGEEVY 59
Rm-Vg-3 ------------------------------------------------------------
Rm-Vg-1 GDLALYSAGQDTYLAQFLNFSVVKHEKLYGGLDNLTAVPLNESVPLFNESSAVDYSQKFR 118
Rm-Vg-2 GYLKVVVTAADPVAKTAR--LFVQLLNVTTSVFDKEVHDHLAPVPGLYHTPLEQYLQHFE 117
Rm-Vg-3 -------------------MINAFLATMQVKGNSRNNAEVQAELLWDADRYPDRKVRFSS 41
. .: . : . :
Rm-Vg-1 LPVKFVLSAGKVVKYEVSPEVPVWALNVYKSVLTLLQNQVETPQEVPVSAAY-------- 170
Rm-Vg-2 KHVVFTVEQLKVVTIEAPVDLPEPVVNLYRGLASVLTFSEPLTHAVPFSKVTPVSPDDEV 177
Rm-Vg-3 SHVKISTDGHPAFRYSATLQHANDMSITASGTYSVDLSQGPFAIEIGVGGTH-------- 93
* : . .. ... : . . :: . . : .. .
Rm-Vg-1 -----YEDGVSGHCRVTYEVHSLTKFLPTEGKVYNVTKTKYLDDCKLPRPVHSVSGAYRK 225
Rm-Vg-2 VVYKVFEEDLVGNCETTYHVLSDVHDLT----VLNLTKTKNYHKCLGLTSVFHYADYEYQ 233
Rm-Vg-3 ----------------------------------LTPRTVSLSYTRRETSANVMFGYTRA 119
.:* .. .
Rm-Vg-1 GYPAQCAKHLPNNFLPGFQEDTSAYESRPLAGCPEGLSPVDSVVTAHEVSYYNVSGS--- 282
Rm-Vg-2 GCPQTCQVHKPVTVFEGLEPEFTKYNDHYGAGCVKEYSQKNDVGDSFLTVRYNVSVQGEY 293
Rm-Vg-3 GADQLRIEYNRNEANSVQQFDVSATSVHPELNGKRLNIVRNLGGPRYFKIQYEHSANEIY 179
* : : : : . : . . : . *: * .
Rm-Vg-1 -LLEGALTDSLVVLPLVTSKVVVRTRLQLELASLS------VPPTNPVAFNGELRTSLEH 335
Rm-Vg-2 GLLETAKAVDKKVLVSGKQQLVATSVLALELLYKKSALRVDVQTTVPVKSYSNLTYAYPD 353
Rm-Vg-3 GLELTDSSGNNRINTNLKIRSPHLRYASNEITLTGDYSPARTNLIFEAYTQSEMEYRLTF 239
* : . : . : *: . . .::
Rm-Vg-1 QLPEVAEYLDLPVYSYLVGSPELVKPEVFTQVLEEVAEELVSLELDTETKKTPALMLKLV 395
Rm-Vg-2 QEYVVHGPALELEHLSLWGPIDRKEAMVAARSLVSRLAELVALDDLEVKDDYADLIVQLL 413
Rm-Vg-3 NYELTGNTAFATSLAMEVPKGSFRNGKVSLRLNRSPETTEGQLLVEHNDRVELDLTGNVR 299
: . . : * : . * * ::
Rm-Vg-1 HMVSLLNTDQLKQALP-QSLLTNTSELEPKQQVLKSLYVELLGKAGSKSAVEVAVHLVKQ 454
Rm-Vg-2 TVVNALPQEDLELLLKTVTPLEKVHVVSEKEYIERKLLLDALSLGGTDATAKTVLKLVKE 473
Rm-Vg-3 TQSANNFDGRIALRSPAVKDFEILVSSRSSSLLNGVYELKIIDQSNNPITATLTLSDRPR 359
: . : .. : :. :. ... :. .: .
Rm-Vg-1 QVLSLYDATRLFRDLSVYQAYVDKDTVELLLELCKTTETYLHPLVKVAVCDALGEAVQKA 514
Rm-Vg-2 EKLTLVEAVHVLSALQTSLVKPSTEVLDELLDICVGGKFVEKRLLFSSSCLALAKAVRKH 533
Rm-Vg-3 DFTGRLDLSGWLFERRPYLQVERKIPEAEHRVYSLKVGDTNSEVVLLADTAKRGDVRVTT 419
: : : . . :: : ... .
Rm-Vg-1 CPTKVHYEAVPYPTYKSSWRR-IPKLKSRYEQTPVVPKHLLPYESVERLVLPQEKQCTVA 573
Rm-Vg-2 CTHHDLSLPTTVQLGKKAMMRNVMKIRSKHTYSSEELDDDVTTEPKELPQYETTVSCSLQ 593
Rm-Vg-3 FKYQPRDKPQNWASVTIEKND----WNPQHEMLMVTVEPKANLKTTFDFLYLITKGSLKS 475
: . . ..:: . :. .

57
Rm-Vg-1 DLYEYVTALEEALKVTKDFKVLVGYLNALGKVAKPEVLPVLMSYLNGTAENLYELAEQEE 633
Rm-Vg-2 DYAKYVKAIADKLEQSKEFHEVTVLLHTLGKLEHPLALKALVPFVLGKHP-LCKLTLGSE 652
Rm-Vg-3 KFIWTLDDLKLGYDLKSKAEGDNAYNSLLKILYLRREIDVHNHYSHTPDSMELSTRILLN 535
. : : . ... . * : : . : . :
Rm-Vg-1 DVTETVYFVKKAVLLALSHVVEYFPKEVNPLVRAVLLNVSEPVDVRTLAFDVWLKSVPTK 693
Rm-Vg-2 DESEACQYLRLVTLHSLKHAAVTQAVLVQPVLQAVYFDKDEDYELRSVALSLLLLTQPTE 712
Rm-Vg-3 AIMMPDRALEMTYRHNRIQNGWETTAVIRHPTFSNDITFNGKIQKDISESTPLLVSAQLQ 595
. :. . : . :. : : . : * : :
Rm-Vg-1 WDLQTVSLVVKLDRSLELKSYVYSALKYVVKDKHPANHVLASRVRGVFVHLDALNVGPRY 753
Rm-Vg-2 PVLARVVLELPREKSLHVASYTYHTLLAFSNHTLPCYKATAYRLKKLLPALTHHKYGFQY 772
Rm-Vg-3 PGDASNKMYFNLVEKLNEITATNKSVHIHIHHEDRSLFDTSMTVYHTITPERPVFAGYYW 655
: . ..*. : . :: :. . : : : * :
Rm-Vg-1 SKYVKKTYYNVAKHFGVESVFKHVANNVSFFPTHLHAGLKYNLGPYVKTLLEGKLLLKGG 813
Rm-Vg-2 SSFHYVTDYSVESGVGLQGYFGVVKGNNTVLPRSLFYGLEFTKGPYTSPLLDVMVVSKGL 832
Rm-Vg-3 SWNTQKQGPKRGHATLFIGRNGAAELNYDSILGKWTATGQSQRAPNGDETVDVVFQGERE 715
* . . . . * : : .* . :: . :
Rm-Vg-1 EKFLAEVFSTYGPSTLRSVFSRNIESTYESGAKYS----------VVKELNIETR-KVEF 862
Rm-Vg-2 QNVHTWLVSEQGPASLLAKAFERVRRGLRASLDDTYQPLNVLLDMLDQHFDFDTKSVVEE 892
Rm-Vg-3 QFRGRIIYNLRRYRFEGLTFDEQGKTSKSLEISAS---------------NSTRNAALKV 760
: : . .. . . : : . ::
Rm-Vg-1 PKAVLFAKFLGGDVVLPVDKELFDTLKTELVEYLKKVSSSGPVTTHLVRVLLPKKYVHVE 922
Rm-Vg-2 PKLVLDTKFLGTDLFLPLDKYFVKELLTKGADWVKEK-WVHPVSARYVQLRVPFTYRKLL 951
Rm-Vg-3 EFSHFEDNNKMTDFSYSFEKRGDKAYRSKLFFPPEKMRTIQTAMNEVGEKISEIDISNFV 820
: : *. ..:* . :: :: .. . . :.
Rm-Vg-1 PTVVGLPVVQTVLRPVVVSLKLKDVSVLAETSVDSLLPVTLNVTGTVQPLVLSVKQNRVF 982
Rm-Vg-2 PTVTGYPVALTTRYPTVVSLSLKDLKLRYQPNVKSLVPSVFAVSALVQPTVYTTAVTSAL 1011
Rm-Vg-3 SDVSDMAVVSGKYLDKNFFSPLTNLLLDEFGEIMGEAVIALSDVLYESPFGELIAHFQRW 880
. * . .*. . *.:: : .: . .: .*
Rm-Vg-1 VPADLTEESPSVLHTSVKHLQGKAHLTLGYDRKYEKKVRVAVKPTFGTTLLSAISTEHVV 1042
Rm-Vg-2 TVTPFTKLVAAVRVIEKTRFTHPHDLALQFS-SVNHTLALTFRPRFTKLFTHETKTMTYT 1070
Rm-Vg-3 VESLFKMAGEEIKMFIEELSRTVQSTAGFFSGVRGRRRRDVSQRWTELSDYVSRMVDDVR 940
. : :. : : :. : . : .
Rm-Vg-1 ETPNPFLLLEKPVVDYKKVLDSLVKPFYYHKVYEAKLVNVEVESVSHGPLLPLPLYGKLY 1102
Rm-Vg-2 TPAFFFTAPQRSVLATMSVLATQSKPFEYRKVLGAKYLGVGVEVTG-LTSEPRKTPLLLL 1129
Rm-Vg-3 NAPIVRQVTRKVEDAADYVTDKISEMLYRARRAVWDAVEAIKENPDYRSLESLIGELYEP 1000
.. .: * . : : : . : . * . . .
Rm-Vg-1 QQPSERMTKFVEYLTHTATKQHLLKVKVLPYEKDPVTEWGTTFKYEHNLD---------F 1153
Rm-Vg-2 AKRKALVKSFLDVVLNPWAVSRKWEARLVRSEKNPVDEYKVVLRLTTTMNGTDPADYVEE 1189
Rm-Vg-3 GTYGANTEFWQNLQRNLRESVTARDVAVLREADRKNGKYIVDFNLPTDLDS--------- 1051
: : : .. :: . :: . :. ::
Rm-Vg-1 LAKTTLKKVYTPTKPLAYDESQETPYERMHYKPAT-------PGYETYESEESYDYYPEY 1206
Rm-Vg-2 AKPLYRTRAHYPTVRQTYPEDLKTPVYEELFEKATGFLPPVLEHHEVLNKVVEASKLTSY 1249
Rm-Vg-3 ------IRQSWHEWLAEEDDDAAERRPKKSAESTP----------DMIDYISSMFQRKWM 1095
: :. . : :. : : .
Rm-Vg-1 NQWN--KTLVKHVLNVTLEG-------KHEGSLKKLTKLDFAYYHTLNKTLKHYYVDVKT 1257
Rm-Vg-2 PKADNVKPLLQNVTNTTYAYGVEFVVTALGPEEPKAVYGHFVYGTSFDKTLKLYQLYVNK 1309
Rm-Vg-3 PPFDGQAMIIGHQHFVTFDG------------TMYSATGDCTYLLARDFVDGNFTVLLKY 1143
: :: : .* . . .* : : . : : ::
Rm-Vg-1 LTRPLVSVFLNLTHPVP-PSPFWYQPSFVGKDLVNATLYVTVGN---EPEPFVLTYNATK 1313
Rm-Vg-2 VDSEYETVVNSALLKSAKPSPFTTYATRKDSHYLYTTSTAEVHTPHYGPLKYSLELEATP 1369

58
Rm-Vg-3 HPENPRIHGRVPKSIIVQLGQSYIEIFPDDGSMFLNGQAVELPLILEGGEVIARRVDDVI 1203
. . . . : : .
Rm-Vg-1 TVQQLKGLHAYDAT---------------------------------------------- 1327
Rm-Vg-2 SVEQLEKTPLMIVDGAIKKIEPRRQIKSSLWRRSEYEDDSYYSKRRSSKHPKYEDEDLPY 1429
Rm-Vg-3 TVEDEKALR--------------------------------------------------- 1212
:*:: :
Rm-Vg-1 ----------PLLHWFVPQCLADQYAG-NTVSYACRLATVHDAHLNKQVVAFKVPTQVSP 1376
Rm-Vg-2 STYSAYSVPVTELPWYYRQCKEDVESGKELVSSACKKVHWHEHTLDKFVLKVTLPTVYPS 1489
Rm-Vg-3 ----------VSCHLYYDVCTVKINGWYFGNTAGLLGTYNNERGDDLMKPKGQVTSNVAQ 1262
: * . . : . . :: : :.: .
Rm-Vg-1 KVKSVALKFLSYLKFRMFPYATFYALPLTEQEEYYVVFKLNKTDVNPYVTVAHGELLLP- 1435
Rm-Vg-2 LALNVTHKLFQTAKVLLYDKSTTEYVESPVVTEGQFLVEAVLEDLYSGVTTANVTVHTPH 1549
Rm-Vg-3 FVKKWETSRGCKGPVSVLSEQVAVDSEGYNLCEKYFKDDDSPLADGFWQESPEPYFDLCL 1322
. . . . : . * . . .. .
Rm-Vg-1 GEKVVLTDLKLSKYLVPNLVLSGLDRLKHSVFEG-HPHPPCSVGKNWVRTYDNVSYPLEV 1494
Rm-Vg-2 FEKVTFNRLPWWRALRPTVAYDMTTQAVGLARKG-YPFPTCMVSPYYLKTFDNVTFPLET 1608
Rm-Vg-3 RHMATPGIEPRHAICNISMAYLLQLEKYSISANLPSECYTCSLPGGATLKFGDYLTLGSD 1382
. .. .:. . : .* : .:.: .
Rm-Vg-1 RPNCKYLVTSDCSAKHDFAVVAQPLDLTVGTKKLIVQLGPTVVELPPPDLYKAEVLLTVN 1554
Rm-Vg-2 LKKESTHVLLRHAVDEPEFYVLYEQRSEDATTLQVVLKNQTLVKLTPP-KDQTTYEVEVN 1667
Rm-Vg-3 NSAPRRPSSMDIVLVVEEDACHADVVRELDNTMRIVDKELLSAGFSNNRFALIGFGHGSG 1442
.. :* . :. .
Rm-Vg-1 GTYYVANTTQDVVLPYKWDRKLLVTVYPTSGPHDPPVVKLTNSLKTFKLLFDGVNFFVWV 1614
Rm-Vg-2 GTLLTVDPLKSHVLQYYYNFSSQVLLYVTAHPEVAPTLWLKVRDSGLVLAYNGSSVLVSV 1727
Rm-Vg-3 HNSMPHVRTARGSIFFESHNLPLATQKMRLDAPATSEGRPVNKDVFDAIRYASLLPFRAN 1502
. : : . . . .. : : . :
Rm-Vg-1 N-PLYQGKTCGLCSNYDNEPYHEFVTPENWLVSNYSEFVASYGFGLPQCKEPLVYTLP-- 1671
Rm-Vg-2 MLPKYTGTLLGLCGDNNRESQYEYVTPEQCVVTEVEDFLNAYSFEAEKVVKGEVYCPPGV 1787
Rm-Vg-3 VAKAIIVIACADCKEEQSELSYSDIQTQLLDHGITLHFVSDKRIEVRKSIIKGKGIYG-- 1560
. * : : * :. : .: .*: : :
Rm-Vg-1 ---------------VHYLEELKKPLGYESSYPTHREYPTHHKYPTHREYPTHREYPTRH 1716
Rm-Vg-2 TLKVGVPVYNKTKITKSKITKISKVRGYKASKVDSDLYDDVVSRIATPECLTERRKVIYE 1847
Rm-Vg-3 --------VDADSVYGSKDVSQKMLLGQPDLRPQVAVAKDICIALAQEVHGSFFSSAMLR 1612
. . * : : .
Rm-Vg-1 EYPTRHEYPTRHEYPTHHKYPTHHEYPTRHEYPTHHEYPTRHEYPTHHKYPPHHEPYPYT 1776
Rm-Vg-2 EGKVCISLVKTTACKRGCKPTTTEKKELEFVCLNKENPTLKKMLKDIQSKRELPLSLADI 1907
Rm-Vg-3 SDGKNWKTVFARRVAKALKPAKQYGGAEDYCERCECTHGSDVRPQVVCRPCRPLKPKVPL 1672
. . * .. . . .
Rm-Vg-1 ANLQREG--------- 1783 Identity
Rm-Vg-2 PTITKSVDVPVDCVPK 1923 22%
Rm-Vg-3 ALYTAED--------- 1679 8%
Figura 23. Alineación de la secuencia de aminoácidos deducida de Vitellogenin 1 (JAC58991.1), Vitellogenin 2 (Vg-3, JAC58954.1) y Vitellogenin 3 (Vg-2, JAC58956.1) de R. microplus. Se muestran los dominios característicos. El péptido señal está sombreado en amarillo. El dominio de la lipoproteína N-terminal está sombreado en gris. El dominio de la función desconocida (DUF1943) está sombreado en cyan. Von Willebrand El dominio del factor D está sombreado en verde. Los posibles sitios de escisión se muestran en negrita y subrayados.

59
Figura 24A y B. Vitelogeninas en R. microplus. A) Comparación esquemática de las tres Vitelogeninas de R.
microplus que muestra sus dominios característicos. S, péptido señal; LPD_N, N-terminal dominio lipoproteína;
DUF1943, dominio de función desconocida; VWD, von Willebrand Tipo de factor D. Las flechas indican las
posiciones aproximadas de sitios potenciales de escisión. B) Análisis filogenético de Vitelogeninas de diferentes
especies de garrapatas. El árbol evolutivo se obtuvo por el método UPGMA basado en las alineaciones de
aminoácidos de la R. microplus Vg-1 (JAC58991.1), Vg-2 (Vg-3, JAC58954.1) y Vg-3 (Vg-2, JAC58956.1),
Haemaphysalis longicornis Vg-1 (BAJ21514.1), Vg-2 BAG12081.1) y Vg - 3 (BAJ21515.1), Dermacentor
VG-1 (AAW78557.2) y Vg-2 (ABW82681.2), Ixodes scapularis Vg-a (XP_002403966.1) y Vg-b
(XP_002415224.1), Amblyomma hebraeum Vg-1 (AGQ57039.1) y Vg-2 (AGQ57040.1) y Ornithodoros
moubata Vg (BAH02666.2). Los valores de bootstrap de 1000 simulaciones se muestran en las ramas. El
análisis filogenético se realizó con el CLC Software Main Workbench 6.0 (CLC bio, Cambridge, MA, EE.UU.).

60
Figura 25. Expresión de vitelogeninas en los ovarios de R. microplus. Se realizaron reacciones de RT-PCR a
partir del RNA total de los ovarios de las garrapatas hembras adultas injurguitadas. Actina fue utilizado como
control. 1) Mpm de 100 bp; 2) Vitelogenina 1 (219 pb); 3) Viteilogenina 3; 4) Vitelogenina 2 (162 pb); 5) actina
(140 pb).
7.23. Productos de PCR´s en geles de agarosa al 1% de la especie R.
microplus en secuencias que codifican para las proteínas vitelogeninas
de ovario
Se obtuvieron las secuencias para las proteínas; vitelogenina 1(Vg-1), vitelogenina 2 (Vg-2)
y vitelogenina 3 (Vg-3) de R. microplus a partir de la base de datos de secuencias genéticas
GenBank (GBBR01000071.1, GBBR01000108.1 y GBBR01000106., respectivamente. Se
utilizaron los siguientes cebadores (Integrated DNA Technologies, Coralville, IA, USA): F-
RmVg1 5 'CCA ACT CGC TCA AGA CCT TC-3' y R-RmVg1 5'-AAA CCA GGG GTT
CCT TGC AC-3 '; F-RmVg2 (3) 5'-CTA CCT AAG GCG CGA GAT TG-3 'y R-RmVg2 (3)
5'-TGG ATT CCG AGA TGT CCT TC-3'; F-RmVg3 (2) 5'-AGC TTC GAC AAG ACC
CTC AA-3 'y R-RmVg3 (2) 5'-GGT GCT GGT GGT GTA GAG GT-3', a partir de cDNA
de ovarios como molde. Las condiciones de PCR fueron: 94ºC durante 5 min, seguido de 35
ciclos, cada uno consistía en una desnaturalización a 94ºC durante 30s, hibridación a 55ºC

61
durante 30s, y extensión a 72ºC durante 60s. Se incluyó un paso de extensión final a 72ºC
durante 7 min. Los productos amplificados se corrieron en geles de agarosa al 1% y se
analizaron en un transiluminador UV (BioRad Laboratories, Philadelphia, PA, EUA). La
amplificación de control se llevó a cabo con los siguientes cebadores: F-5'- CCC ATC TAC
GAA GGT TAC GCC-3 'y R-5'-CGC ACG ATT TCA CGC TCA G-3 ', diseñado a partir de
R. microplus actina (GenBank número de acceso AY255624.1). Las condiciones de PCR
fueron las mismas que las mencionadas anteriormente.

62
8. DISCUSIÓN
La vacunación se considera una alternativa ambientalmente segura y económicamente
sostenible para el control de los ectoparásitos. Los beneficios del uso de vacunas contra la
garrapata, como parte de un programa de control integral, incluyen una reducción en el uso
de acaricidas, lo cual prolonga la vida útil de los acaricidas y retrasa el inicio de la resistencia,
esto a la vez disminuye la incidencia de enfermedades transmitidas por R. microplus y reduce
los costos de producción (De La Fuente y col., 2007; Rodríguez y col., 1995a, b; De La
Fuente y col., 1998).
Se han realizado esfuerzos significativos en todo el mundo para identificar y caracterizar a
las proteínas de diferentes especies de garrapatas para el desarrollo de nuevas y mejores
vacunas anti-garrapata. En la actualidad, únicamente existe una vacuna comercialmente
disponible en el mundo, la cual está basada en el antígeno Bm86 y es comercializada bajo el
nombre de GavacTM. Esfuerzos previos se habían enfocado principalmente en antígenos del
intestino y de las glándulas salivares, y más recientemente de órganos internos como los
ovarios, que son esenciales para la reproducción de la garrapata. Por ejemplo, Rachinsky y
col., han reportado una expresión diferencial de proteínas en los ovarios de garrapatas
infectadas o no infectadas con Babesia. Entre las proteínas del ovario que estuvieron sobre-
reguladas en las garrapatas infectadas se incluyen; la calreticulina, dos subunidades de la
miosina, una proteína del retículo endoplásmico, una cis-trans isomerasa peptidil-prolil
(PPIasa), una subunidad de citocromo c oxidasa, una glutamina sintetasa y una familia de
inhibidores serina-proteasa del tipo Kunitz (Rachinsky y col., 2007). Estos descubrimientos
son apoyados adicionalmente por un estudio de transcriptoma del ovario (Heekin y col.,
2013). En el presente trabajo se siguieron los procedimientos clásicos de vacunología, a partir
de extractos de proteínas totales como inmunógenos, y mediante su detección por los
métodos de ELISA, Western blot en 1D y 2D y análisis de LC-MS/MS.
Para el desarrollo de la prueba de ELISA, se propuso un protocolo experimental, en el cual
se fijaron en la placa proteínas de larva, en lugar de proteínas de tejidos adultos, a partir de

63
la hipótesis de que el suero del ganado inmunizado puede reconocer a las proteínas presentes
tanto en larvas como en ovarios de garrapatas adultas. Si estos sueros, que contienen
anticuerpos generados contra proteínas del intestino y del ovario adulto, pueden detectar
proteínas de larvas, sería ventajoso, dado que los blancos moleculares estarían presentes y
podrían ser reconocidos desde las primeras etapas parasitarias de las garrapatas.
Se sabe que una vez finalizada la vitelogénesis, el huevo contiene todas las substancias
necesarias, así como el material genético necesario, para el desarrollo del embrión durante la
embriogénesis, la cual ocurre fuera del organismo materno (Logullo y col., 2002). En
particular, la vitelina (una combinación de vitelogeninas procesadas y la principal proteína
de la yema de los huevos) sirve como una reserva de nutrientes para el embrión, antes de la
eclosión e inclusive para los periodos de no alimentación de la larva, ninfa y garrapata adulta
(Rosell-Davis y Coons, 1989). Se han encontrado subunidades de vitelina en huevos y larvas
no alimentadas en garrapatas Dermacentor variabilis (Rosell-Davis y Coons, 1989). Más
aún, se han encontrado enzimas que degradan vitelina y hemoglobina, tales como BYC,
VTDCE, THAP y BmCl1, las cuales son proteasas de yema maternalmente producidas y
acumuladas en ovocitos, en huevos y larvas de la garrapata R. microplus (Bergamo y col.,
2010; Seixas y col., 2012). Aun cuando las garrapatas en sus etapas inmaduras no presentan
órganos reproductivos, los huevos, larvas e inclusive las ninfas, preservan mucho del material
maternalmente transmitido a través de los ovarios. El aumento de la concentración de
anticuerpos, observado en este estudio, sugiere que el suero contiene anticuerpos que
reconocen proteínas compartidas tanto por larvas como por ovarios adultos.
Con respecto al análisis proteómico, en este estudio se lograron identificar diez proteínas
inmunogénicas. Algunas de estas proteínas ya han sido evaluadas previamente como vacunas
experimentales. Por ejemplo, en 2002 se reportó que la proteína GP80, que en realidad
corresponde al fragmento C-terminal de la vitelogenina-1, tiene un papel protector en contra
de las infestaciones por garrapata en ovejas inmunizadas (Tellam y col., 2002). Asimismo,
la proteína catepsina de yema ha sido evaluada en su forma nativa y recombinante, que genera
una respuesta humoral en el ganado inmunizado y protegiéndolo contra las infestaciones por

64
garrapatas (Seixas y col., 2012). Por consiguiente, las proteínas identificadas tienen un gran
potencial como candidatos vacunales, lo cual amerita su evaluación.
8.1. Vitelogeninas en garrapatas
Las evidencias recientes sugieren que las garrapatas presentan múltiples vitelogeninas.
Thompson y col. (2007) reportaron el primer mRNA de vitelogenina de garrapatas. Ellos
mostraron que el mensajero de vitelogenina fue detectado en hembras Dermacentor
variabilis, después del apareamiento y alimentación con sangre hasta repleción. Esta
expresión fue regulada por ecdisteroides. En la etapa de pre-oviposición (apareadas y
repletas), se encontró una elevada expresión de vitelogenina en el cuerpo graso e intestino, y
baja expresión en el ovario de hembras. En D. variabilis, Khalil y col., (2011) reportaron un
segundo gen de vitelogenina. La inyección de hembras vírgenes con 20 hidroxiecdisona
indujo la expresión de la vitelogenina 2, la vitelogénesis y la oviposición. Mediante ensayos
de RT-PCR, la expresión de vitelogenina 2 se detectó solamente en garrapatas hembras,
después del apareamiento y alimentación hasta repleción. El análisis de Northern blot mostró
que la vitelogenina 2 se expresa en el cuerpo graso e intestino de hembras vitelogénicas, pero
no en el ovario. Mientras tanto, Boldbaatar y col., (2010) han reportado la presencia de
múltiples vitelogeninas en la garrapata Haemaphysalis longicornis. Los perfiles de expresión
de tres genes de vitelogeninas se determinaron por análisis de RT-PCR, a partir de muestras
de RNA total de diferentes tejidos de garrapatas hembras adultas, parcialmente alimentadas,
de la cepa partenogenética. La vitelogenina-1 se expresó en el intestino solamente. La
vitelogenina-2 se expresó en el ovario y el cuerpo graso, mientras que la vitelogenina-3 se
expresó solamente en el cuerpo graso. No se encontró expresión de estas vitelogeninas en las
glándulas salivales ni en los túbulos de Malpighi. La evidencia acumulada indica también la
existencia de varias vitelogeninas en garrapatas R. microplus, como sucede en las garrapatas
H. longicornis, D. variabilis e Ixodes scapularis (Boldabaatar y col., 2010) y coincide con un
reporte reciente de múltiples bandas inmunogénicas relacionadas a la vitelogenina,
encontradas en garrapatas R. annulatus (Taheri y col., 2014).
Con respecto a la expresión de las vitelogeninas en R. microplus, se encontró que las
vitelogeninas 1 y 2 se expresan en los ovarios, no así la vitelogenina 3. Esto podría sugerir

65
que en R. microplus, la vitelogenina 3 sería importada a los ovarios a partir de tejidos extra-
ováricos, tales como cuerpo graso o intestino. Sin embargo, las vitelogeninas 1 y 2 podrían
ser activamente sintetizadas en los ovarios, además de otros posibles sitios. Las garrapatas
muestran múltiples fuentes (tejidos) de mRNA de vitelogeninas, por ejemplo, el cuerpo
graso, intestino y ovario, pero estas moléculas son expresadas en forma específica de tejido
entre las garrapatas. La contribución relativa de cada gen de vitelogenina durante la
vitelogénesis, al presente, no se conoce. Más aún, los mecanismos proteolíticos que operan
para generar las diferentes subunidades, a partir de los diferentes tipos de vitelogeninas, y su
utilización como proteína de yema durante el desarrollo del ovocito, necesitan ser
dilucidados.
En años recientes, la aplicación de la tecnología proteómica para el estudio de las garrapatas
ha permitido la identificación de cientos de antígenos potencialmente protectores, pero hasta
donde se sabe, ninguno de estos candidatos se ha utilizado en ensayos de vacunación. Esta
limitación se debe probablemente a las dificultades asociadas con el tiempo y los costos
necesarios para los ensayos, para evaluar la eficacia de las vacunas bajo condiciones
controladas y de campo (Villar y col., 2017).

66
9. CONCLUSION
En el presente trabajo, se lograron identificar diez proteínas de R. microplus con un gran
potencial inmunogénico, bajo un enfoque inmunoproteómico en el Centro de Investigación
y Asistencia en Tecnología y Diseño del Estado de Jalisco (CIATEJ), A.C. en colaboración
con el Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP).
Asimismo, se demostró que el tejido obtenido de los ovarios de R. microplus induce una
mayor respuesta humoral, ya que actúa directamente como un sitio activo en la síntesis de la
proteína vitelogenina.
Estos resultados aportarán conocimiento suficiente para comprender e inferir mejor el
proceso de maduración de los ovarios y el ciclo reproductivo de R. microplus, ya que sentará
las bases para explorar su potencial como posibles candidatos al desarrollo de una vacuna
efectiva. Se sabe que la vitelina sirve como una reserva de nutrientes embrionarios antes de
la eclosión, e incluso durante los períodos de no alimentación de las larvas, las ninfas y los
adultos. El contenido de vitelina tiene una proporción inversa con respecto al desarrollo, sin
embargo, una vez que las garrapatas inician el período de subsistencia rápida, se re-activa la
maquinaria transcripcional de nuevas moléculas de vitelogenina en el parásito. Estos eventos
aseguran una bio-disponibilidad constante de blancos moleculares en diferentes etapas del
ciclo de vida de las garrapatas al ser presentadas con el ganado previamente inmunizado con
estos antígenos (vitelogenina). Por lo que, la generación de vacunas dirigidas contra blancos
específicos del ovario ofrecen la posibilidad de interrumpir el ciclo reproductivo de las
garrapatas y de esta forma controlar y eliminar las poblaciones de R. microplus. Sin embargo,
es necesario realizar más estudios de inmunizaciones con las proteínas identificadas, ya que
desempeñan un papel esencial en la reproducción de las garrapatas, lo cual permanece aún
por ser investigado. Finalmente, todas las proteínas inmunogénicas identificadas muestran
un escenario de aparente complejidad y elevan la necesidad de un acercamiento racional e
integrador para seleccionar y diseñar mejores vacunas candidatas anti-garrapatas.

67
10. PERSPECTIVAS
Se busca a corto y mediano plazo el poder obtener proteínas recombinantes de las proteínas
inmunogénicas identificadas en el presente trabajo. Comenzando con las proteínas del
Ovario, de las cuales se puede diseñar el gen sintético, u otra opción es mediante el diseño
de péptidos sintéticos, para lo cual se realizó un análisis de predicción de péptidos antigénicos
mediante el programa [PREDICTED ANTIGENIC PEPTIDES;
http://imed.med.ucm.es/Tools/antigenic.pl].
Figura 26. Esquema de predicción de péptidos antigénicos de la proteína vitelogenina 2 de R. microplus.
En este análisis de la proteína vitelogenina se observaron 62 picos, lo que indica que es
altamente inmunogénica. Por este motivo se podrían diseñar genes sintéticos a partir de la
secuencia de la proteína o péptidos sintéticos. La tabla 8 muestra el nombre de las proteínas
así como el gi, la localización y finalmente los péptidos para sintetizar.
Tabla 8. Secuencia de péptidos para sintetizar
Nombre Proteínas ig Localización Péptidos para sintetizar Bm86 Bm86 ADQ19686 - -ISSICSDFGNEFCRNA
-CDCGEWGAMNKTTR -MYPKLLIKKNSATE
P0 P0 AGQ49465 -282-301 -AAGGGAAAAKPEESKKEEAK
2-Ova-sol Vitelogenina 1 JAC58991 -874-893 -174-193
-GDVVLPVDKELFDTLKTELV -GVSGHCRVTYEVHSLTFLP
3-Ova-Sol Vitelogenina 2 JAC58954 -1720-1739 -1903-1922
-GSSVLVSVMLPKYTGTLLGC -SLADIPTITKSVDVPVDCVP

68
4ab-Ova-
sol
Catepsina de yema
AAX76981.1 -6–25 -243-262
-IMSNLFHSSSEEVGQTIPLN -SQIDSINKALNASKTSGDEY
1-Int-insol Componente factor de
iniciación de transcripción
TFIID putativo
GACK01007464 -39-58 -YRYVTNILEDARLYSTHSKK
2-Int-insol Subunidad alfa F0F1 ATP
sintasa putativa
GACK01004976 254-273 336-355
-STVAQIVKRLTQSDAMKYT -AYPGDVFYLHSRLLERAAKM
1-Ova-
insol
Canal Selectivo aniónico
dependiente al voltaje
mitocondrial
GU994210.1 -223-242 -65-84
-VYDLDMETSVRAKVNNSGQI -EYGLTLKEKWNTDNTLSTE
En la tabla 9 se muestran los diferentes software que se utilizaron para realizar el análisis de
las regiones inmunogénicas de las proteínas, los cuales dan como resultado los péptidos mas
inmunogénicos.
Tabla 9. Programas que se utilizaron para analizar y diseñar los péptidos sintéticos.
Nombre de las Proteínas
Vaxijen 0.5 IEDB Analysis Resource
SVMTrip:Tool to Predict Linear
Antigenic Epitopes
Predicted Antigenic Pedtides
Bm86 0.7616 (PA) 23 (5) 2 33
P0 0.4818 (NA) 17 (1) 2 10
2-Ova-sol 0.4793 (NA) 77 (4) 2 56
3-Ova-Sol 0.4511 (NA) 90 (4) 2 65
4ab-Ova-sol 0.6283 (PA) 13 (5) 2 13
1-Int-insol 0.4722 (NA) 6 (14) 1 14
2-Int-insol 0.4879 (NA) 8 (24) 2 26
1-Ova-insol 0.5360 (PA) 15 (6) 2 13

69
11. REFERENCIAS
Almazán, C., Lagunes, R. y otros (2010). Identification and characterization of Rhipicephalus (Boophilus) microplus candidate protective antigens for the control of cattle tick infestations. Parasitology Research 106 (2): 471–479. Almazan, C., Moreno-Cantu, O. y otros (2012). Control of tick infestations in cattle vaccinated with bacterial membranes containing surface-exposed tick protective antigens. Vaccine 30 (2): 265–272. Alonso-Díaz, M.A., Rodríguez-Vivas, R.I. y otros (2006). Ixodicide resistance of the Boophilus microplus tick to ixodicides. Archivos Medicina Veterinaria 38 (2): 105-113. Antunes, S., Galindo, R., y otros (2012). Functional genomics studies of Rhipicephalus (Boophilus) annulatus ticks in response to infection with the cattle protozoan parasite, Babesia bigemina. International Journal of Parasitology 42 (2): 187–195. Avendaño, L. y Correa, A. (2002). Tick infestations. Parasites, external. Elsevier Science 2210-2215. Badii, M. y Garza, V. (2007). Resistencia en insectos, plantas y microorganismos. CULCYT//Impacto Ecológico 4 (18): 9-25. Baffi, M., Rocha, G. y otros (2007) Identification of point mutations in a putative carboxylesterase and their association with acaricide resistance in Rhipicephalus (Boophilus) microplus (Acari: Ixodidae). Veterinary Parasitology 148 (3-4): 301-309. Baffi, M., Rocha, G. y otros (2008). Esterase enzymes involved in pirethroid and organophosphate resistance in a Brazilian population of Rhipicephalus (Boophilus) microplus (Acar, Ixodidae). Molecular Biochemistry Parasitology 160 (1), 70-73. Barker, S.C. y Murrell, A. (2004). Systematic and evolution of ticks with a list of valid genus and species names. Parasitology 129 Suppl: S15-36. Benavides, E. y Romero, A. (2001). Consideraciones para el control integral de parásitos externos del ganado. Carta FEDEGAN 70, 64-86. Benavides, E., (2008). Evaluación crítica de las estrategias de extensión de alternativas para el control de garrapatas en animales domésticos en América Latina. VI Seminario Internacional de Parasitología Animal, Boca del Río Veracruz, 3 a 5 de septiembre del 2008. pp. 6. Bergamo, E.A., Seixas, A. y otros (2010). Vitellin- and hemoglobin-digesting enzymes in Rhipicephalus (Boophilus) microplus larvae and females. Comparative Biochemistry and Physiology 157 : 326-335. Boldbaatar, D., Umemiya-Shirafuji, R. y otros (2010). Multiple vitellogenins from the Haemaphysalis longicornis tick are crucial for ovarian development. Journal of Insect Physiology 56 (11): 1587-1598. Canales, M., Enríquez, A. y otros (1997). Larges-cale production in Pichia pastoris of the recombinant vaccine Gavac2 against cattle tick. Vaccine 15 (4): 414–422.

70
Chen, J.S., Sappington, T.W. y otros (1997) Extensive sequence conservation among insect, nematode and vertebrate vitellogenins reveals ancient common ancestry. Journal of Molecular Evolution 44 (4): 440-451. Cobon, G., Hungerford, J. y otros (1995). Vaccination against Boophilus microplus: the Australian field experience. Recombinant Vaccines for the Control of Cattle Tick, Havana, Cuba: Elfos Scientiae, pp. 163–176. Conacyt-Sagarpa, (2005). Resistencia de la garrapata Boophilus microplus a los ixodicidas en el sureste de México. Folleto técnico No. 1 Mérida, México. pp. 12. Cossart, P. y Toledo-Arana, A. (2008). Listeria monocytogenes, a unique model in infection biology: an overview. Microbes and Infection 10: 1041-1050. Crampton, A. L., P. Green, y otros (1999). "Monooxygenases Play Only a Minor Role in Resistance to Synthetic Pyrethroids in the Cattle Tick, Boophilus Microplus." Experimental and Applied Acarology 23 (11): 897-905. Cruz-Vazquez, C., Fernandez-Ruvalcaba, M. (2000). Anti-tick repellent effect of Andropogon gayanusgrass on plots of different ages experimentally infested with Boophilus microplus larvae. Parasitología al día 248 (3-4): 88-91. De La Fuente, J. y Kocan K.M. (2006). Strategies for development of vaccines for control of ixodid tick species. Parasite Immunology 28 (7): 275-283. De la Fuente, J. y Merino, O. (2013). Vaccinomics, the new road to tick vaccines. Vaccine 2013, 31 (50): 5923–5929. De la Fuente, J., Almazán, C. y otros (2007). A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Animal Health Research Reviews 8 (1): 23–28. De La Fuente, J., Rodriguez, M. y Garcia-Garcia, J.C. (2000). Immunological control of ticks through vaccination with Boophilus microplus gut antigens. Annals of the New York Academy of Sciences 916: 617–621. De la Fuente, J., Rodriguez, M. y otros (1998). Field studies and cost-effectiveness analysis of vaccination with Gavac (TM) against the cattle tick Boophilus microplus. Vaccine 16 (4): 366–373. De la Fuente, J., Rodríguez, M. y otros (1999). Vaccination against ticks (Boophilus spp.): the experience with the Bm86-based vaccine Gavac. Genetic Analysis 15 (3): 143–148. Errecalde, C., Prieto, G., Lüders, C., García, H., 2003. Naturaleza y control de la quimiorresistencia en ectoparásitos. Revista Colombiana de Ciencias Pecuarias 16 (3): 257-267. Esteves, E., Fogasa A.C. y otros (2009). Antimicrobial activity in the tick Rhipicephalus (Boophilus) microplus eggs: Cellular localization and temporal expression of microplusin during oogenesis and embryogenesis. Developmental and Comparative Immunology 33 (8): 913-919.

71
Estrada-Peña, A. y Venzal, M. (2006). High resolution predictive mapping for Boophilus annulatus and Boophilus microplus (Acari: Ixodidae) in Mexico and Southern Texas. Veterinary Parasitology 142 (3-4): 350-358. FAO (2003). Resistencia a los antiparasitarios: estado actual con énfasis en América Latina. Estudio FAO, Producción y Sanidad Animal 157 pp. 55. FAO (2005). Integrated control programs for ticks on cattle: an examination of some possible components. Queensland: FAO 43-61 pp. FAO (2007). Tick-borne livestock diseases and their vectors. FAO Corporate Repository. Fernandes, E.K.K., Bitencourt, V.R.E.P. y Roberts D.W. (2012). Perspectives on the potential of entomopathogenic fungi in biological control of tick. Experimental Parasitology 130 (3): 300-305. Fragoso, H., Hoshmand, R.P. y otros (1998). Protection against Boophilus annulatus infestations in cattle vaccinated with the B. microplus Bm86-containing vaccine Gavac. Vaccine 16 (2): 1990–1992. Francischetti, I., Sa-Nunes, A. y otros (2010). The role of saliva in tick feeding. Frontiers Bioscience Land Mark Editors14, 2051- 2088. Furlong, J. (1998). Poder infestante de larvas de Boophilus microplus (Acari:Ixididae) em pastagem de Melinis minotiflora, Brachiaria decumbens e Brachiaria mutica. Ciencia Rural, Santa María 28 (4): 635-648. Goldberg, M.B. (2001). Actin-based motility of intracellular microbial pathogens. Microbiology and Molecular Biology Reviews 65 (4): 595-626. Gough, J. M. and D. H. Kemp (1993). Localization of a low abundance membrane protein (Bm86) on the gut cells of the cattle tick Boophilus microplus by immunogold labeling. Journal of Parasitology 79 (6): 900-907. Gouin, E., Welch, M.D. y otros (2005). Actin-based motility of intracellular pathogens. Current Opinion in Microbiology 8: 35-45. Graf, J.F., Gogolewski, R. y otros (2004). Tick control: an industry point of view. Parasitology 129 (Suppl): S427–S442. Graham, O.H. y Hourrigan J.L. (1977). Eradication programs for the arthropod parasites of livestock. Journal of Medical Entomology 13 (6): 629-658. Guerrero, F., Nene V. y otros (2006). Sequencing a new target genome: the Boophilus microplus (Acari: Ixodidae) genome project. Journal of Medical Entomology 43(1): 9-16. Guerrero, F.D., Miller, R.J. y Pérez de León, A.A. (2012). Cattle tick vaccines: many candidate antigens, but will a commercially viable product emerge? International Journal of Parasitology 42 (5): 421–427. Hajdusek, O., Almazan, C. y otros (2010). Characterization of ferritin 2 for the control of tick infestations. Vaccine 28 (17): 2993–2998.

72
Hajdusek, O., Sojka, D. y otros (2009). Knockdown of proteins involved in iron metabolism limits tick reproduction and development. Proceedings of the National Academy of Sciences of the United States of America 106 (4):1033–1038. Hawkes, R., Niday, E. y otros (1982). A dot-immunobinding assay for monoclonal and other antibodies. Analytical Biochemestry. 119 (1): 142-147. He, H., Chen, A.C. y otros (1999). Identification of a point mutation in the para-type sodium channel gene from a pyrethroid-resistant cattle tick. Biochemical Biophysical Research Communications 261 (3): 558-561. Heekin, A., Guerrero, D. y otros (2013). The ovarian transcriptome of the cattle tick, Rhipicephalus (Boophilus) microplus, feeding upon a bovine host infected with Babesia bovis. Parasites and Vectors 6 (1): 276-292. Hemingway, J., Hawkes, N. y otros (2004). The molecular basis of insecticide resistance in mosquitoes. Insect Biochemistry Molecular Biology 34 (7): 653-665. IICA (1980). Reunión de Directores de Saludos Animal. Resandina I, Bogotá, Colombia. Jamroz, R., Guerrero, F. y otros (2000). Molecular and biochemical survey of acaricide resistance mechanisms in larvae from Mexican strains of the southern cattle tick, Boophilus microplus. Journal Insect Physiology 46 (5): 685-695. Jonsson, N. (2006). The productivity effects of cattle tick (Boophilus microplus) infestation on cattle, with particular reference to Bos indicus cattle and their crosses. Veterinary Parasitology 137 (1-2): 1-10. Jonsson, N.N., Matschoss A.L. y otros (2000). Evaluation of tick- GARD (PLUS), a novel vaccine against Boophilus microplus, in lactating Holstein–Friesian cows. Veterinary Parasitology 88 (3-4): 275–285. Kaufman, W. (2010). Ticks: Physiological aspects with implications for pathogen transmission. Ticks Tick Borne Diseases 1 (1), 11-22. Khalil, S.M.S., Donohue, K.V. y otros (2011). Full-length sequence, regulation and developmental studies of a second vitellogenin gene from the American dog tick, Dermacentor variabilis. Journal of Insect Physiology 57 (3): 400-408. Klafke, G.M., Sabatini G.A. y otros (2006). Larval immersion tests with ivermectin in populations of the cattle tick Rhipicephalus (Boophilus) microplus (Acari: Ixodidae) from State of Sao Paulo, Brazil. Veterinary Parasitology 142 (3): 386-390. Kongsuwan, K., Josh P., y otros (2010). Activation of several key components of the epidermal differentiation pathway in cattle following infestation with the cattle tick, Rhipicephalus (Boophilus) microplus. International Journal for Parasitology 40 (4): 499-507. Larregina A.T. y Falo L.D. (2005). Changing paradigms in cutaneous immunology: adapting with dendritic cells. Journal of Investigative Dermatology 124 (1): 1–12.

73
Logullo, C., Da Silva, V. (1998). Isolation of an aspartic proteinase precursor from the egg a hard tick, Boophilus microplus. Parasitology 116 (6): 525-532. Logullo, C., Moraes, J. y otros (2002). Binding and storage of heme by vitellin from cattle tick, Boophilus microplus.Insect Biochemistry and Molecular Biology 32 (12): 1805-1811. Lowry, O.H., Rosebrough, N.J. y otros (1951). Protein measurement with the Folin phenol reagent. Journal of Biological Chemistry 193 (1): 265-275. Mackinnon, M.J., Meyer, K., Hetzel, D.J.S. (1991). Genetic variation and covariation for growth, parasite resistance and heat tolerance in tropical cattle. Livestock Production Science 27 (2-3): 105-122. Martínez-Velázquez, M., Castillo-Herrera y otros (2011). Acaricidal effect and chemical composition of essential oils extracted from Cuminum cyminum, Pimenta dioica and Ocimum basilicum against the cattle tick Rhipicephalus (Boophilus) microplus (Acari: Ixodidae). Parasitology Research 108 (2): 481-487. Martínez-Velázquez, M., Rosario-Cruz, R. y otros (2011). Acaricidal effect of essential oils from Lippia graveolens (Lamiales: Verbenaceae), Rosmarinus offcinalis (Lamiales: Lamiaceae), and Allium sativum (Liliales: Liliaceae) Against Rhipicephalus (Boophilus) microplus (Acari: Ixodidae). Journal of Medical Entomology 48 (4): 822-827. Martins, J. R. y Furlong, J. (2001). Avermectin resistance of the cattle tick Boophilus microplus in Brazil. Veterinary Record 149(2): 64. Martins, J.R., Evans D.E. y otros (2002). Partial Strategic Tick Control within a Herd of European Breed Cattle in the State of Rio Grande do Sul, Southern Brazil. Experimental and Applied Acarology 27 (3): 241-251. Miller, R.J., Davey R, B, y George J.E. (2005). First Report of Organophosphate-resistant Boophilus microplus (Acari: Ixodidae) within the United States. Journal of Medical Entomology 42 (5): 912-917. Mulenga, A., Sugimoto, C. y Onuma, M. (2000) Issues in tick vaccine development: identification and characterization of potential candidate vaccine antigens. Microbes and Infections 2 (11): 1353–1361. Nari, A. y Fiel, C. (1994). Enfermedades parasitarias de importancia económica en bovinos. Bases epidemiológicas para su prevención y control en Argentina y Uruguay. Montevideo, Uruguay. Editorial Hemisferio Sur. pp: 289-299. Ndumu, P.A., George, J.P.D. y Choudhury, M.K. (1999). Toxicity of neem seed oil (Azadirachta indica) against the larvae of Amblyomma variegatum, a three-host tick in cattle. Phytotherapy Research 13 (6): 532-534. Neira, J., Carvajal, L. y otros (2009) Evaluación del efecto de la tintura de tabaco (Nicotiana tabacum) en el control biológico de La garrapata. Revista Colombiana de Ciencias Pecuarias 3 (22): 551-552.

74
Nuttall, P.A., Trimnell, A.R. y otros (2006). Exposed and concealed antigens as vaccine targets for controlling ticks and tick-borne diseases. Parasite Immunology 28 (4): 155–163. Ojeda-Chi, M.M., Rodríguez-Vivas, R.I. y otros (2011). Control de Rhipicephalus microplus (Acari: Ixodidae) mediante el uso del hongo entomopatógeno Metarhizium anisopliae (Hypocreales: Clavicipitaceae). Revista Mexicana de Ciencias Pecuarias 2 (2): 177-192. P. Willadsen (1997). Novel vaccines for ectoparasites. Veterinary Parasitology 71 (2-3): 209–222. Padgett, K.A., Casher, L.E. y otros (2009). Effect of prescribed fire for tick control in California chaparral. Journal of Medical Entomology 46 (5): 1138-45. Parizi, L.F., Githaka, N.W. y otros (2012). The quest for a universal vaccine against ticks: cross-immunity insights. Veterinary Journal 194 (2): 158–165. Parizi, L.F., Pohl, P.C. y otros (2009). New approaches toward anti-Rhipicephalus (Boophilus) microplus tick vaccine. Brazilian Journal of Veterinary Parasitology 18 (1): 1–7. Parizi, L.F., Utiumi, K.U., y otros (2011). Cross immunity with Haemaphysalis longicornis glutathione S-transferase reduces an experimental Rhipicephalus (Boophilus) microplus infestation. Experimental Parasitology 127(1): 113–118. Pathak, D., Mathur, V.C. y otros (2004). In vitro effect of indigenous plant extracts on ixodid ticks of small ruminants. Indian Journal of Animal Sciences 74 (6): 616-617. Peter, R.J., Van den Bossche, P. y otros (2005). Tick, fly, and mosquito control–Lessons from the past, solutions for the future. Veterinary Parasitology 132 (3-4): 205–215. Prullage, J., Tran, H. y otros (2011). The combined mode of action of fipronil and amitraz on the motility of Rhipicephalus sanguineus. Veterinary Parasitology 179 (4): 302-310. Rachinsky, A., Guerrero, F.D. y otros (2007). Differential protein expression in ovaries of uninfected and Babesiainfected southern cattle ticks, Rhipicephalus (Boophilus) microplus. Insect Biochemistry and Molecular Biology 37(12): 1291-1308. Rachinsky, A., Guerrero, F.D. y otros (2008). Proteomic profiling of Rhipicephalus (Boophilus) microplus midgut responses to infection with Babesia bovis. Veterinary Parasitology 152(3): 294-313. Rand, K.N., Moore T. y otros (1989). Cloning and expression of a protective antigen from the cattle tick Boophilus microplus. Proceedings of the National Academy of Sciences 86 (24): 9657-9661. Redondo, M., Fragoso, H. y otros (1999). Control of chemically resistant Boophilus microplus populations on grazing cattle vaccinated with Gavac2 in México. Experimental and Applied Acarology 23 (10): 841–849. Rodríguez Valle, M. (2000). Respuesta inmunológica contra garrapatas. Biotecnología Aplicada 17 (4): 215-220.

75
Rodríguez, M., Massard, C.L. y otros (1995). Effect of vaccination with a recombinant Bm86 antigen preparation on natural infestations of Boophilus microplus in grazing dairy and beef pure and crossbred cattle in Brazil. Vaccine 13 (18): 1804–1808. Rodríguez, M., Penichet, M. y otros (1995). Control of Boophilus microplus populations in grazing cattle vaccinated with a recombinant Bm86 antigen preparation. Veterinary Parasitology 57 (4): 339–349. Rodríguez-Vivas, R., Arieta-Román, R. y otros (2010). Uso de lactonas macrocíclicas para el control de la garrapata Rhipicephalus (Boophilus) microplus en el ganado bovino. Archivos Medicina Veterinaria 42, 115-123. Rodríguez-Vivas, R., Rosado, J. y otros 2008. Situación actual de la resistencia a los ixodicidas de Riphicephalus (Boophilus) microplus en el sureste de México: diagnóstico, epidemiología y control. VI Seminario Internacional de Parasitología Animal, Boca del Río Veracruz, México. pp. 20. Rodríguez-Vivas, R.I., Quiñones, A.F. y Fragoso, S.H. (2005) Epidemiología y control de la garrapata Boophilus en México. Enfermedades de importancia económica en producción animal. McGraw Hill, Ciudad de México, México pp: 571-592. Rosado-Aguilar, J.A., Aguilar-Caballero, A. y otros (2010). Acaricidal activity of extracts from Petiveria alliacea (Phytolaccaceae) against the cattle tick, Rhipicephalus (Boophilus) microplus (Acari: ixodidae). Veterinary Parasitology 168 (3-4):299-303. Rosado-Aguilar, J.A., Aguilar-Caballero, A.J. y otros (2010) Screening of the acaricidal effcacy of phytochemical extracts on the cattle tick Rhipicephalus (Boophilus) microplus (acari: ixodidae) by larval immersion test. Tropical and Subtropical Agroecosystem 12 (1): 417-422. Rosario-Cruz, R., Miranda-Miranda, E. y otros (1997). Detection of esterase activity in susceptible and organophosphate resistant strains of the cattle tick Boophilus microplus. Bulletin Entomology Research 87 (2): 197. Sangster, N.C. (2001). Managing parasiticide resistance. Veterinary Parasitology 98 (1-3): 89-109. Sardá-Ribeiro, V.L.C., Tiogo, E. y otros (2007). Acaricidal propieties of extracs from the aerial parts of Hypericum polyanthemun on the cattle tick Boophilus microplus. Veterinary Parasitology 147 (1-2): 199-203. Seixas, A., Oliveira, P. y otros 2012. Rhipicephalus (Boophilus) microplus embryo proteins as target for tick vaccine. Veterinary Immunology and Immunopathology 148 (1-2): 149–156. Shahein, Y.E., A.E. EI-Hakim y otros (2008). Molecular cloning, expression and characterization of a functional GSTmu class from the cattle tick Boophilus annulatus. Veterinary Parasitology 152 (1-2): 116-126. Shimizu, S., Mashashi, N. y otros (1999). Bcl-2 family proteins regulate the release of apoptogenic cytochrome c by the mitochondrial channel VDAC. Nature 399 (483-487). Taheri, M., Nabian, S. y otros (2014). Study of vitellogenin in Boophilus annulatus tick larvae and its immunological aspects. Tropical Biomedicine 31 (3): 398-405.

76
Tan, A.A.W.L., Francischetti, I.M.B. y otros (2012). Laboratory of Malaria and Vector Research. National Institute of Allergy and Infectious Diseases Rockville USA. Taylor, M., Coop, M.A. y Wall R.L. (2007). Veterinary Parasitology. Third edition. Blackwell Publishing. London, UK. 697 p. Tellam, R.L., Kemp, D. y otros (2002). Reduced oviposition of Boophilus microplus feeding on sheep vaccinated with vitellin. Veterinary Parasitology 103 (1-2): 141-156. Temeyer (2004). Organophosphate resistance in mexican strains of Boophilus microplus: a major threat to the U.S. cattle industry. SAAS Bulletin. Biochemistly and Biotechnology 17: 43-51. Thompson, D.M., Khalil S.M.S. y otros (2007) Sequence and the developmental and tissue-specific regulation of the first complete vitellogenin messenger RNA from ticks responsible for heme sequestration. Insect Biochemistry and Molecular Biology 37 (4): 363-374. Tirloni, L., Reck, J. y otros (2014). Proteomic Analysis of Cattle Tick Rhipicephalus (Boophilus)
microplus Saliva: A Comparison between Partially and Fully Engorged Females. PlosOne 9 (4): e94831. Trimnell, A.R., Davies, G.M., y otros (2005). A cross-reactive tick cement antigen is a candidate broad-spectrum tick vaccine. Vaccine 23 (34), 4329–4341. Towbin, H., Staehelien, T. y otros (1979). Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Procedures in Natural Academy of Science. U.S.A. 76 (1979) 4350-4354. Untalan, P.M., Guerrero, F.D. y otros (2005). Proteome analysis of abundantly expressed proteins from unfed larvae of the cattle tick, Boophilus microplus. Insect Biochemistry and Molecular Biology 35 (2): 141-151. Utech, K.B., Wharton, R.H. y Kerr, J.D. (1978). Resistance to Boophilus microplus (Canestrini) in different breeds of cattle. Australian Journal of Veteterinary Research 29 (4): 163-81. Verrisimo, J.C. y Machado, S.G. (1995). Fase de vida livre do ciclo evolutivo do carrapato Boophilus microplus. Zootecnia 33 (1): 41-53. Wilkinson, P.R. (1970). Factors affecting the distribution and abundance of the cattle tick in Australia: Observations and hypotheses. France Acarology 12 (3): 492-508. Willadsen P, Bird P, Cobon GS and Hungerford J (1995). Commercialisation of a recombinant vaccine against Boophilus microplus. Parasitology 110: S43–S50. Willadsen, P. (2001). The molecular revolution in the development of vaccines against ectoparasites. Veterinary Parasitology (2001) 101 (3-4): 353–367. Willadsen, P. (2004). Anti-tick vaccines. Parasitology 129 (Suppl.): S367–S387. Willadsen, P. y Kemp, D.H. (1988). Vaccination with ‘concealed’ antigens for tick control. Parasitology Today 4 (7): 196–198.

77
Wilson, L.J., Sutherst, R.W. y Kerr, J.D. (1989). Trapping of larvae of the cattle tick, Boophilus microplus, by Stylosanthes scabra under field grazing conditions. Australian Journal of Agricultural Research, 40 (6): 1301-1308.

78
12. APENDICE
13. Electroforesis de Proteínas SDS-PAGE 1D
Este protocolo es usado para separar proteínas de acuerdo a su peso molecular. Estas migran hacia el ánodo, a través de la malla de poliacrilamida al aplicar una corriente eléctrica.
1. Ensamblar la celda con los vidrios y los espaciadores del equipo de mini geles
Mini-PROTEAN® Tetra Electrophoresis System de Bio-Rad y montarlo sobre el
contenedor porta vidrios para el vaciado de los geles.
2. Preparar la solución del gel separador, a la adecuada concentración de acrilamida.
Inmediatamente, transferir esta solución a la celda de vidrios. Con la ayuda de una
micropipeta de 5mL verterlo por una esquina del espaciador. Llenar (aprox. 5 ml
de solución) hasta dejar un espacio superior de aproximadamente 1 cm (Para el gel
concentrador).
3. Permitir la polimerización en al menos 30 min.
4. Preparar el gel concentrador. Inmediatamente transferir la solución a la celda de
vidrios. Insertar el peine (según el número. de pozos que se requieran) procurando
que no queden burbujas entre el peine y el gel.
5. Permitir la polimerización en al menos 20 min.
6. Una vez polimerizado el gel, remover cuidadosamente los peines y montarlos en el
corazón de los electrodos de la cámara de electroforesis. Adicionar el amortiguador
de corrida 1× (en Soluciones), tanto en el compartimiento interno (aprox. 100 ml)
como en la parte baja de la cámara (hasta donde indique la marca en la cámara).
7. Preparar las muestras biológicas. Adicionar Amortiguador de muestra 1× o 2×,
según sea el caso. En caso de muestras líquidas adicionar Buffer 2× a una
proporción 1:1. Para pastillas de proteínas precipitadas, adicionar de 30 a 40 µl de
amortiguador de muestra 1× (en Soluciones). Hervir 5 min y centrifugar antes de
cargar el sobrenadante a los pozos del gel de poliacrilamida.
8. Adicionar las proteínas comerciales de peso molecular estándar en uno de los
pozos.

79
9. Conectar los electrodos de la cámara de electroforesis a una fuente de poder y correr
durante 1 h a 120 voltios aprox. Terminar la corrida cuando la marca del azul de
bromofenol, contenido en el amortiguador de muestra, cruce prácticamente todo el
gel separador.
10. Una vez terminada la corrida, desensamblar la cámara y recuperar los geles.
Visualizar las proteínas separadas tiñendo el gel con solución colorante de azul de
Coomassie (en Soluciones).

80
13.1. Protocolo para geles de separación y concentración de proteínas en
geles de poliacrilamidaa.
Gel Separador:
___________________________________________________
Concentración final de acrilamida en geles separadores (%)b
Soluciones Stock (ml) 5 6 7 7.5 8 9 10 12 13 15
Acrilamida 30%/
bisacrilamida 0.8%
4× Tris-Cl/SDS pH 8.8
Agua destilada
Persulfato de amonio al 10%
TEMED
2.50
3.75
8.75
0.05
0.01
3.00
3.75
8.25
0.05
0.01
3.50
3.75
7.75
0.05
0.01
3.75
3.75
7.50
0.05
0.01
4.00
3.75
7.25
0.05
0.01
4.50
3.75
6.75
0.05
0.01
5.00
3.75
6.25
0.05
0.01
6.00
3.75
5.25
0.05
0.01
6.50
3.75
4.75
0.05
0.01
7.50
3.75
3.75
0.05
0.01
Gel Concentrador:
- Acrilamida 30% /bisacrilamida 0.8%
- 4× Tris-Cl/SDS, pH 6.8
- Agua destilada (mezclar suavemente)
- Persulfato amonio 10%
- TEMED
0.65 ml
1.25 ml
3.05 ml
0.025 ml
0.005 ml
aEsta preparación equivale a 15 ml de solución para gel de separación, la cual alcanza para 3 minigeles con un grosor de 1.00 mm. bEl porcentaje del gel de poliacrilamida depende de los pesos moleculares de las proteínas a separar.
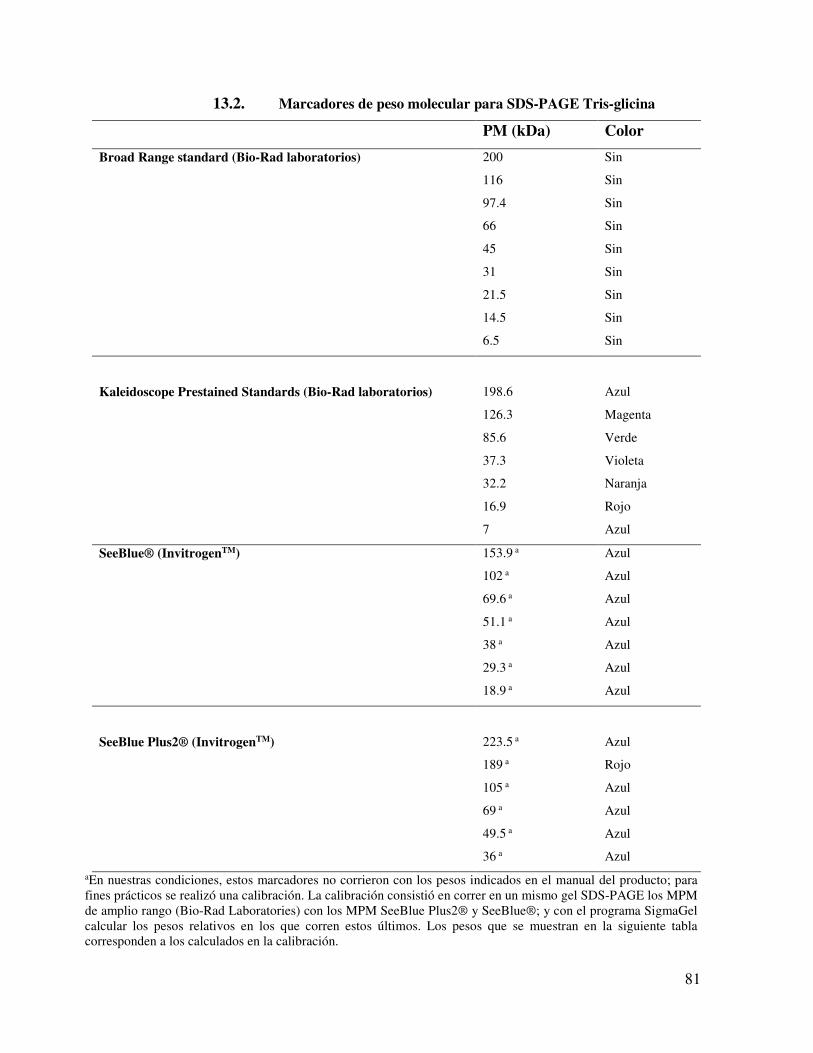
81
13.2. Marcadores de peso molecular para SDS-PAGE Tris-glicina
PM (kDa) Color
Broad Range standard (Bio-Rad laboratorios) 200 Sin
116 Sin
97.4 Sin
66 Sin
45 Sin
31 Sin
21.5 Sin
14.5 Sin
6.5 Sin
Kaleidoscope Prestained Standards (Bio-Rad laboratorios) 198.6 Azul
126.3 Magenta
85.6 Verde
37.3 Violeta
32.2 Naranja
16.9 Rojo
7 Azul
SeeBlue® (InvitrogenTM) 153.9 a Azul
102 a Azul
69.6 a Azul
51.1 a Azul
38 a Azul
29.3 a Azul
18.9 a Azul
SeeBlue Plus2® (InvitrogenTM) 223.5 a Azul
189 a Rojo
105 a Azul
69 a Azul
49.5 a Azul
36 a Azul
aEn nuestras condiciones, estos marcadores no corrieron con los pesos indicados en el manual del producto; para fines prácticos se realizó una calibración. La calibración consistió en correr en un mismo gel SDS-PAGE los MPM de amplio rango (Bio-Rad Laboratories) con los MPM SeeBlue Plus2® y SeeBlue®; y con el programa SigmaGel calcular los pesos relativos en los que corren estos últimos. Los pesos que se muestran en la siguiente tabla corresponden a los calculados en la calibración.

82
13.3. Carga de muestra apropiada para tinción con Coomassie y plata
para tiras IPG de 7cM
pH Carga de muestra apropiada (µg de proteína) Tinción con plataTinción con Coomassie
4-7 4-8 20-120 6-11 8-16 40-240
3-10, 3-10(NL) 2-6 10-60
13.4. Isoelectroenfoque
El siguiente procedimiento esta explicado para cada tira IPG, para el equipo utilizado
(Ettan IPGphor II de GE Healthcare) se pueden correr simultáneamente hasta 12 tiras IPG.
El protocolo de corrida, que depende de las características de la muestra y el tamaño de
tira IPG, debe ser basado en las especificaciones del manual y del instructivo de las tiras
IPG. El protocolo utilizado en este trabajo esta descrito en la Tabla 6.
1. Preparar la mezcla para rehidratación con la cantidad la de muestra adecuada.
(determinado empíricamente), y llevar a un volumen de 125µL con la solución de
rehidratación.
2. Sobre una canaleta de una placa para western blot (preferentemente utilizar el
strip holder, que se vende por separado), verter la mezcla de rehidratación
preparada.
3. Tomar la tira IPG (almacenadas a -20°C) y retirarle la cubierta de plástico.
4. Colocar la tira con el gel expuesto hacia abajo, sobre la mezcla de rehidratación.
5. Verter encima aceite mineral para evitar evaporación y dejar rehidratar toda la
noche.
6. Tomar la tira IPG y colocarla gel arriba sobre una de las canaletas de la charola
Manifold (Manifold Tray), empezando por la canaleta número uno.
7. Llenar la charola con aceite mineral hasta que cubra las canaletas por completo.
8. Colocar un parche para electrodo (electrode pad), previamente rehidratado con
150µL de agua bidestilada en cada extremo de la tira. Aproximadamente 1/3 del
parche debe de estar en contacto con el gel de la tira.
9. Colocar los dos electrodos sobre los parches y cerrar.
10. Programar el protocolo de corrida
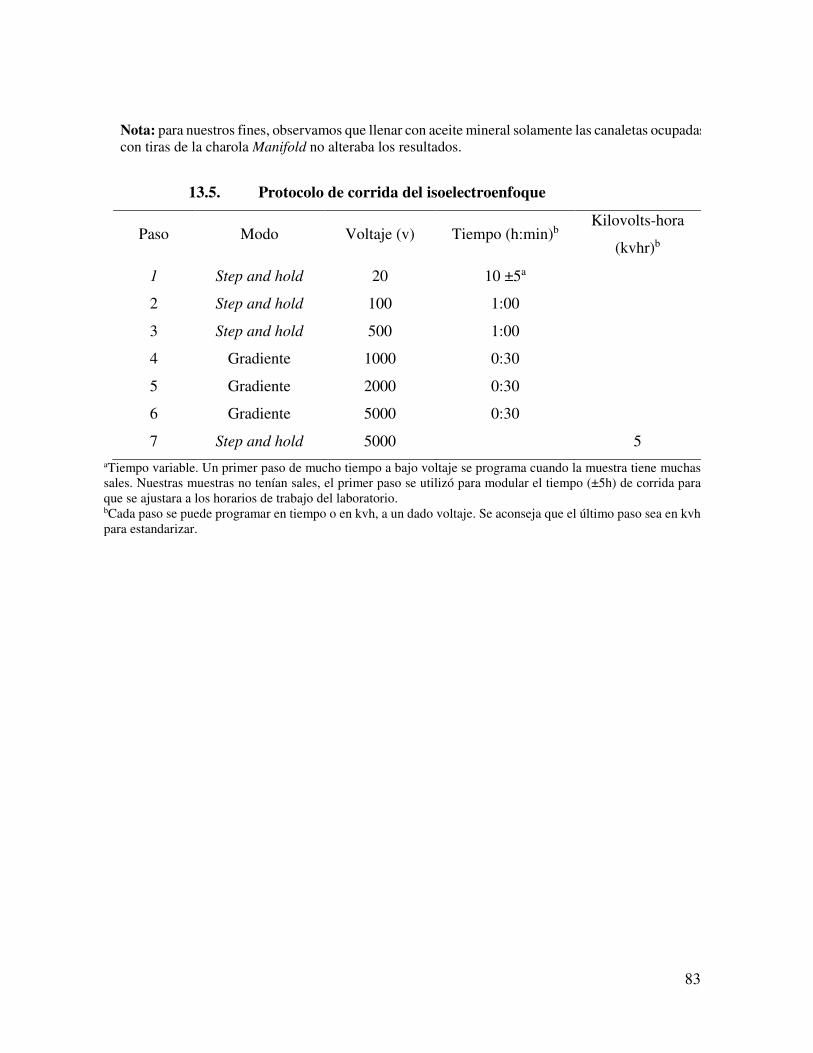
83
Nota: para nuestros fines, observamos que llenar con aceite mineral solamente las canaletas ocupadascon tiras de la charola Manifold no alteraba los resultados.
13.5. Protocolo de corrida del isoelectroenfoque
Paso Modo Voltaje (v) Tiempo (h:min)b Kilovolts-hora
(kvhr)b
1 Step and hold 20 10 ±5a
2 Step and hold 100 1:00
3 Step and hold 500 1:00
4 Gradiente 1000 0:30
5 Gradiente 2000 0:30
6 Gradiente 5000 0:30
7 Step and hold 5000 5 aTiempo variable. Un primer paso de mucho tiempo a bajo voltaje se programa cuando la muestra tiene muchas sales. Nuestras muestras no tenían sales, el primer paso se utilizó para modular el tiempo (±5h) de corrida para que se ajustara a los horarios de trabajo del laboratorio. bCada paso se puede programar en tiempo o en kvh, a un dado voltaje. Se aconseja que el último paso sea en kvh para estandarizar.

84
13.6. Segunda Dimensión
1. Después del isoelectroenfoque, equilibrar la tira IPG en 1mL de amortiguador de
equilibrio y DTT (Di-tio-trietol) al 1%(p/v) 15 min
2. Realizar un segundo paso de equilibrio con amortiguador de equilibrio y
iodoacetamida al 2.5%(p/v) durante 15 min.
3. Preparar un gel de poliacrilamida como descrito en la sección de electroforesis en
geles de poliacrilamida SDS-PAGE (1-D) con un peine preparativo.
4. Colocar la tira dentro del pozo preparativo, con el extremo básico del lado del pozo
para marcadores de peso molecular y el gel, haciendo contacto con uno de los dos
vidrios de la celda.
5. Colocar 4-7µL de marcadores de peso molecular de kaleidoscopio, 2-3µL de
SeeBlue Plus2® o SeeBlue®, o 1.5-3µL de Broad Range.
6. Montar los geles en la cámara y correr como descrito anteriormente.
13.7. Western Blot
1. Preparar un gel SDS-PAGE con las proteínas que se desea separar.
2. Cortar un recuadro de membrana de nitrocelulosa del tamaño aproximado del gel, con
especial cuidado de no maltratar ni ensuciar la membrana, utilizar guantes y procurar
no tocarla.
3. En una cámara fría colocar 1L de buffer de transferencia, previamente enfriado a 4°C.
4. Montar dentro de la prensa del equipo de electrotransferencia los componentes en el
siguiente orden: 1) esponja 2)papel 3)gel 4)membrana 5)papel 6)esponja.
5. Cerrar la prensa asegurándose que no queden burbujas de aire entre el gel y la
membrana, y de haber montado el gel de tal manera que los marcadores de peso
molecular queden fijados en el lado izquierdo de la membrana.
6. Insertar la prensa en el cassette, dentro de la cámara de electrotransferencia, en tal
posición que las proteínas del gel migren hacia el polo positivo situado detrás de la
membrana, para que queden fijas en ella.
7. Colocar la unidad de enfriamiento, previamente enfriada a -20°C.
8. Conectar la cámara de electrotransferencia en la fuente de poder y programar la
corrida a 100 volts durante 1hora.

85
9. Terminada la corrida marque la membrana, antes de mover el gel, en los extremos del
gel.
10. Bloquear con leche al 5% en PBS-Tween 20 al 0.05% toda la noche a 4°C y, en
agitación.
11. Lavar con PBS-Tween 20 al 0.1% 5 veces a temperatura ambiente con agitación
manual vigorosa.
12. Incubar toda la noche, a 4°C y en agitación, con el anticuerpo primario (antisueros de
bovino) diluido en leche al 5% en PBS-Tween 20 0.05%.
13. Lavar con PBS-Tween 20 al 0.1% 5 veces a temperatura ambiente con agitación
manual vigorosa.
14. Colocar el anticuerpo secundario ratón anti-IgG bovino acoplado a peroxidasa de
rábano picante, en dilución 1:3000 (Bio-Rad) en leche al 2.5% en PBS-Tween 20 al
0.05%.
15. Incubar durante 2.5 horas a temperatura ambiente en agitación.
16. Lavar con PBS-Tween 20 al 0.1% 5 veces a temperatura ambiente con agitación
manual vigorosa.
17. Preparar las soluciones por separado, primero la A y luego la B; tienen que ser
preparadas al momento del revelado.
18. Mezclar las soluciones (adicionar la A en la B).
19. Adicionar la mezcla en la membrana de nitrocelulosa
20. Revelar el color entre 5-30 minutos.
21. Parar la reacción lavándola muy bien con agua bidestilada.
22. Dejar la secar a temperatura ambiente.
23. Almacenar en protección de la luz.

86
13.8. Revelado de western blot con el método de peroxidasa
Volumen final
(mL) Solución A Solución B
HRP (mg) Metanol (mL) PBS (mL) Perhidrol (µL)
120 60 20 100 120
60 30 10 50 60
30 15 5 25 30
15 7.5 2.5 12.5 15
13.9. Soluciones
13.10. Solución amortiguadora de PBS pH 7.4 (Phosphate buffer saline)
Por cada 200 mL de amortiguador, disolver una tableta de PBS 7.4 de Invitrogen™.
13.11. Acrilamida al 30% /bisacrilamida al 0.8%
Acrilamida (Bio-Rad) 29.2 g - Disolver en 40 ml de agua destilada. Una vez
disuelto aforar hasta 100 ml.
- Filtrar a través de papel filtro “Whatman” no. 1.
- Almacenar a 4°C hasta 1 mes, previamente
protegido de la luz.
Bisacrilamida (Bio-Rad) 0.8 g
13.12. Tris-Cl/SDS 4×, pH 8.8 (1.5 M Tris-Cl, 0.4% SDS)
Trizma-base (Sigma) 9.08g - Disolver en 20 ml de agua destilada y ajustar a
pH 8.8 con HCl.
- Aforar a un volumen final de 50 ml con agua
destilada.
- Filtrar la solución a través de un filtro de 0.45 µm.
- Almacenar a 4°C.
SDS (Sigma) 0.2 g
13.13. Tris-HCl/SDS 4×, pH 6.8 (0.5 M Tris-HCl, 0.4% SDS)
Trizma-base (Sigma) 3.025g - Disolver en 20 ml de agua destilada y ajustar a pH
6.8 con HCl. SDS (Sigma) 0.2 g

87
- Aforar a un volumen final de 50 ml con agua
destilada.
- Filtrar la solución a través de un filtro de 0.45 µm.
- Almacenar a 4°C.
13.14. Persulfato de
amonio al 10%
13.15. SDS-amortiguador 2×
10 mg (0.01 g) en 1 ml de agua
destilada.
Trizma-base
SDS
1.52 g
2 g
Nota: Almacenar en alícuotas hasta 2 semanas a -20 °C.
Disolver en 50 ml de agua destilada y ajustar el pH a
6.8.
Posteriormente adicionar:
Glicerol 20 mL
Agua destilada (aforar a 100 mL)
13.16. Amortiguador de muestra 1× (BM 1×) (Muestras sólidas)
SDS-amortiguador 2× 500 µL
β-mercaptoetanol 5% 50 µL
Azul de bromofenol (stock 10 mg/mL) 50 µL
Agua desionizada 400 µL
13.17. Amortiguador de muestra 2× (BM 2×) (Muestras líquidas)
SDS-amortiguador 2× 400 µL
β-mercaptoetanol 5% 50 µL
Azul de bromofenol (stock 10 mg/mL) 50 µL
13.18. Amortiguador de
electroforesis 5×(Stock)
13.19. Amortiguador de
electroforesis 1×.
Glicina (Sigma) 72 g Amortiguador 5× stock 100 ml
Trizma-base (Sigma) 15.1 g Agua destilada 400 ml
SDS (Sigma) 5.0 g
Aforar con agua destilada a 1 litro
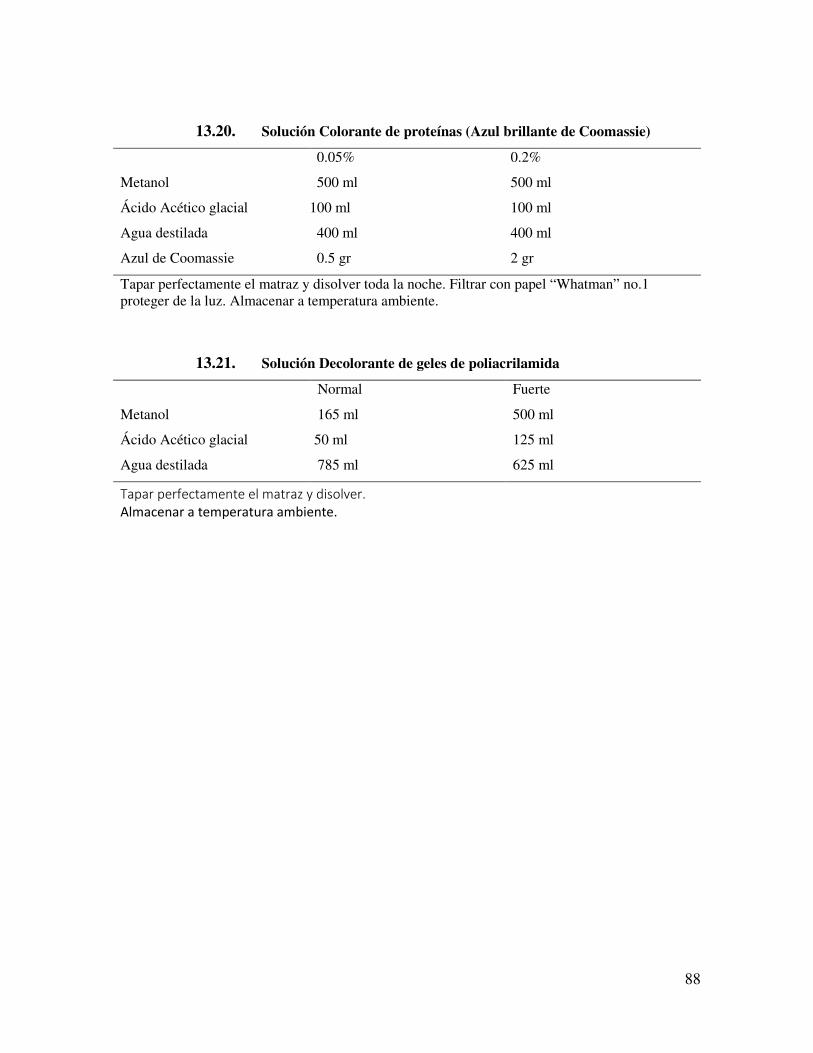
88
13.20. Solución Colorante de proteínas (Azul brillante de Coomassie)
0.05% 0.2%
Metanol 500 ml 500 ml
Ácido Acético glacial 100 ml 100 ml
Agua destilada 400 ml 400 ml
Azul de Coomassie 0.5 gr 2 gr
Tapar perfectamente el matraz y disolver toda la noche. Filtrar con papel “Whatman” no.1 proteger de la luz. Almacenar a temperatura ambiente.
13.21. Solución Decolorante de geles de poliacrilamida
Normal Fuerte
Metanol 165 ml 500 ml
Ácido Acético glacial 50 ml 125 ml
Agua destilada 785 ml 625 ml
Tapar perfectamente el matraz y disolver.
Almacenar a temperatura ambiente.

89
13.22. Solución Equilibradora
SDS de 2D
13.23. Amortiguador de
lisis para 2D
Tris-HCl 0.5 M, pH 6.8 (1.5 gr en 25 ml de agua)
12.5 ml Tris HCl pH 7.8 200µL
SDS 10% (10 gr en 100 ml de agua )
23 ml EDTA 200µL
β-mercaptoetanol
5 ml Urea 9.6g
Glicerol 10%
8 ml DTT 154mg
Azul de Bromofenol 0.05% (0.005 gr en 10 ml de agua).
2.5 ml
Glicerol 2mL
Agua destilada 49 ml Chaps 600mg
Anfolinas 3mL
13.24. Amortiguador de Transferencia
Trizma-base 25 mM
Glicina 190 mM
Metanol 20 %
3.03 gr
14.25 gr
200 ml
Disolver primeramente el trizma y la glicina. Una vez disuelto agregar el metanol y finalmente, aforar a 1 litro con agua destilada. Enfriar a 4°C, hasta su uso. Nota: No es necesario ajustar el pH.

90
13.25. ELISA
Sensibilización de la placa 1.- Fijar el Ag a la placa a razón de 0.25µg por pozo en buffer de carbonatos pH 9.6, en un volumen de 50 µl por pozo. Incubar a 4 °C toda la noche. 2.- Lavar 5 veces con 100 µl de PBS-Tween-20 al 0.05% Bloqueo 3.- Colocar 50 µl de la solución de bloqueo (SFB 5% v/v en 0.05% Tween-20/PBS) 4.- Lavar 5 veces con 100 µl 5% leche en 0.05% Tween-20/PBS. Reacción Ag-Ac 5.- Colocar 50 µl por pozo del suero problema diluido 1:100 en PBS-Tween. En el blanco colocar solamente PBS-Tween. Incubar 1 hr a temperatura ambiente ó 30 min a 37 °C. 6.- Lavar 5 veces con 100 µl de PBS-Tween-20 al 0.05% Conjugado 7.- Diluir el α-IgG bovino 1:2000 con PBS-Tween. Colocar 100 µl por pozo. Incubar 1 hr a temperatura ambiente ó 30 min a 37 °C. 8.- Lavar 6 veces con 100 µl de PBS-Tween-20 al 0.05% Substrato Previamente se coloca a temperatura ambiente el sustrato que se va a utilizar 10.- Colocar 100 µl de sustrato por pozo incubar 10 min a temperatura ambiente. Leer la absorbancia a 405 nm. Detener la reacción con 100 µl de SDS 1%

91
13.26. Tablas con información acerca del material, reactivos, equipos y kit utilizados en la tesis.
REACTIVOS
Acrilamida (SIGMA) Ampicilina sódica (SIGMA) EDTA (SIGMA) Marcador de peso molecular 1KB Plus Protein Standars (BIO-RAD)
Solución de Rehidratación DeStrak™ Rehydration Solution (GE Healthcare)
Aceite mineral (BIO-RAD) Alcohol metílico (SIGMA) Fosfato de potasio monobásico (SIGMA)
Medio LB (SIGMA) Tiras para electroforesis en 2-D ReadyStrip™ IPG Strips 7cm, pH 3-10 (BIO-RAD)
Acetona (SIGMA) Azul brillante G-250 (BIO-RAD)
Fosfato de sodio dibásico (SIGMA)
Membrana de nitrocelulosa Amersham™ Hybond™ -ECL (GE Healthcare)
Trizma base (SIGMA)
Ácido crorhídrico (SIGMA) Azul de bromofenol (SIGMA)
Glicerol (SIGMA) Membrana de nitrocelulosa, 0.45µm (BIO-RAD)
Triton X-100 (SIGMA)*
Ácido acético glacial (SIGMA)
BIS (SIGMA) Glicina (SIGMA) Mercaptoetanol (SIGMA) Trizol (Invitrogen)
Agar (SIGMA) Buffer de carga 10X BlueJuice Gel Loading Gel (Invitrogen)
Histadina Persulfato de amonio (SIGMA) Tween- 20 (SIGMA)*
Agarosa UltraPure™ Agarose (Invitrogen)
Buffer IPG para electroforesis en 2-D IPG Buffer, pH 4-7 (GE Healthcare)
IgG-Bovine Peróxido de hidrógeno (ZUUM)*
Urea (SIGMA)
Agua destilada Buffer IPG para electroforesis en 2-D IPG Buffer, pH 3-10 (GE Healthcare)
Hidróxido de potasio (SIGMA)
SDS (SIGMA) HRP Color Development Reagent, 4 CN (BIO-RAD)
Agua desionizada Cloroformo (SIGMA) Hidróxido de sodio (SIGMA) SYBR® Safe DNA Gel Stain (ThermoFisher SCIENTIFIC)
Agua inyectable (PiSA) Cloruro de potasio (SIGMA) Iodacetamida (GE Healthcare)
TEMED (SIGMA)

92
Alcohol etílico absoluto (SIGMA)
Cloruro de sodio (SIGMA) IPTG (Promega) Tiras para electroforesis en 2-D ReadyStrip™ IPG Strips 7cm, pH 4-7 (BIO-RAD)
Alcohol isopropílico (SIGMA) DTT (GE Healthcare) Leche descremada en polvo Svelty (Nestlé)
MATERIAL
Agujas de las verdes de 0.8x38mm 216x38mM
Fibras para electro-transferencia Mini Trans-Blot Fiber Pads (BIO-RAD)
Matraz Erlenmeyer 200 y 1000 mL KIMAX (KIMBLE)
Portageles
Botes de vidrio con tapa 500 mL (WHEATON)
Filtros para electro-transferencia Mini Trans Blot Filter Paper (BIO-RAD)
Micropipetas PIPETMAN Classic 0.2-2, 0.5-10, 0.2-20, 50-200 y 100-1000 µL (GILSON)
Probeta 10, 250, 500 y 1000 mL KIMAX (KIMBLE) Puntas para micropipetas 0.1-10 y 100-1000µL (NEPTUNE)
Cajas de Petri (plásticos y derivados de Durango S. A.)
Guantes de nitrilo (AMBIDERM)* Gradillas
Microtubos 1.5, 0.5 y 0.2 mL (Axygen)
Puntas para micropipetas 10-100µL
(Corning Inc)
Cubetas para electroporación Sigma-Aldrich electroporation cuvettes (SIGMA-ALDRICH)
Incubadoras para Tiras Mini-Incubation Trays (BIO-RAD)
Papel aluminio (Reynolds Wrap)
Soporte para vidrio para electroforesis de geles de acrilamida MiniPROTEAN 3 System Glass Plates Spacer Plates (BIO-RAD)
Espátula
Jeringas de las rosas 186x1 1 ½ (1.20mMx38mM)
Papel para electrodo para electroforesis en 2-D Paper Wicks IPGphor™ (GE Healthcare)
Vidrios para equipo de electroforesis para geles de acrilamida MiniPROTEAN 3 System Glass Plates Short Plates (BIO-RAD)
Frasco de vidrio con tapa 100, 250, 500 Y 1000 mL (WHEATON)
Matraz aforado 100, 500 y 1000 mL KIMAX (KIMBLE)
Parafilm M (Bermis)
Vacutainer 7 mL rojos sin eparina

93
KIT Columnas de purificación Bio-Scale™ Mini Profinity™ IMAC Cartridges
Champion™ pET101 Directional TOPO® Expression Kit with BL21 Star ™ (DE3) one Shot® Chemically Competent E. coli
Platinum Taq DNA Polimeasa (Invitrogen)
QIAEX II Gel Extraction Kit (150) (QIAGEN)
SuperScript® III First-Strand Synthesis SuperMix
Dream Taq PCR Master Mix (Thermo Scientific)
Platinum PCR SuperMix (Invitrogen)
ProteoSilver™ Silver Stain Kit (SIGMA)
Phusion® High-Fidelity PCR Kit

94
EQUIPOS Agitador Orbital Platform Vari Mix (Thermolyne)
Centrifuga Universal 320 R (Hettich Zentrifugen)
Incubadora con agitación MAxQ 4450 (Thermo SCIENTIFIC)
Termoblock AccuBlock™ Digital Dry Bath (Labnet Internatinal, Inc.)
Agitador Rotor R-06 (SOLBAT) Congelador (TOR-REY) Incubadora Shaker (New Brunswick Scientific)
Transiluminador Fisher Biotech Transilluminadtor FBTI-88 (Fisher Scientific)
Agitador RotoMix Type 48200 (Thermolyne)
Equipo de electroforesis de geles de acrilamida Mini PROTEAN Tetra Cell (BIO-RAD)
Incubadora Shaker KS 3000i control (IKA)
Termoagitador Hotplate Stirrer (Biomega Research Product Inc.)
Autoclave Sterilizer SE510 (Yamato) Equipo de electroforesis de geles de agarosa Mini-Sub Cell GT (BIO-RAD)
Microcentrifuga LaboGene mini (LABOGENE)
Termociclador Biometra T Gadient Thermocycler (Biometra)
Balanza analítica BP 221S (Srtorius)
Equipo de electrotranferencia Mini Trans-Blot Cell (BIO-RAD)
Microcentrifuga refigerada Sorvall Fresco (Sorvall)
Ultracongelador-86 Ultralow Freezer (NUAIRE)
Campana de extracción (Novatech)
Espectrofotómetro NANODROP 2000 Spectrophotometer (Thermo SCIENTIFIC)
Potenciómetro Orion 3 STAR pH Portable (Thermo SCIENTIFIC)
Ultracongelador-86 Ultralow Freezer (NUAIRE)
Campana de flujo laminar (Lumina)
Fuente de poder PowerPac™ HC (BIO-RAD)
Refrigerador (TOR-REY)
Ultracongelador-86 Ultralow Freezer (NUAIRE)
Campana de flujo laminar (Lab Tech)
Fotodocumentador Molecular Imager Gel Doc™ XR+with Image Lab™ Software (BIO-RAD)
Sistema para electroforesis en 2-D Ettan IPGphor 3 (GE Healthacare)
Vortex Select Vortexer (Select BioProducts)
Campana de flujo laminar (VECO)
Horno de microondas (LG) Sonicador Ultrasonic liquid processor XL-2000 SERIES (MISIONIX)

95
14. ANEXOS
14.1. Anexo 1. Tabla 1. Datos de animales con los que se trabajaron en
INIFAP, así como los grupos que se realizaron con estos mismos (tabla 2).
Tabla 1. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias Relación de inventario de semovientes Especie: Bovinos y Ovinos CENID o CIR:CENID PARASITOLOGÍA VETERINARIA No. Fecha de
ingreso No. De arete Raza Edad Peso Sexo Proyecto Tipo de
dieta 1 14-Ago-12 ´052 276653 SA/CS 8 190 M CIATEJ INTEGRAL
2 14-Ago-12 ´051 276641 CS 7 143 M CIATEJ INTEGRAL
3 14-Ago-12 ´050 842416 CS/ CS 6 147 M CIATEJ INTEGRAL
4 14-Ago-12 ´042 276630 SA/CS 7 118 M CIATEJ INTEGRAL
5 14-Ago-12 ´048 282722 CS 7 131 M CIATEJ INTEGRAL
6 14-Ago-12 ´047 307113 LN 18 202 M CIATEJ INTEGRAL
7 14-Ago-12 ´045 276623 CS/ CS 6 138 M CIATEJ INTEGRAL
8 14-Ago-12 ´046 264564 CS 6 128 M CIATEJ INTEGRAL
9 14-Ago-12 ´044 307200 CZ 18 122 M CIATEJ INTEGRAL
10 14-Ago-12 ´043 276621 CS 6 106 M CIATEJ INTEGRAL
Biólogo responsable: Dr. Rodrigo Rosario Propiedad: INIFAP Ubicación: CENIT-PAVET
Tabla 2. Grupos de animales inmunizados Proteínas de intestino solubles Proteínas de intestino insolubles Testigo Arete No. 51 Arete No. 50
Arete No. 44 Arete No. 47
Arete No. 48 Arete No. 45
Proteínas de ovario solubles Proteínas de ovario insolubles Arete No. 46 Arete No. 42
Arete No. 52 Arete No. 43
14.2. Anexo 2. De las proteínas que se identificaron mediante LC-MS/MS se realizó
su análisis para llegar a su peso molecular, punto isoelétrico y nombre, los
cuales se muestran a continuación.
14.3. Proteína 1-Int-Sol
La proteína que se encontró era una proteína de IgG de la cadena pesada [Bos taurus] con un
gi: 91982959, 52 kDa y un IP de 6.5. Dado que esta proteína fue identificada como una
proteína de bovino y no de garrapata, no le realizamos los análisis correspondientes, ya que
no era de nuestro interés. Por estas razones se decidió no continuar con el análisis de las

96
proteínas señaladas como intestino soluble y se procedio a investigar las otras cortadas de los
geles.
14.4. Análisis in silico de la proteína de garrapata de 1-intestino insoluble
14.5. Proteína 1-Int-Insol
Los análisis de LC-MS/MS se realizaron en, La Universidad de California en Davis. Los
cuales nos proporcionaron el siguiente link en el que con el gi de la proteína, se encuentra la
información que a continuación consultamos (organismo, nombre de la proteína, secuencia
de aa., artículo en que fue publicado y secuencia de nucleótidos). Además, en la figura 1 se
muestra una imagen representativa de la plataforma (scaffold) que se utilizó para el análisis
de las proteínas (1-Int-Insol, 2-Int-Insol, 1-Ova-Soluble, 2-Ova-Soluble, 3-Ova-Soluble, 4a-
Ova-Soluble, 4b-Ova-Soluble, 1-Ova-Insoluble, 2-Ova-Insoluble, 3-Ova-Insoluble y 4-Ova-
Insoluble), proporcionado por la Universidad de California.
http://www.ebi.ac.uk/ena/data/view/GACK01007464
Organismo: Rhipicephalus pulchellus
gi: L7M0M0
Product o: Putative transcription initiation factor tfii-d component
MAAPKSTPKDAQVMSAILKDMGIVDYEPRVINQMLEFTYRYVTNILEDARLYSTHSKKRAVDVDDVRLAIQMQAEKTFTS
PPPRDLLMEIARQKNSTPLPLIKANAGPRLPPDRYSLTACNYRLKSAKKPRIQVGLPPQLGLATMQGNSAAGLSKVTPAL
SLNSKTGATLAVVSKPASAPTVTIVSRPATANKMHPTPIIKLSQRPPLNVSTAVSSAAPTPIVQVTAAPQPTPPAPTTTA
AVPATEATTTPPVITTATTEPATQPSVPQPIINIKSETPSEINDKKRKREEDDDYDAAD
Referencias: An insight into the sialotranscriptome of the Zebra tick, Rhipicephalus pulchellus Tan A.A.W.L., Francischetti I.M.B., Manjunatha K.K.R., Ribeiro J.M.C Tan A.A.W.L., Francischetti I.M.B., Manjunatha K.K.R., Ribeiro J.M.C.
Submitted (16-NOV-2012) to the INSDC. Laboratory of Malaria and Vector Research,
National Institute of Allergy and Infectious Diseases, 12735 Twinbrook Parkway
room 2E32D, Rockville, MD 20852, USA
>ENA|GACK01007464|GACK01007464.1 TSA: Rhipicephalus pulchellus RpIx75-677044 mRNA
sequence. : Location:1..900
ATGGCGGCCCCGAAAAGCACGCCGAAGGATGCCCAAGTCATGAGTGCAATACTAAAGGAC
ATGGGAATAGTTGACTACGAGCCGAGAGTCATCAATCAGATGTTGGAGTTCACTTACCGT
TACGTTACCAATATCCTCGAGGACGCCAGGTTGTATTCGACCCATTCGAAAAAGAGAGCC
GTGGACGTCGATGACGTGCGTTTGGCTATTCAAATGCAAGCAGAAAAAACGTTCACGTCC
CCACCACCGAGAGATTTGCTGATGGAAATCGCGCGTCAAAAGAACAGTACGCCGCTGCCG
CTTATCAAAGCGAATGCCGGACCTCGTCTCCCGCCGGATAGGTACAGCCTCACTGCCTGC
AACTATCGGCTGAAGTCTGCAAAAAAGCCAAGGATCCAGGTTGGCCTTCCACCACAGCTT
GGGCTGGCAACCATGCAAGGAAACAGTGCAGCAGGTTTATCGAAAGTAACACCTGCGTTG
AGCCTGAACAGCAAGACAGGTGCCACACTCGCCGTGGTCAGCAAGCCGGCATCTGCACCA
ACGGTGACAATTGTGTCACGACCTGCCACTGCCAACAAAATGCATCCAACACCCATCATC

97
AAACTCTCTCAACGGCCACCCCTCAATGTCAGCACAGCAGTTTCATCAGCAGCACCCACA
CCCATTGTGCAAGTGACAGCTGCCCCGCAGCCTACACCCCCAGCTCCCACTACTACTGCA
GCAGTGCCTGCAACTGAAGCTACCACCACCCCACCAGTCATCACCACAGCTACCACAGAA
CCTGCCACTCAGCCATCTGTGCCACAGCCAATCATCAACATCAAGAGTGAAACGCCCTCT
GAAATCAATGACAAAAAGAGGAAGCGTGAAGAGGACGATGACTATGATGCAGCTGATTGA
Figura 1. Imagen representativa del programa scaffold, en el cual se muestra el gi, la probabilidad de la proteína posible (nombre y organismo al que pertenece), y los aa que se encontraron de la proteína.
Posteriormente, la secuencia de la proteína se metió al programa ExPASy: SIB
Bioinformatics Resource Portal-Home, de libre acceso, el cual proporciona el peso molecular
así como el punto isoeléctrico figura 2.

98
Theoretical pI/Mw (average) for the user-entered sequence: 10 20 30 40 50 60
MAAPKSTPKD AQVMSAILKD MGIVDYEPRV INQMLEFTYR YVTNILEDAR LYSTHSKKRA
70 80 90 100 110 120
VDVDDVRLAI QMQAEKTFTS PPPRDLLMEI ARQKNSTPLP LIKANAGPRL PPDRYSLTAC
130 140 150 160 170 180
NYRLKSAKKP RIQVGLPPQL GLATMQGNSA AGLSKVTPAL SLNSKTGATL AVVSKPASAP
190 200 210 220 230 240
TVTIVSRPAT ANKMHPTPII KLSQRPPLNV STAVSSAAPT PIVQVTAAPQ PTPPAPTTTA
250 260 270 280 290
AVPATEATTT PPVITTATTE PATQPSVPQP IINIKSETPS EINDKKRKRE EDDDYDAAD
Theoretical pI/Mw: 9.56 / 32012.79
Figura 2. Cálculo del peso y punto isoeléctrico de la proteína 1-Int-Insol.
14.6. Anexo 3. Análisis in silico de la proteína de garrapata de 2-intestino
insoluble
14.7. Proteína 2-Int-Insol
Rhipicephalus pulchellus
gi: L7MAA0
Nombre de la proteína, producto: Putative f0f1-type atp synthase alpha subunit
Secuencia aa. de la proteína
MAFVSRRLACTLARHFPKQTPKVTHEAISALALRKLSTTPCSYAAAGSAEVSSILEERILGQTSTTNLEETGRVLSIGDG
IARVYGLKNIQAEEMVEFSSGLKGMALNLEPDNVGIVVFGNDKLIKEGDIVKRTGAIVDVPVGPELLGRVVDALGNPIDG
KGPVACKSRARVGIKAPGIIPRISVKEPMLTGIKAVDSLVPIGRGQRELIIGDRQTGKTAIAIDGIINQKRFNDGSDEKK
KLYCIYVAIGQKRSTVAQIVKRLTQSDAMKYTIIVSATASDAAPLQYLAPYSGCAMGEYFRDNGMHALIIYDDLSKQAVA
YRQMSLLLRRPPGREAYPGDVFYLHSRLLERAAKMNDTFGGGSLTALPVIETQAGDVSAYIPTNVISITDGQIFLETELF
YKGIRPAINVGLSVSRVGSAAQTKAMKQVAGSMKLELAQYREVAAFAQFGSDLDAATQQLLNRGVRLTELLKQGQYTPMA
IEDQVAVIYTGVRGYLDKLDPAKINDFEKEFLQHIRSHHKDVLATIAQEGHISEATDAKLKNIVTSFMANFTA
Secuencia de nucleótidos
ENA|GACK01004976|GACK01004976.1 TSA: Rhipicephalus pulchellus RpMit-677801 mRNA
sequence. : Location:1.1000
ATGGCTTTCGTCTCTCGCAGGTTGGCCTGCACCTTGGCCAGGCACTTCCCGAAGCAAACC
CCGAAAGTTACTCACGAAGCGATAAGTGCGCTCGCACTACGAAAGCTTTCGACCACGCCA
TGCTCCTATGCAGCCGCCGGCTCGGCCGAGGTTTCCAGCATCCTCGAGGAGCGCATTCTG
GGCCAGACCTCTACGACTAACCTCGAGGAAACTGGCCGAGTGCTCAGCATTGGAGACGGT
ATCGCCCGCGTCTATGGCCTCAAGAACATTCAAGCCGAAGAGATGGTGGAGTTTTCCAGC
GGGTTGAAAGGTATGGCCCTTAACTTGGAGCCGGACAACGTGGGTATCGTCGTGTTCGGT
AACGACAAGCTTATTAAGGAGGGCGACATTGTGAAGCGTACCGGAGCCATCGTGGACGTA
CCGGTCGGTCCCGAGCTCCTTGGTCGCGTTGTGGATGCCCTTGGCAATCCCATAGATGGC

99
AAAGGACCCGTGGCTTGCAAGTCGCGGGCTCGTGTCGGCATAAAGGCGCCAGGCATTATC
CCGCGTATCTCAGTAAAGGAGCCTATGCTTACAGGAATCAAAGCAGTCGACAGCTTAGTG
CCTATTGGTCGTGGCCAGCGGGAACTCATTATTGGAGACAGGCAGACCGGCAAAACTGCT
ATCGCCATTGATGGAATTATCAACCAGAAGCGTTTCAATGATGGTTCCGACGAAAAGAAA
AAGCTGTACTGTATTTACGTTGCCATTGGACAGAAGAGAAGCACAGTCGCACAGATCGTA
AAACGATTGACCCAGAGCGATGCCATGAAGTACACCATCATCGTCAGTGCCACTGCATCG
GACGCTGCCCCGCTTCAGTACCTTGCGCCCTATTCGGGATGTGCCATGGGCGAGTATTTC
CGCGACAACGGCATGCACGCCCTCATCATCTACGACGATTTGTCCAAGCAGGCCGTCGCG
TACCGTCAAATGTCTCTGCTGCTGCGACGGCCCCCAGGTC.
Referencia
References: An insight into the sialotranscriptome of the Zebra tick, Rhipicephalus pulchellus
Tan A.A.W.L., Francischetti I.M.B., Manjunatha K.K.R., Ribeiro J.M.C.
Tan A.A.W.L., Francischetti I.M.B., Manjunatha K.K.R., Ribeiro J.M.C.
Submitted (16-NOV-2012) to the INSDC. Laboratory of Malaria and Vector Research,
National Institute of Allergy and Infectious Diseases, 12735 Twinbrook Parkway room
2E32D, Rockville, MD 20852, USA.

100
Figura 3. Imagen representativa del programa scaffold, en el cual se muestra el gi, la probabilidad de la proteína posible (nombre y organismo al que pertenece), y los aa que se encontraron de la proteína.
Posteriormente la secuencia de la proteína se metio al programa ExPASy: SIB Bioinformatics
Resource Portal-Home de libre acceso la cual nos proporcionó su peso molecular asi como
su punto isoeléctrico figura 4.
Theoretical pI/Mw (average) for the user-entered sequence:
10 20 30 40 50 60
MAFVSRRLAC TLARHFPKQT PKVTHEAISA LALRKLSTTP CSYAAAGSAE VSSILEERIL
70 80 90 100 110 120
GQTSTTNLEE TGRVLSIGDG IARVYGLKNI QAEEMVEFSS GLKGMALNLE PDNVGIVVFG
130 140 150 160 170 180
NDKLIKEGDI VKRTGAIVDV PVGPELLGRV VDALGNPIDG KGPVACKSRA RVGIKAPGII
190 200 210 220 230 240
PRISVKEPML TGIKAVDSLV PIGRGQRELI IGDRQTGKTA IAIDGIINQK RFNDGSDEKK
250 260 270 280 290 300
KLYCIYVAIG QKRSTVAQIV KRLTQSDAMK YTIIVSATAS DAAPLQYLAP YSGCAMGEYF
310 320 330 340 350 360
RDNGMHALII YDDLSKQAVA YRQMSLLLRR PPGREAYPGD VFYLHSRLLE RAAKMNDTFG
370 380 390 400 410 420
GGSLTALPVI ETQAGDVSAY IPTNVISITD GQIFLETELF YKGIRPAINV GLSVSRVGSA
430 440 450 460 470 480
AQTKAMKQVA GSMKLELAQY REVAAFAQFG SDLDAATQQL LNRGVRLTEL LKQGQYTPMA
490 500 510 520 530 540
IEDQVAVIYT GVRGYLDKLD PAKINDFEKE FLQHIRSHHK DVLATIAQEG HISEATDAKL

101
550
KNIVTSFMAN FTA
Theoretical pI/Mw: 8.98 / 59905.07
Figura 4. Cálculo del peso y punto isoeléctrico de la proteína 2-Int-Insol.
14.8. Anexo 4. Análisis in silico de la proteína de garrapata de 1-ovario
insoluble
http://www.ebi.ac.uk/ena/data/view/GU994210
14.9. Proteína 1-Ova-Insol
Rhipicephalus microplus Gen: VDAC 273 AA.
gi:E7CF11
Producto: mitochondrial volatge dependent anion-selective cannel
Secuencia aa. de la proteína
MAPPCYADLGKQARDLFNKNYHFGVVKLDCKSTTQTGVEFNVSGTSLNDTGKVNASLETKYKVPEYGLTLKEKWNTDNTL
STEICSEEKLARGLKVAFHANFAPQTGKKSGALKAAYKFDNVHLNGDVDLGPPGPLVHGAAVLHYQGWLAGGQVSFDTTK
NRLSKTNFAVGFQAGDFVLHTNVNDGQEFAGSVFQRVNPQLQTGVQLAWTAGTNATRFGLGCVYDLDMETSVRAKVNNSG
QIGLGFTHRLRPGISLTLSTMLDGKNFNQGGHK
Secuencia de nucleótidos
>ENA|GU994210|GU994210.1 Rhipicephalus microplus mitochondrial volatge dependent
anion-selective channel
(VDAC) gene, partial cds; nuclear gene for mitochondrial product. :
Location:1..819
ATGGCTCCTCCGTGCTACGCAGACTTGGGCAAGCAGGCTCGCGACCTGTTCAACAAGAAC
TACCACTTCGGCGTCGTGAAGCTCGACTGCAAGTCGACCACCCAGACGGGCGTCGAGTTC
AACGTGAGCGGCACCAGCCTCAATGACACCGGCAAGGTGAACGCCTCGCTCGAGACCAAG
TACAAGGTTCCCGAGTACGGGCTCACGCTCAAGGAAAAATGGAACACCGACAACACGCTC
TCGACCGAGATCTGCAGCGAGGAGAAGCTCGCGCGCGGCCTCAAAGTCGCCTTCCACGCC
AACTTCGCACCCCAGACTGGCAAGAAGTCGGGAGCCCTGAAGGCGGCCTACAAGTTCGAC
AACGTGCACCTGAACGGCGACGTCGACCTGGGACCTCCGGGTCCACTCGTGCACGGTGCG
GCCGTGCTCCACTACCAGGGCTGGCTCGCCGGCGGGCAGGTCTCGTTCGACACCACCAAG
AACCGGCTGAGCAAGACAAACTTCGCCGTGGGTTTCCAGGCCGGCGACTTCGTACTGCAC
ACGAACGTCAACGACGGCCAGGAGTTTGCCGGGTCCGTGTTTCAACGGGTGAACCCGCAG
TTGCAGACGGGAGTCCAGCTGGCCTGGACCGCTGGCACGAACGCGACCCGATTCGGCCTG
GGTTGCGTCTACGACCTCGACATGGAGACCTCGGTTCGCGCGAAGGTGAACAACTCTGGT
CAGATCGGGCTCGGGTTCACGCACCGCCTGAGACCGGGCATCAGCCTGACCCTCTCCACC
ATGCTGGACGGCAAGAACTTCAACCAGGGAGGACACAAG
Referencia
The identification of a VDAC-like protein involved in the interaction of Babesia bigemina sexual stages with Rhipicephalus microplus midgut cells.
Rodríguez-Hernández E., Mosqueda J., Alvarez-Sánchez ME., Neri AF., Mendoza-Hernández G., Camacho-Nuez M.
Posgrado en Ciencias Genómicas, Universidad Autónoma de la Ciudad de México, San Lorenzo Núm. 290, esquina Roberto Gayol, Colonia del Valle Sur, delegación Benito Juárez, México DF, CP 03100, Mexico.
Vet. Parasitol. 187(3-4): 538-541 (2012 Jul).

102
Figura 5. Imagen representativa del programa scaffold, en el cual se muestra el gi, la probabilidad de la proteína posible (nombre y organismo al que pertenece), y los aa que se encontraron de la proteína.
Posteriormente la secuencia de la proteína se metio al programa ExPASy: SIB Bioinformatics Resource Portal-Home de libre acceso la cual nos proporcionó su peso molecular asi como su punto isoeléctrico figura 6.
Theoretical pI/Mw (average) for the user-entered sequence:
10 20 30 40 50 60
MAPPCYADLG KQARDLFNKN YHFGVVKLDC KSTTQTGVEF NVSGTSLNDT GKVNASLETK
70 80 90 100 110 120
YKVPEYGLTL KEKWNTDNTL STEICSEEKL ARGLKVAFHA NFAPQTGKKS GALKAAYKFD
130 140 150 160 170 180
NVHLNGDVDL GPPGPLVHGA AVLHYQGWLA GGQVSFDTTK NRLSKTNFAV GFQAGDFVLH
190 200 210 220 230 240
TNVNDGQEFA GSVFQRVNPQ LQTGVQLAWT AGTNATRFGL GCVYDLDMET SVRAKVNNSG
250 260 270
QIGLGFTHRL RPGISLTLST MLDGKNFNQG GHK
Theoretical pI/Mw: 8.94 / 29520.23

103
Figura 6. Cálculo del peso y punto isoeléctrico de la proteína 1-Ova-Insol.
14.10. Anexo 5. Análisis in silico de la proteína de garrapata de 2-ovario
insoluble
http://www.ebi.ac.uk/ena/data/view/AY312432
Proteína 2-Ova-Insol
Productoe: serpin [Rhipicephalus microplus]
gi: 07Z021
Secuencia de aa
>gi|32351460|gb|AAP75707.1| serpin [Rhipicephalus microplus]
MLAKFLFLASALAVAHCDTDDSTLLARAHNQFAVNLLKQLATENPSSNVFFSPTSIAAAFGMAYLGARGG
SESELNSVFGHADVGLTDRSRLLTAYKNLLELSASPNVTLDVANMVLAQDRFPISDSYKQQLREIFDADL
RSANFVEDGPRVAAEVNAWVRERQGARSRYPPEGQPLDIVLFILNAVYFKGTWVTFDAHRTINKPSSPGT
TEVSKPAMHLKARFPYARVEPLHASALEIPYEGDRFTMVVLLPDNATGLAAVRNGLSLAALEDVGSRLSF
RDVILQLPKFDMSLSYGLVPAMKAIGLNSVFGGSADFSGISEAVPLVISDVLHKAAVEVNEEGTIATAVT
GLGFVPLSVHYNPPPPIEFTVDHPFIFYIRDRSTNRVLFIGEVNTL
Secuencia e nucleótidos
ATGCTGGCGAAATTTCTGTTTCTGGCGAGCGCGCTGGCGGTGGCGCATTGCGATACCGAT
GATAGCACCCTGCTGGCGCGCGCGCATAACCAGTTTGCGGTGAACCTGCTGAAACAGCTG
GCGACCGAAAACCCGAGCAGCAACGTGTTTTTTAGCCCGACCAGCATTGCGGCGGCGTTT
GGCATGGCGTATCTGGGCGCGCGCGGCGGCAGCGAAAGCGAACTGAACAGCGTGTTTGGC
CATGCGGATGTGGGCCTGACCGATCGCAGCCGCCTGCTGACCGCGTATAAAAACCTGCTG
GAACTGAGCGCGAGCCCGAACGTGACCCTGGATGTGGCGAACATGGTGCTGGCGCAGGAT
CGCTTTCCGATTAGCGATAGCTATAAACAGCAGCTGCGCGAAATTTTTGATGCGGATCTG
CGCAGCGCGAACTTTGTGGAAGATGGCCCGCGCGTGGCGGCGGAAGTGAACGCGTGGGTG
CGCGAACGCCAGGGCGCGCGCAGCCGCTATCCGCCGGAAGGCCAGCCGCTGGATATTGTG
CTGTTTATTCTGAACGCGGTGTATTTTAAAGGCACCTGGGTGACCTTTGATGCGCATCGC
ACCATTAACAAACCGAGCAGCCCGGGCACCACCGAAGTGAGCAAACCGGCGATGCATCTG
AAAGCGCGCTTTCCGTATGCGCGCGTGGAACCGCTGCATGCGAGCGCGCTGGAAATTCCG
TATGAAGGCGATCGCTTTACCATGGTGGTGCTGCTGCCGGATAACGCGACCGGCCTGGCG
GCGGTGCGCAACGGCCTGAGCCTGGCGGCGCTGGAAGATGTGGGCAGCCGCCTGAGCTTT
CGCGATGTGATTCTGCAGCTGCCGAAATTTGATATGAGCCTGAGCTATGGCCTGGTGCCG
GCGATGAAAGCGATTGGCCTGAACAGCGTGTTTGGCGGCAGCGCGGATTTTAGCGGCATT
AGCGAAGCGGTGCCGCTGGTGATTAGCGATGTGCTGCATAAAGCGGCGGTGGAAGTGAAC
GAAGAAGGCACCATTGCGACCGCGGTGACCGGCCTGGGCTTTGTGCCGCTGAGCGTGCAT
TATAACCCGCCGCCGCCGATTGAATTTACCGTGGATCATCCGTTTATTTTTTATATTCGC
GATCGCAGCACCAACCGCGTGCTGTTTATTGGCGAAGTGAACACCCTG
Referencia
"Cloning and partial characterization of a serine proteinase inhibitor (serpin) from Boophilus
microplus."
Zhou J., Zhou Y., Gong H., Chen L.
Submitted (JUN-2003) to the EMBL/GenBank/DDBJ databases

104
Cited for: NUCLEOTIDE SEQUENCE.
Figura 7. Imagen representativa del programa scaffold, en el cual se muestra el gi, la probabilidad de la proteína posible (nombre y organismo al que pertenece), y los aa que se encontraron de la proteína.
Posteriormente la secuencia de la proteína se metio al programa ExPASy: SIB Bioinformatics
Resource Portal-Home de libre acceso la cual nos proporcionó su peso molecular asi como su
punto isoeléctrico figura 8.
Theoretical pI/Mw (average) for the user-entered sequence:
10 20 30 40 50 60
MLAKFLFLAS ALAVAHCDTD DSTLLARAHN QFAVNLLKQL ATENPSSNVF FSPTSIAAAF
70 80 90 100 110 120
GMAYLGARGG SESELNSVFG HADVGLTDRS RLLTAYKNLL ELSASPNVTL DVANMVLAQD
130 140 150 160 170 180
RFPISDSYKQ QLREIFDADL RSANFVEDGP RVAAEVNAWV RERQGARSRY PPEGQPLDIV

105
190 200 210 220 230 240
LFILNAVYFK GTWVTFDAHR TINKPSSPGT TEVSKPAMHL KARFPYARVE PLHASALEIP
250 260 270 280 290 300
YEGDRFTMVV LLPDNATGLA AVRNGLSLAA LEDVGSRLSF RDVILQLPKF DMSLSYGLVP
310 320 330 340 350 360
AMKAIGLNSV FGGSADFSGI SEAVPLVISD VLHKAAVEVN EEGTIATAVT GLGFVPLSVH
370 380 390
YNPPPPIEFT VDHPFIFYIR DRSTNRVLFI GEVNTL
Theoretical pI/Mw: 5.51 / 43043.08 Figura 8. Cálculo del peso y punto isoeléctrico de la proteína 2-Ova-Insol.
14.11. Anexo 6. Análisis in silico de la proteína de garrapata de 3-ovario
insoluble
http://www.ebi.ac.uk/ena/data/view/AY519366
3-0va-insol
Organismo: Rhipicephalus microplus
gi: Q6R128
Product o: beta-actina
Secuencia de aa.
SSSLEKSYELPDGQVITIGNERFRCPEALFQPSFLGMEACGIHETTYNSIMKCDVDIRKDLYANTVLSGGTTMYPGIADR
MQKEITALAPSTMKIKIIAPPERKYSVWIGG
Secuencia de nucleótidos
ENA|AY519366|AY519366.1 Boophilus microplus clone 1 beta-actin mRNA, partial cds.
: Location:1.332
TCCTCGTCCCTGGAGAAGTCGTACGAGCTTCCCGACGGTCAGGTCATCACGATCGGCAAC
GAGCGGTTCCGTTGCCCCGAGGCCCTGTTCCAGCCCTCGTTCCTGGGTATGGAAGCGTGC
GGCATCCACGAGACCACCTACAACTCCATCATGAAGTGCGACGTGGACATCAGGAAGGAC
CTGTACGCAAACACCGTCCTCTCCGGCGGCACCACCATGTACCCAGGCATCGCCGACCGC
ATGCAGAAGGAAATCACCGCGCTTGCGCCGTCCACCATGAAGATCAAGATCATCGCGCCC
CCCGAGAGGAAGTACTCGGTCTGGATCGGTGG
Referencias
1 Two partial beta-actin clones from the southern cattle tick, Boophilus microplus Holmes S.P., Barron
P.D., He H., Pietrantonio P.V.
2 Holmes S.P., Barron P.D., He H., Pietrantonio P.V.
Submitted (07-JAN-2004) to the INSDC. Entomology, Texas A&M University, Heep Ctr Rm 412, College
Station, TX 77843, USA

106
Figura 9. Imagen representativa del programa scaffold, en el cual se muestra el gi, la probabilidad de la proteína posible (nombre y organismo al que pertenece), y los aa que se encontraron de la proteína.
Posteriormente la secuencia de la proteína se metio al programa ExPASy: SIB Bioinformatics Resource Portal-Home de libre acceso la cual nos proporcionó su peso molecular asi como su punto isoeléctrico figura 10.
Theoretical pI/Mw (average) for the user-entered sequence:
10 20 30 40 50 60
MCDDEVAALV VDNGSGMCKA GFAGDDAPRA VFPSIVGRPR HQGVMVGMGQ KDSYVGDEAQ
70 80 90 100 110 120
SKRGILTLKY PIEHGIVTNW DDMEKIWHHT FYNELRVAPE EHPVLLTEAP LNPKANREKM
130 140 150 160 170 180
TQIMFETFNT PAMYVAIQAV LSLYASGRTT GIVLDSGDGV SHTVPIYEGY ALPHAILRLD
190 200 210 220 230 240
LAGRDLTDYL MKILTERGYS FTTTAEREIV RDIKEKLCYV ALDFEQEMAT AASSSSLEKS
250 260 270 280 290 300
YELPDGQVIT IGNERFRCPE ALFQPSFLGM ESCGIHETTY NSIMKCDVDI RKDLYANTVL

107
310 320 330 340 350 360
SGGTTMYPGI ADRMQKEITA LAPSTMKIKI IAPPERKYSV WIGGSILASL STFQQMWISK
370
QEYDESGPSI VHRKCF
Theoretical pI/Mw: 5.30 / 41823.81
Figura 10. Cálculo del peso y punto isoeléctrico de la proteína 3-Ova-Insol.
14.12. Anexo 7. Análisis in silico de la proteína de garrapata de 4-ovario
insoluble
http://www.ebi.ac.uk/ena/data/view/GACK01008723
4-Ova-Insol
Organismo: Rhipicephalus pulchellus
gi: L7LW49
Producto: hypothetical protein
Secuencia de aa
MSAQNQPDPLFDIRNAFYIGNYQYCITEAQKGKNVSPEHKLEKDVFLYRAYIVQRKYGVVLDEIRSMAPDELQCIRMLAD
YLSGDMARKDQVVKQLDQKLSKSLDVNNVILPVVAATIYYHEQNYEAALRVLHQNDSLECAALTLQCYLKLDRLDLARKE
LKRMQEKDDDATLTQLAQAWVNLYLGGEKLQEAFYIYQELAEKNTATPLLLNGQATAYIAQGKYEEAEALLQEAIEKDSN
HVETLINLVVLSQHLGKSPEVTSRLLSQLRDTNNSHPFVKEYQSKEQEFDRLARQYAPA
Secuencia de nucleótidos
>ENA|GACK01008723|GACK01008723.1 TSA: Rhipicephalus pulchellus RpIx75-842552 mRNA
sequence. : Location:1.900
ATGTCAGCTCAGAATCAGCCTGATCCCCTTTTTGATATCCGAAATGCCTTCTACATCGGA
AACTACCAGTACTGCATCACCGAAGCTCAGAAAGGAAAGAACGTCAGTCCTGAGCACAAG
CTGGAGAAAGACGTATTCCTGTACAGAGCTTACATAGTGCAACGGAAGTACGGCGTCGTC
TTGGACGAAATTCGTTCGATGGCTCCCGACGAACTGCAGTGCATCCGTATGCTCGCCGAT
TACCTCTCCGGGGACATGGCTCGAAAGGACCAAGTTGTGAAGCAGCTTGACCAGAAGCTA
TCCAAGAGCTTGGACGTCAACAATGTGATTCTTCCAGTTGTGGCAGCCACCATCTACTAC
CATGAACAGAACTACGAAGCTGCTCTGCGAGTCCTTCACCAGAACGACTCCTTGGAGTGT
GCTGCACTTACCCTACAATGCTATCTTAAACTGGACAGACTTGACTTGGCTAGGAAAGAA
CTGAAAAGAATGCAAGAGAAGGATGATGATGCGACGTTAACGCAACTTGCACAAGCCTGG
GTCAACCTTTATCTGGGTGGAGAAAAGCTGCAGGAGGCCTTCTACATCTATCAGGAGCTG
GCTGAGAAGAACACGGCTACTCCACTGCTGCTGAATGGACAGGCAACAGCCTATATTGCT
CAGGGAAAATATGAAGAAGCAGAGGCTCTGCTGCAGGAAGCAATTGAAAAGGATAGCAAC
CATGTGGAAACGCTCATCAACCTGGTGGTGCTGTCTCAGCATTTGGGAAAATCTCCAGAG
GTGACAAGCCGTCTATTAAGCCAACTCCGTGATACGAACAACAGTCATCCATTCGTGAAG
GAATACCAGAGCAAAGAGCAGGAATTTGACCGTCTGGCGAGGCAATATGCACCTGCTTAA
Referencia
An insight into the sialotranscriptome of the Zebra tick, Rhipicephalus pulchellus Tan
A.A.W.L., Francischetti I.M.B., Manjunatha K.K.R., Ribeiro J.M.C.

108
[2]Tan A.A.W.L., Francischetti I.M.B., Manjunatha K.K.R., Ribeiro J.M.C. Submitted (16-NOV-
2012) to the INSDC. Laboratory of Malaria and Vector Research, National Institute of Allergy
and Infectious Diseases, 12735 Twinbrook Parkway room 2E32D, Rockville, MD 20852, USA
Figura 11. Imagen representativa del programa scaffold, en el cual se muestra el gi, la probabilidad de la proteína posible (nombre y organismo al que pertenece), y los aa que se encontraron de la proteína.
Posteriormente la secuencia de la proteína se metio al programa ExPASy: SIB Bioinformatics
Resource Portal-Home de libre acceso la cual nos proporcionó su peso molecular asi como su
punto isoeléctrico figura 12.
Theoretical pI/Mw (average) for the user-entered sequence:
10 20 30 40 50 60
MSAQNQPDPL FDIRNAFYIG NYQYCITEAQ KGKNVSPEHK LEKDVFLYRA YIVQRKYGVV
70 80 90 100 110 120
LDEIRSMAPD ELQCIRMLAD YLSGDMARKD QVVKQLDQKL SKSLDVNNVI LPVVAATIYY
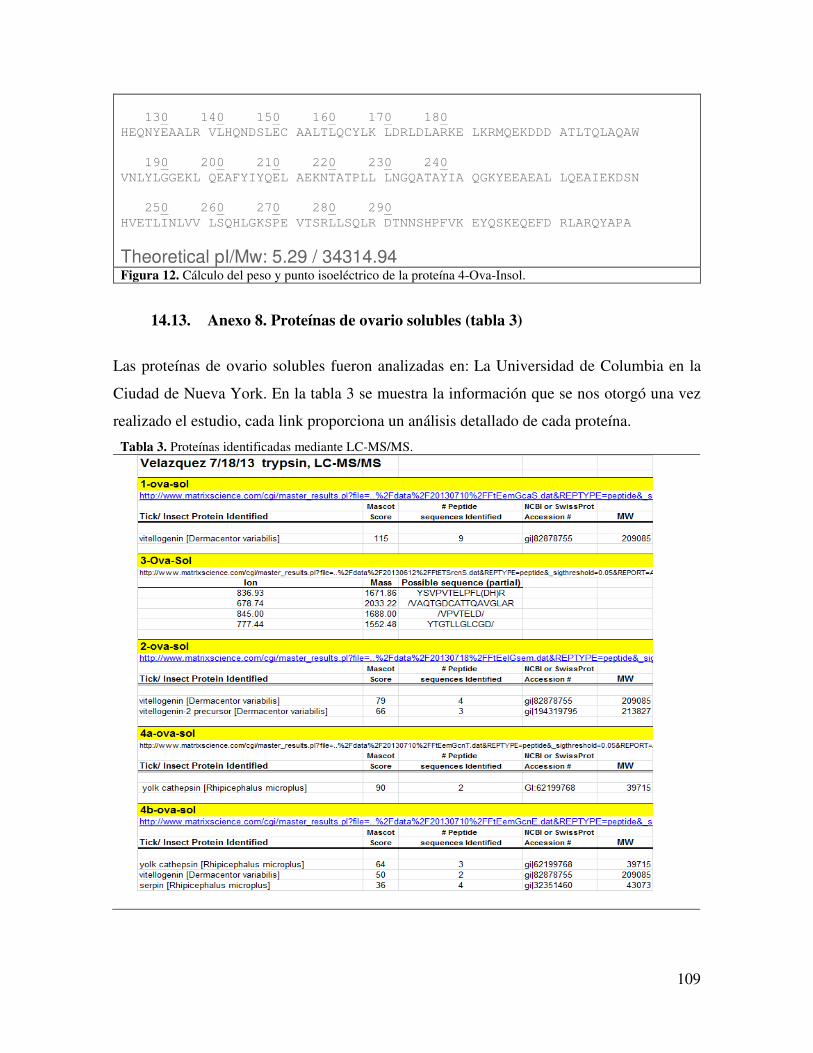
109
130 140 150 160 170 180
HEQNYEAALR VLHQNDSLEC AALTLQCYLK LDRLDLARKE LKRMQEKDDD ATLTQLAQAW
190 200 210 220 230 240
VNLYLGGEKL QEAFYIYQEL AEKNTATPLL LNGQATAYIA QGKYEEAEAL LQEAIEKDSN
250 260 270 280 290
HVETLINLVV LSQHLGKSPE VTSRLLSQLR DTNNSHPFVK EYQSKEQEFD RLARQYAPA
Theoretical pI/Mw: 5.29 / 34314.94 Figura 12. Cálculo del peso y punto isoeléctrico de la proteína 4-Ova-Insol.
14.13. Anexo 8. Proteínas de ovario solubles (tabla 3)
Las proteínas de ovario solubles fueron analizadas en: La Universidad de Columbia en la
Ciudad de Nueva York. En la tabla 3 se muestra la información que se nos otorgó una vez
realizado el estudio, cada link proporciona un análisis detallado de cada proteína.
Tabla 3. Proteínas identificadas mediante LC-MS/MS.

Full length article
Identification of immunogenic proteins from ovarian tissue andrecognized in larval extracts of Rhipicephalus (Boophilus) microplus,through an immunoproteomic approach
Patricia Berenice Ramírez Rodríguez a, Rodrigo Rosario Cruz b,Delia In�es Domínguez García b, Rodolfo Hern�andez Guti�errez a,Rodolfo Esteban Lagunes Quintanilla c, Daniel Ortu~no Sahagún d, Celia Gonz�alez Castillo d,Abel Guti�errez Ortega a, Sara Elisa Herrera Rodríguez a, Adriana Vallejo Cardona a,Mois�es Martínez Vel�azquez a, *
a Centro de Investigaci�on y Asistencia en Tecnología y Dise~no del Estado de Jalisco, AC. Av. Normalistas 800, Col. Colinas de la Normal, C.P. 44270,
Guadalajara, Jalisco, Mexicob Unidad de Investigaci�on en Biotecnología, Salud y Ambiente (BIOSA-UAGro) de la Unidad Acad�emica de Ciencias Naturales de la Universidad Aut�onoma de
Guerrero, Carretera Federal Chilpancingo-Acapulco, Ex-Rancho el “Shalako”, Petaquillas, Guerrero, C.P. 39105, Mexicoc Centro Nacional de Investigaciones en Parasitología Veterinaria-INIFAP, Carretera Federal Cuernavaca Cuautla No. 8534, Col. Progreso, C.P. 62550, Jiutepec,
Morelos, Mexicod Instituto de Investigaci�on en Ciencias Biom�edicas (IICB), CUCS, Universidad de Guadalajara, Sierra Mojada No. 950, Col. Independencia, C.P. 44340,
Guadalajara, Jalisco, Mexico
h i g h l i g h t s g r a p h i c a l a b s t r a c t
� A classical approach immunizing
cattle with total ovarian proteins was
followed.
� The detection of ovarian proteins was
achieved by ELISA plates coated with
proteins from larval homogenates.
� Several immunogenic proteins such
as Vitellogenins and yolk Cathepsin
were identified.
� Vitellogenin-1 and Vitellogenin-2
transcripts are synthesized in the
ovaries of R. microplus.
� Vitellogenins represent attractive
candidates for development of anti-
tick vaccines.
a r t i c l e i n f o
Article history:
Received 21 April 2016
Received in revised form
3 October 2016
Accepted 6 October 2016
Available online 8 October 2016
a b s t r a c t
Rhipicephalus (Boophilus) microplus ticks are obligatory hematophagous ectoparasites of cattle and act as
vectors for disease-causing microorganisms. Conventional tick control is based on the use of chemical
acaricides; however, their uncontrolled use has increased tSresistant tick populations, as well as food and
environmental contamination. Alternative immunological tick control has shown to be partially effective.
The only anti-tick vaccine commercially available at present in the world is based on intestinal Bm86
* Corresponding author. Av. Normalistas 800, Col. Colinas de la Normal, C.P. 44270, Guadalajara, Jalisco, Mexico.
E-mail addresses: [email protected] (P.B. Ramírez Rodríguez), [email protected] (R. Rosario Cruz), [email protected] (D.I. Domínguez García),
[email protected] (R. Hern�andez Guti�errez), [email protected] (R.E. Lagunes Quintanilla), [email protected] (D. Ortu~no Sahagún), celia_glz@
hotmail.com (C. Gonz�alez Castillo), [email protected] (A. Guti�errez Ortega), [email protected] (S.E. Herrera Rodríguez), [email protected] (A. Vallejo Cardona),
[email protected] (M. Martínez Vel�azquez).
Contents lists available at ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/locate /yexpr
http://dx.doi.org/10.1016/j.exppara.2016.10.005
0014-4894/© 2016 Elsevier Inc. All rights reserved.
Experimental Parasitology 170 (2016) 227e235

Keywords:
Tick
Boophilus microplus
Ovary
Immunogen
Vitellogenin
Vaccine
protein, and shows a variable effectiveness depending on tick strains or geographic isolates. Therefore,
there is a need to characterize new antigens in order to improve immunological protection. The aim of
this work was to identify immunogenic proteins from ovarian tissue extracts of R. microplus, after cattle
immunization. Results showed that ovarian proteins complexed with the adjuvant Montanide ISA 50 V
generated a strong humoral response on vaccinated cattle. IgG levels peaked at fourth post-
immunization week and remained high until the end of the experiment. 1D and 2D SDS-PAGE-
Western blot assays with sera from immunized cattle recognized several ovarian proteins. Reactive
bands were cut and analyzed by LC-MS/MS. They were identified as Vitellogenin, Vitellogenin-2 pre-
cursor and Yolk Cathepsin. Our findings along with bioinformatic analysis indicate that R. microplus has
several Vitellogenin members, which are proteolytically processed to generate multiple polypeptide
fragments. This apparent complexity of vitellogenic tick molecular targets gives the opportunity to
explore their potential usefulness as vaccine candidates but, at the same time, imposes a challenge on the
selection of the appropriate set of antigens.
© 2016 Elsevier Inc. All rights reserved.
1. Introduction
Rhipicephalus (Boophilus) microplus ticks are obligatory hema-
tophagous ectoparasites of cattle and act as vectors for disease-
causing microorganisms such as protozoa of the genus Babesia
spp. and bacteria Anaplasma spp. (P�erez de Le�on et al., 2010).
Together, these microorganisms and ticks may induce anemia and
weight loss and decrease milk and meat production in the para-
sitized cattle, as well as causing the depreciation of cattle skin value
in themarket. The conventional R. microplus tick control is based on
the use of chemical acaricides (Martins et al., 2002). However,
uncontrolled use of these agents has increased the incidence of tick
populations resistant to acaricides (Crampton et al., 1999; Martins
and Furlong, 2001; Klafke et al., 2006; Parizi et al., 2009). Addi-
tionally, chemical treatments have severe limitations due to the
food safety and environmental pollution problems they cause. The
alternative immunological control of tick infestations has shown to
be partially effective. Proven vaccines such as Gavac™ and Tick-
Gard™ are based on the recombinant Bm86 antigen, a R. microplus
midgut cell membrane protein (Rand et al., 1989; Willadsen et al.,
1989; Gough and Kemp, 1993). These vaccines are used to immu-
nize cattle in order to induce immunoglobulins that have the ca-
pacity to bind the tick's intestinal cells after blood feeding, and
subsequently cause the ectoparasite's death (Maritz-Olivier et al.,
2012; Rodriguez-Valle et al., 2012).
Currently, the only anti-tick vaccine commercially available in
the world is Gavac™, based on Bm86. Its high protection, durability
and environmentally friendly characteristics have made it widely
used (Rodríguez, 2000). Nevertheless, its effectiveness varies
depending on tick strains or geographic isolates due to the varia-
tions in Bm86 sequence between different tick populations (García-
García et al., 2000). Consequently, research efforts are still being
made worldwide to identify new candidate antigens for a more
effective vaccine to control tick infestations (de la Fuente et al.,
2007). The development of anti-tick vaccines converge in the
identification, molecular cloning and in vitro production of proteins
that play important roles in tick physiology, such as cell signaling,
modulation of host immune response, pathogen transmission,
embryogenesis, digestion, and intermediary metabolism (Parizi
et al., 2009). It is known that a significant portion of immuno-
globulins that are taken up with blood meal retain biological ac-
tivity during their passage through the tick's gut, hemolymph and
salivary glands. Therefore, the potential targets for a protective
immune response could be any of the antigens in the internal or-
gans that may be in contact with host antibodies, including anti-
gens located in ovaries (Wang and Nuttall, 1999). In this context, the
aim of this work was to identify immunogenic proteins from
ovarian tissue of R. microplus utilizing an immunoproteomic
approach. Additionally, it was hypothesized that sera from immu-
nized cattle can recognize proteins present in both larvae and
ovaries. Antigens reported here could be potentially used as vaccine
targets, which could result in a detrimental effect on oogenesis,
embryogenesis and reproductive rates of R. microplus and, there-
fore, are a promising option for addressing the tick population
control.
2. Material and methods
2.1. Ticks
Engorged adult female ticks of R. micropluswere obtained from a
susceptible strain reared under laboratory conditions at CENID-
PAVET-INIFAP, located in Jiutepec, Morelos, Mexico.
2.2. Tissue dissection and protein extraction
A lysate of total proteins wasmade from dissected ovarian tissue
of 15e20 engorged female ticks, collected the following day after
the spontaneous detachment from calves (approximately 21 days
after larvae infestation), following the protocol described by
Rachinsky et al (Rachinsky et al., 2007). with modifications. 100 mg
of ovarian tissue were weighed and placed in an Eppendorf tube
and 1 ml of PBS was added. Tissue was disrupted with a Teflon®
conical pestle and then suspension was passed through a syringe
(barrel size 1 ml; needle size 29 Gauge x½”) 10 times, to help tissue
fragmentation, until it was fully disintegrated. Suspension was
incubated at 70 �C for 5 min and immediately after, at 26 �C for
5 min; this was repeated three times. Next, lysate was sonicated for
5 min and centrifuged at 13,000 rpm for 15 min at 4 �C, then the
supernatant was separated. Protein samples were quantified by the
DC protein assay (Bio-Rad, USA) (Lowry et al., 1951). The absorbance
was read at 750 nm by a Microplate Spectrophotometer (XMark™,
Bio-Rad, USA).
2.3. One-dimensional electrophoresis
Analysis of the ovarian proteins was carried out using one-
dimensional (1D) electrophoresis gels to verify patterns. Briefly,
polyacrylamide gels 7 cm wide x 8 cm long and 1 mm thick (12%)
were made. SDS-PAGE (sodium dodecyl sulfate-polyacrylamide gel
electrophoresis) was conducted with Mini-PROTEAN® Tetra Elec-
trophoresis System (Bio-Rad). 10 mg of ovarian proteins were dis-
solved in SDS-PAGE sample buffer and heated at 100 �C for 5 min.
Molecular weight markers (Precision Plus Protein Standards, Bio-
P.B. Ramírez Rodríguez et al. / Experimental Parasitology 170 (2016) 227e235228

Rad) were diluted 1:19 in SDS-PAGE sample buffer and heated at
100 �C for 5 min. Gels were run at 120 V for 80 min in electro-
phoresis buffer (192 mM glycine, 25 mM Tris, 0.1% SDS), stained
with Coomassie blue and visualized/photographed using a Gel-Doc
XR Documentation System (Bio-Rad).
2.4. Bovines
Six animals aged six months, European crossbred with average
weight of 140 kgwere used. They were kept in experimental animal
facilities at CENID-PAVET-INIFAP. The animals were individually
housed in isolation pens and were fed on fodder receiving water ad
libitum. Animals were randomly assigned to two experimental
groups. The first group (four animals) was immunized with total
ovarian proteins, whereas the second group (two animals) only
with PBS plus adjuvant as a control.
2.5. Immunization
The experimental protocol was approved by the Institutional
Committee of Animal Experimentation of CENID-PAVET-INIFAP.
Cattle were injected subcutaneously in the neck with 2 ml/dose
(100 mg of total ovarian proteins) using a 5 ml syringe and an 18G
needle. Emulsion was made with ovarian protein lysate in PBS plus
adjuvant Montanide ISA 50 V (anhydromannitolether-
octodecenoate in mineral oil) (Seppic, Paris, France) at 1:1 vol ratio.
Two immunizations were applied; the first immunization on the
first day and the booster immunization after 15 days (Fig. 1). All the
animals were checked regularly for any signs of local reaction or
clinical abnormalities post immunization and received treatment in
accordance with the animal experimentation rules described in the
Guide for Care and Use of Laboratory Animals for the CENID-PAVET-
INIFAP.
2.6. Blood collection
Blood samples were collected from cattle every week for 10
weeks. Three samples of 7 ml blood were collected into sterile
tubes using vacutainer syringe needles (0.8 � 38 mm) from caudal
vein. Samples were maintained at 4 �C until arrival at the labora-
tory. Serum was separated from cellular components by centrifu-
gation at 5000 rpm for 1 min before being stored at � 20 �C.
Antibody titers were determined using an antigen-specific indirect
enzyme-linked immunosorbent assay (ELISA).
2.7. ELISA test
Total protein lysate from larvae was prepared according to the
method of Shahein et al. (Shahein et al., 2008). Briefly, one gram of
6-day-old larvae was homogenized in ice-cold phosphate-buff-
ered saline (PBS), filtered, sonicated, and centrifuged at 15,000�g
for 20 min at 4 �C. Supernatant was separated and protein con-
centration quantified by the DC protein assay (Bio-Rad) as
mentioned above. 1 mg of total proteins was diluted in 100 ml
carbonate buffer (50 mM Na2CO3) and used to coat a 96 well
microplate, and then incubated overnight at 4 �C. The plate was
washed four times with 300 ml washing buffer (PBS/0.5% Tween
20), then blocked with blocking buffer (PBS/0.5% Tween 20 and 5%
low fat milk) during one hour at room temperature (25 �C). Next,
the incubation plate was washed with washing buffer as
mentioned above. Immunized and non-immunized calves’ sera
were added by triplicate into wells at a 1:100 dilution ratio. The
plate was washed as described above. A secondary antibody (anti-
bovine IgG-Phosphatase-Coupled) was used at a 1:2000 dilution
ratio. The plate was once more washed and then 200 ml of sub-
strate paranitrophenyl phosphate (PNN) (Sigma 104) was added.
The plate was incubated for 30 min and read at 405 nm in a
Fig. 1. Antibody response in immunized cattle. IgG levels were determined by ELISA in sera from cattle immunized with the ovarian proteins or adjuvant/saline control. Antibody
titers are expressed as the O.D. 405 nm value for the serum dilution 1:100. Arrows indicate the immunization times.
P.B. Ramírez Rodríguez et al. / Experimental Parasitology 170 (2016) 227e235 229

Microplate Spectrophotometer (XMark™, Bio-Rad, USA).
2.8. Two-dimensional electrophoresis
For 2D electrophoresis, immobiline dry-strips gel with linear pH
3e10 and 7 cm in length (Bio-Rad) were rehydrated in gel overnight
with 12 mg of ovary proteins in 125 ml of rehydration buffer (7 M
urea, 2 M thiourea, CHAPS), 1 M DTT, and 20 mM IPG Buffer (3e10).
The first dimensional separation was performed using an Ettan
IPGphor™ 3 Isoelectrofocusing (IEF) system (GE Healthcare); the
programwas run with four steps: Step 1: (Step) 300 V/2:30 h; Step
2: (Gradient) 1000 V/0:30 h; Step 3: (Gradient) 5000 V/1:30 h, and
Step 4: (Step) 5000 V/0:35 h. After IEF running, the immobiline dry-
strips gel containing the focused ovary proteins was equilibrated
following two incubation steps: with 1% DTT (w/v), and with 2.5%
iodoacetamide (w/v) for 20 min, respectively. The immobiline dry-
strips gel was then transferred onto a 12% polyacrylamide slab gel
and 2D separation was carried out under the same running con-
ditions as mentioned above for one-dimensional electrophoresis.
Separated proteins were visualized by Coomassie blue staining.
Two-dimensional electrophoresis images were captured using the
Gel-Doc XR Documentation System (Bio-Rad) and analyzed by
Sigma Gel Software in order to calculate the molecular weight of
the proteins. 2D electrophoresis was performed in quintuplicate by
sample.
2.9. Western blot assays
1D and 2D immunoblots were performed according to the
protocol developed by Towbin et al (Towbin et al., 1979). and
Hawkes et al (Hawkes et al., 1982). with some modifications. In
brief, total ovary proteins were separated in 12% SDS-PAGE gels and
transferred to nitrocellulose membranes using a Mini Trans-Blot
apparatus (Bio-Rad) in transfer buffer (25 mM Tris, 192 mM
glycine, 0.1% SDS, 20% methanol) at 110 V for 60 min. After blotting,
the membrane was blocked in Tris-buffered saline (TBS) containing
0.1% (v/v) Tween 20 (TBS-T) and 5% low fat milk, for 4 h at room
temperature. The blot was briefly washed in TBS-T, then incubated
with sera from control, pre-immune or immunized animals in a
1:50 dilution in TBS-T overnight at 4 �C. Following the incubation,
the membrane was washed 5� 5 minwith TBS-T and incubated for
4 h at room temperature with a 1:2000 dilution of goat anti-bovine
IgG coupled to horseradish peroxidase (Jackson ImmunoResearch
Laboratories, Inc., PA, USA). The membrane was then washed three
times with TBS-T. Detection was carried out using the reagent 4-
chloro-1-naphthol (4CN, Bio-Rad).
2.10. LC-MS/MS analysis
The proteins recognized by bovine's sera in nitrocellulose
membrane were obtained from the corresponding replicate gel and
analyzed by LC-MS/MS. Gel spots were transferred to pre-digested
tubes and 100 ml 0.01 M DTT/0.1 M Tris, pH 8.5 was added. The tube
was placed in a heating block at 55 �C for 1e2 h. After cooling the
tube to room temperature, the liquid was removed and replaced
with 100 ml 0.03 M iodoacetamide/0.1 M Tris, pH 8.5. This was
allowed to react for 30 min in the dark after which the liquid was
removed and the gel was washed as described below.
Gel pieces were prepared for digestion by washing twice with
200 ml 0.05 M Tris, pH 8.5/30% acetonitrile for 20 minwith shaking,
and once with 100 ml acetonitrile for several minutes until the gel
was opaque white. After removing the acetonitrile, the gel pieces
were dried for 20e30 min in a Speed-Vac concentrator.
The gel pieces were digested by adding 0.10 mg modified trypsin
(sequencing grade, RocheMolecular Biochemicals, Indianapolis, IN)
in 50 ml 0.025 M Tris, pH 8.5, or enough volume to completely
hydrate the gel. The tubes were placed in a heating block at 32 �C
and left overnight.
Peptides were extracted with 2 � 50 ml 50% acetonitrile/2% TFA
and the combined extracts were reduced in volume in a Speed-Vac
concentrator and brought up to 20 ml with 0.1% formic acid. 5 ml
aliquots were analyzed by LC-MS/MS.
LC-MS/MS analysis was done on a Waters Q-Tof Ultima hybrid
quadrupole/time-of-flight mass spectrometer with a nano-
electrospray source. The eluant was introduced into the source
through a 15 mm i. d. New Objective PicoTip. Capillary voltage was
set at 1.8 kV and cone voltage 32 V; collision energy was set ac-
cording to mass and charge of the ion, from 14eV to 50eV. Chro-
matography was performed on an LC Packings HPLC with a C18
PepMap column using a linear acetonitrile gradient with flow rate
of 200 nl/min.
Raw data files were processed using the MassLynx ProteinLynx
v.4.1 software and. pkl files were submitted for searching at www.
matrixscience.com using the Mascot algorithm. De novo
sequencing was accomplished by manual inspection of either the
raw MS/MS spectra or spectra processed with the MaxEnt 3 algo-
rithm contained in the Waters MassLynx 4.1 software package.
2.11. RNA isolation and cDNA synthesis
100 mg of ovarian tissue were dissected, weighed and placed
into an Eppendorf tube and 1 ml of TRIZOL reagent (Invitrogen,
Carlsbad, CA, USA) was added to extract total RNA. cDNA was
synthesized from 5 mg of RNA using the Superscript® III First-Strand
Synthesis System (Invitrogen, Carlsbad, CA, USA) according to
manufacturer's instructions.
2.12. Endpoint polymerase chain reaction
R. microplus Vitellogenin 1, Vitellogenin 2 and Vitellogenin 3
encoding sequences were obtained from the GenBank genetic
sequence database (GBBR01000071.1, GBBR01000108.1 (Vg-3 in
this study) and GBBR01000106.1 (Vg-2 in this study), respectively).
The following primers (Integrated DNA Technologies, Coralville, IA,
USA) were used: F-RmVg1 50-CCA ACT CGC TCA AGA CCT TCe30 and
R-RmVg1 50-AAA CCA GGG GTT CCT TGC AC-3’; F-RmVg2(3) 50-CTA
CCTAAG GCG CGAGAT TGe30 and R-RmVg2(3) 50-TGGATT CCGAGA
TGT CCT TCe3’; F-RmVg3(2) 50-AGC TTC GAC AAG ACC CTC AA-30
and R-RmVg3(2) 50-GGT GCT GGT GGT GTA GAG GTe 30, using cDNA
from ovaries as template. PCR conditions were 94 �C for 5 min
followed by 35 cycles, each consisting of denaturation at 94 �C for
30 s, annealing at 55 �C for 30 s, and extension at 72 �C for 60 s. A
final extension step at 72 �C for 7 min was included. The amplified
products were run on 1% agarose gels and analyzed on a UV
transilluminator (BioRad Laboratories, Philadelphia, PA, USA).
Control amplification was carried out using the following primers:
F-50- CCC ATC TAC GAA GGT TAC GCC-30 and R-50-CGC ACG ATT TCA
CGC TCAGe30, designed from R. microplus actin (GenBank accession
number AY255624.1). PCR conditions were the same as mentioned
above.
2.13. Bioinformatic analysis
All primers were designed with the public Primer3 software
v.4.0.0 (http://primer3.wi.mit.edu/ (Rozen and Skaletsky, 2000);).
Phylogenetic analysis was performed using the CLC Main Work-
bench 6.0 software (CLC bio, Cambridge, MA, USA), with 1000
boostrap replicates.
P.B. Ramírez Rodríguez et al. / Experimental Parasitology 170 (2016) 227e235230

3. Results
3.1. Antibody titers in vaccinated cattle
In this work, we sought to explore the ovaries as a potential
source of new vaccine candidates to fight cattle ticks. To this end,
we dissected ovarian tissue from engorged adult female ticks and
disrupted it to obtain a suspension of total ovarian proteins
(Fig. 2A). This sample was complexed with the adjuvant Montanide
ISA 50 V and used to immunize cattle. Ovarian proteins generated a
strong humoral immune response in vaccinated cattle. Antibody
titers increased after each immunization, peaked at the fourth
week and remained high until the end of the experiment (Fig. 1).
Samples from control animals did not show significant changes in
IgG levels. Since sera with the highest IgG levels were found on the
fourth week, they were utilized for 1D and 2D western blot assays.
3.2. Detection of immunogenic proteins by western blot assays
Sera collected on the fourth post-immunizationweekwere used
as the first antibody against ovarian proteins run on 1D SDS-PAGE
and immunoblots were developed. Some proteins were recog-
nized with sera from immunized cattle but not from control or pre-
immune sera (Fig. 2BeD).
In order to obtain a higher resolution on the protein recognition,
we performed 2D SDS-PAGE-Western blot experiments and found
that sera from immunized cattle detected several ovarian proteins
(Fig. 3). The reactive spots observed on the nitrocellulose mem-
brane were identified in the gel replicate and the corresponding
spots were manually cut out with new sterile micropipette ends,
collected in sterile Eppendorf tubes, digested with trypsin, and
analyzed by LC-MS/MS. Samples were named as they were cut out
from the gel as OvaP1, OvaP2, OvaP3, OvaP4a and OvaP4b (Table 1).
3.3. Identification of immunogenic proteins by LC-MS/MS
Tryptic peptides from samples were analyzed using sensitive
liquid chromatography tandem mass spectrometry (LC-MS/MS).
Proteins were identified as follows: samples OvaP1 and OvaP2were
identified as Vitellogenin, sample OvaP3 as Vitellogenin-2 precur-
sor and samples OvaP4a and OvaP4b as Yolk Cathepsin (Table 1).
4. Discussion
Considerable efforts have been undertaken worldwide to iden-
tify and characterize tickmolecules for the development of novel or
improved anti-tick vaccines. At present, however, there is only one
anti-cattle tick vaccine commercially available in the world, which
is based on the Bm86 antigen and is marketed under the trade
name Gavac™. Past efforts had been focused mainly on gut and
salivary gland antigens, and more recently on other internal organs
as the ovaries, which are essential for tick reproduction. For
example, Rachinsky et al. reported a differential protein expression
in ovaries of uninfected and Babesia-infected R. microplus ticks.
Among the ovarian proteins that were up-regulated in infected
ticks were calreticulin, two myosin subunits, an endoplasmic re-
ticulum protein, a peptidyl-prolyl cis-trans isomerase (PPIase), a
cytochrome c oxidase subunit, a glutamine synthetase, and a family
of Kunitz-type serine protease inhibitors (Rachinsky et al., 2007).
These findings were additionally supported through an ovarian
transcriptome study (Heekin et al., 2013). In the present work, we
followed a classical vaccinology approach, using total ovarian pro-
tein extracts as immunogens, combined with ELISA, 1D and 2D
western blot, and LC-MS/MS analyses. In this discovery phase, we
could identify three immunogenic proteins, namely Vitellogenin,
Vitellogenin-2 precursor and Yolk Cathepsin. Interestingly, despite
its importance, R. microplus tick vitellogenesis process has not been
studied in detail and vitellogenic proteins are just being discovered.
Fig. 2. Electrophoretic pattern of ovarian proteins and immunodetection in 1D-SDS-PAGE. (A) Total ovarian proteins were run on one-dimensional electrophoresis SDS-PAGE (12%)
and stained with Coomassie blue. Gel replicas were transferred to nitrocellulose membranes and incubated separately with control (B), pre-immune (C) or immune sera (D), and
Western blot assays were developed. (1) Molecular weight marker; (2) Ovarian proteins. Arrows indicate the reactive bands. Representative 1D immunoblotting is shown.
P.B. Ramírez Rodríguez et al. / Experimental Parasitology 170 (2016) 227e235 231

Vitellogenin is the precursor for Vitellin, the major yolk protein of
eggs in both vertebrate and invertebrate animals and shares a
common ancestry (Chen et al., 1997). In ticks, Vitellogenin is syn-
thesized in the fat body and midgut, secreted into the hemolymph
and taken up by Vitellogenin receptors on the surfaces of the oo-
cytes and internalized into the oocytes. Inside the oocytes, Vitel-
logenin subunits are recombined and converted to Vitellin, the egg
storage protein (Thompson et al., 2007).
In this study, we hypothesized that sera from immunized cattle
can recognize proteins present in both larvae and ovaries (Fig. 1). If
these ovary-targeted sera contain antibodies that can detect larval
proteins, it would be advantageous, given that molecular targets
would be present and would be recognized since the first tick
parasitic stage. It is known that once vitellogenesis has finished, the
egg contains all substances needed for embryo development during
embryogenesis which occurs outside maternal organism, as well as
the genetic material (Logullo et al., 2002). In particular, Vitellin (a
combination of processed Vitellogenins and the major yolk protein
of eggs) serves as an embryonic nutrient reserve before hatching
and even during non-feeding periods of larval, nymphal and adult
ticks (Rosell-Davis and Coons, 1989). Vitellin subunits have been
found in eggs and unfed larvae in Dermacentor variabilis ticks
(Rosell-Davis and Coons, 1989). Moreover, Vitellin and Vitellin-and
hemoglobin-digesting enzymes such as BYC, VTDCE, THAP and
BmCl1, which are maternally produced yolk proteases and accu-
mulate in oocytes, have been found in eggs and larvae from
R. microplus ticks (Bergamo et al., 2010; Seixas et al., 2012). Even
though immature stages of ticks do not have reproductive organs,
eggs, larvae and even nymphs preserve much of the maternally
transmitted material via the ovaries. The increased antibody titer
observed in Fig. 1, suggests that sera contain antibodies that are
recognizing proteins shared by both larvae and ovaries.
Evidence for multiple Vitellogenins has been reported in ticks.
Thompson et al (Thompson et al., 2007). reported the first complete
Vitellogenin mRNA from ticks. They showed that Vitellogenin
message was detected in Dermacentor variabilis females, after
mating and blood feeding to repletion. This expression was regu-
lated by ecdysteroids. Vitellogenin message was found at high
levels in fat body and midgut, and at low levels in the ovary of pre-
ovipositional (mated, replete) females. Khalil et al (Khalil et al.,
2011). reported a second Vitellogenin gene in D. variabilis. Injec-
tion of virgin females with 20 hydroxyecdysone induced Vitello-
genin 2 expression, vitellogenesis and oviposition. Using RT-PCR
assays, Vitellogenin 2 expression was detected only in female ticks
after mating and feeding to repletion. Northern blot analysis
showed that Vitellogenin 2 is expressed in fat body and midgut of
vitellogenic females, but not in the ovary. Meanwhile, Boldbaatar
et al (Boldbaatar et al., 2010). have reported the presence of mul-
tiple Vitellogenins in Haemaphysalis longicornis ticks. The expres-
sion profiles of three Vitellogenin genes were determined by RT-
PCR analyses using total RNA samples from different tissues of
partially fed adult female ticks of the parthenogenetic race.
Vitellogenin-1 was expressed in the midgut only. Vitellogenin-2
was expressed in the ovary and fat body, and Vitellogenin-3 was
expressed only in the fat body. No Vitellogenin expression was
detected in the salivary glands or Malpighian tubules. Accumu-
lating evidence also indicates the existence of several Vitellogenins
in R. microplus ticks (Fig. 4).
In 2002, the GP80 protein, in fact the C-terminal fragment of
Vitellogenin-1, was reported as having a protective role against tick
infestations on immunized sheep (Tellam et al., 2002). Recently,
Tirloni et al. (Tirloni et al., 2014), reported the encoding sequences
of Vitellogenin-1, Vitellogenin-2 and Vitellogenin-3 from
R. microplus Porto Alegre tick strain. These genes were apparently
Fig. 3. Detection of immunogenic proteins of R. microplus in 2D-SDS-PAGE. (A) Total ovarian proteins of R. microplus were run on two-dimensional electrophoresis SDS-PAGE (12%,
pH 3e10) and stained with Coomassie blue. Arrows indicate the samples cut out from the gel. (B) The gel replica was transferred to a nitrocellulose membrane and this was
incubated with serum from cattle immunized with ovary proteins, which was used as the first antibody for the Western blot assay. Arrows indicate the reactive bands. Repre-
sentative 2D immunoblotting is shown.
Table 1
Ovary proteins identified by LC-MS/MS analysis.
Spot Name gi Protein Identified Mass (AMU) IP Sequence
1 OvaP1 82878755 Vitellogenin 85 6.0 FVLSAGK
2 OvaP2 82878755 Vitellogenin 65 4.8 FVLSAGK
3 OvaP3 523713961 Vitellogenin-2 Precursor 75 5.5 YTGTLLGLCGD
4a OvaP4a 62199768 Yolk Cathepsin 60 4.8 VYTIFK
4b OvaP4b 62199768 Yolk Cathepsin 50 4.8 VYTIFK
P.B. Ramírez Rodríguez et al. / Experimental Parasitology 170 (2016) 227e235232
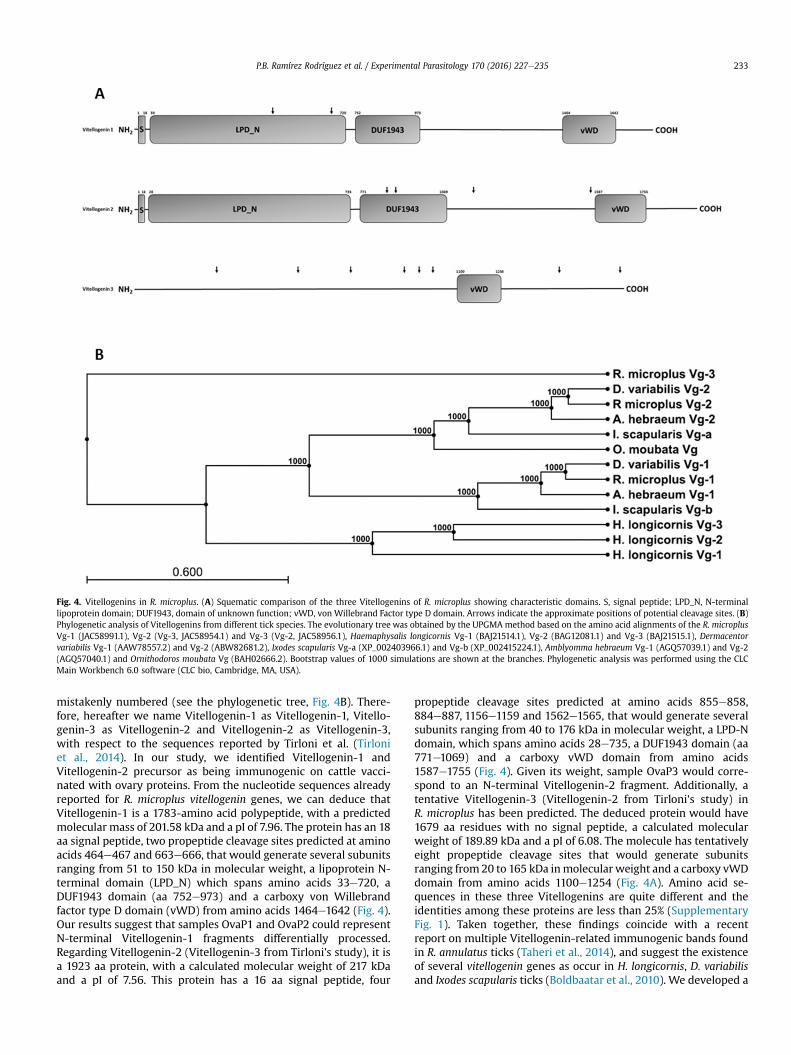
mistakenly numbered (see the phylogenetic tree, Fig. 4B). There-
fore, hereafter we name Vitellogenin-1 as Vitellogenin-1, Vitello-
genin-3 as Vitellogenin-2 and Vitellogenin-2 as Vitellogenin-3,
with respect to the sequences reported by Tirloni et al. (Tirloni
et al., 2014). In our study, we identified Vitellogenin-1 and
Vitellogenin-2 precursor as being immunogenic on cattle vacci-
nated with ovary proteins. From the nucleotide sequences already
reported for R. microplus vitellogenin genes, we can deduce that
Vitellogenin-1 is a 1783-amino acid polypeptide, with a predicted
molecular mass of 201.58 kDa and a pI of 7.96. The protein has an 18
aa signal peptide, two propeptide cleavage sites predicted at amino
acids 464e467 and 663e666, that would generate several subunits
ranging from 51 to 150 kDa in molecular weight, a lipoprotein N-
terminal domain (LPD_N) which spans amino acids 33e720, a
DUF1943 domain (aa 752e973) and a carboxy von Willebrand
factor type D domain (vWD) from amino acids 1464e1642 (Fig. 4).
Our results suggest that samples OvaP1 and OvaP2 could represent
N-terminal Vitellogenin-1 fragments differentially processed.
Regarding Vitellogenin-2 (Vitellogenin-3 from Tirloni's study), it is
a 1923 aa protein, with a calculated molecular weight of 217 kDa
and a pI of 7.56. This protein has a 16 aa signal peptide, four
propeptide cleavage sites predicted at amino acids 855e858,
884e887, 1156e1159 and 1562e1565, that would generate several
subunits ranging from 40 to 176 kDa in molecular weight, a LPD-N
domain, which spans amino acids 28e735, a DUF1943 domain (aa
771e1069) and a carboxy vWD domain from amino acids
1587e1755 (Fig. 4). Given its weight, sample OvaP3 would corre-
spond to an N-terminal Vitellogenin-2 fragment. Additionally, a
tentative Vitellogenin-3 (Vitellogenin-2 from Tirloni's study) in
R. microplus has been predicted. The deduced protein would have
1679 aa residues with no signal peptide, a calculated molecular
weight of 189.89 kDa and a pI of 6.08. The molecule has tentatively
eight propeptide cleavage sites that would generate subunits
ranging from20 to 165 kDa inmolecular weight and a carboxy vWD
domain from amino acids 1100e1254 (Fig. 4A). Amino acid se-
quences in these three Vitellogenins are quite different and the
identities among these proteins are less than 25% (Supplementary
Fig. 1). Taken together, these findings coincide with a recent
report on multiple Vitellogenin-related immunogenic bands found
in R. annulatus ticks (Taheri et al., 2014), and suggest the existence
of several vitellogenin genes as occur in H. longicornis, D. variabilis
and Ixodes scapularis ticks (Boldbaatar et al., 2010). We developed a
Fig. 4. Vitellogenins in R. microplus. (A) Squematic comparison of the three Vitellogenins of R. microplus showing characteristic domains. S, signal peptide; LPD_N, N-terminal
lipoprotein domain; DUF1943, domain of unknown function; vWD, von Willebrand Factor type D domain. Arrows indicate the approximate positions of potential cleavage sites. (B)
Phylogenetic analysis of Vitellogenins from different tick species. The evolutionary tree was obtained by the UPGMA method based on the amino acid alignments of the R. microplus
Vg-1 (JAC58991.1), Vg-2 (Vg-3, JAC58954.1) and Vg-3 (Vg-2, JAC58956.1), Haemaphysalis longicornis Vg-1 (BAJ21514.1), Vg-2 (BAG12081.1) and Vg-3 (BAJ21515.1), Dermacentor
variabilis Vg-1 (AAW78557.2) and Vg-2 (ABW82681.2), Ixodes scapularis Vg-a (XP_002403966.1) and Vg-b (XP_002415224.1), Amblyomma hebraeum Vg-1 (AGQ57039.1) and Vg-2
(AGQ57040.1) and Ornithodoros moubata Vg (BAH02666.2). Bootstrap values of 1000 simulations are shown at the branches. Phylogenetic analysis was performed using the CLC
Main Workbench 6.0 software (CLC bio, Cambridge, MA, USA).
P.B. Ramírez Rodríguez et al. / Experimental Parasitology 170 (2016) 227e235 233

phylogenetic tree using the full amino acid sequences of Vitello-
genins from several tick species (Fig. 4B). In this analysis,
Vitellogenin-1 from D. variabilis, R. microplus and A. hebraeum
formed a cluster together. Vitellogenin-2 (Vitellogenin-3 from Tir-
loni's study) formed a cluster with Vitellogenin-2 from D. variabilis
and A. hebraeum with a 100% bootstrap probability, whereas
Vitellogenin-3 from R. microplus (Vitellogenin-2 from Tirloni's
study) clustered on a separate branch. Additionally, we wanted to
confirm the mRNA expression of these three Vitellogenins on
ovaries from engorged R. microplus female ticks (Fig. 5).
As can be seen, vitellogenin 1 and vitellogenin 2 are expressed in
high levels, whereas vitellogenin 3 is not expressed. This suggests
that in R. microplus, Vitellogenin 3 would be imported to ovaries
from extra-ovarian tissues, such as fat body or midgut. However,
Vitellogenin 1 and Vitellogenin 2 would be actively synthesized in
the ovaries, apart from other possible sites. Ticks show multiple
tissue sources for vitellogenins mRNAs, i.e., fat body, midgut and
ovary, but these molecules are expressed in a tissue-specific
manner among ticks. The relative contribution of each vitellogenin
gene during vitellogenesis at present is, however, not clear. More-
over, the proteolytic mechanisms resulting in small subunit prod-
ucts of different types of Vitellogenins and their utilization as yolk
protein during oocyte development, need to be elucidated. On the
other hand, samples OvaP4a and OvaP4b were identified as Yolk
Cathepsin. This protein is implicated in Vitellin hydrolysis in
R. microplus eggs (Logullo et al., 1998) and has been evaluated in
both native and recombinant forms, eliciting humoral response in
immunized cattle, and protecting them against R. microplus infes-
tation (Seixas et al., 2012). This scenario shows the apparent
complexity on ovarian tickmolecular targets and raises the need for
a rational and integrative approach to select and design anti-tick
vaccine candidates.
5. Conclusion
We identified three immunogenic proteins from ovaries of
R. microplus, following an immunoproteomic approach. Further-
more, we demonstrated that the ovaries of R. microplus act as an
active synthesis site of Vitellogenins. Our findings will allow us to
better understand R. microplus ovarian maturation, as well as to
explore their potential as vaccine candidates. The identified pro-
teins merit further investigation because they play an essential role
on tick reproduction. It is known that Vitellin (a combination of
processed Vitellogenins), serves as an embryonic nutrient reserve
before hatching and even during non-feeding periods of larval,
nymphal and adult ticks (Rosell-Davis and Coons, 1989). Vitellin
content decreases as development proceeds, however, once ticks
mate, and the rapid-engorgement feeding period commences,
synthesis of new Vitellogenin molecules occurs. These events
would secure a constant availability of molecular targets in ticks
feeding upon immunized cattle with these antigens (Vitellogenins).
This view is additionally supported by the evidence that dsRNA-
mediated silencing of three Vitellogenin genes affected egg devel-
opment and oviposition in H. longicornis ticks (Boldbaatar et al.,
2010). In brief, vaccines targeting these ovarian proteins offer the
opportunity to disrupt tick reproduction and thus, potentially
control R. microplus tick populations. Ongoing immunization ex-
periments will demonstrate their protective efficacy against tick
infestations.
Conflict of interest
The authors have declared that no competing interests exist.
Acknowledgements
This work was supported by Grants SEP-CONACYT-CB-2008-01-
104006, Proyectos de Desarrollo Científico para Atender Problemas
Nacionales-2013-PDCPN2013-01-216321 and CONACYT-Programa
de Estancias Sab�aticas Nacionales, Estancias Sab�aticas al Extranjero
y Estancias Cortas para la Consolidaci�on de Grupos de Investigaci�on
2014-246310 to MMV. PBRR and RELQ received a scholarship from
CONACYT. The authors thank Kirk Allen for support in drafting and
revising this manuscript.
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://
dx.doi.org/10.1016/j.exppara.2016.10.005.
References
Bergamo, A., Seixas, A., de Oliveira, V., Michel, A.F., Termignoni, C., 2010. Vitellin-and hemoglobin-digesting enzymes in Rhipicephalus (Boophilus) micropluslarvae and females. Comp. Biochem. Phys. B 157, 326e335.
Boldbaatar, D., Umemiya-Shirafuji, R., Liao, M., Tanaka, T., Xuan, X., Fujisaki, K., 2010.Multiple vitellogenins from the Haemaphysalis longicornis tick are crucial forovarian development. J. Insect. Physiol. 56, 1587e1598.
Chen, J.S., Sappington, T.W., Raikhel, A.S., 1997. Extensive sequence conservationamong insect, nematode, and vertebrate vitellogenins reveals ancient commonancestry. J. Mol. Evol. 44, 440e451.
Crampton, A.L., Green, P., Baxter, G.D., Barker, S.C., 1999. Monooxygenases play onlya minor role in resistance to synthetic pyrethroids in the cattle tick, Boophilusmicroplus. Exp. Appl. Acarol. 23, 897e905.
de la Fuente, J., Almaz�an, C., Canales, M., P�erez de la Lastra, J.M., Kocan, K.M.,Willadsen, P., 2007. A ten-year review of commercial vaccine performance forcontrol of tick infestations on cattle. Anim. Health Res. Rev. 8, 23e28.
García-García, J.C., Montero, C., Redondo, M., Vargas, M., Canales, M., Boue, O.,Rodríguez, M., Joglar, M., Machado, H., Gonz�alez, I.L., Vald�es, M., M�endez, L., dela Fuente, J., 2000. Control of ticks resistant to immunization with Bm86 incattle vaccinated with the recombinant antigen Bm95 isolated from the cattletick, Boophilus microplus. Vaccine 18, 2275e2287.
Gough, J.M., Kemp, D.H., 1993. Localization of a low abundance membrane protein(Bm86) on the gut cells of the cattle tick Boophilus microplus by immunogoldlabeling. J. Parasitol. 79, 900e907.
Hawkes, R., Niday, E., Gordon, J., 1982. A dot-immunobinding assay for monoclonaland other antibodies. Anal. Biochem. 119, 142e147.
Heekin, A.M., Guerrero, F.D., Bendele, K.G., Saldivar, L., Scoles, G.A., Dowd, S.E.,Gondro, C., Nene, V., Djikeng, A., Brayton, K.A., 2013. The ovarian transcriptomeof the cattle tick, Rhipicephalus (Boophilus) microplus, feeding upon a bovine
Fig. 5. Vitellogenins expression in ovaries of R. microplus. RT-PCR reactions were per-
formed using total RNA of ovaries from engorged adult female ticks. Actinwas used as a
control. (1) 100 bp molecular weight marker; (2) Vitellogenin 1 (219 bp); (3) Vitello-
genin 3; (4) Vitellogenin 2 (162 bp); (5) actin (140 bp).
P.B. Ramírez Rodríguez et al. / Experimental Parasitology 170 (2016) 227e235234

host infected with Babesia bovis. Parasit. Vectors 6, 276.Khalil, S.M.S., Donohue, K.V., Thompson, D.M., Jeffers, L.A., Ananthapadmanaban, U.,
Sonenshine, D.E., Mitchell, R.D., Roe, R.M., 2011. Full-length sequence, regula-tion and developmental studies of a second vitellogenin gene from the Amer-ican dog tick, Dermacentor variabilis. J. Insect Physiol. 57, 400e408.
Klafke, G.M., Sabatini, G.A., de Albuquerque, T.A., Martins, J.R., Kemp, D.H.,Miller, R.J., Schumaker, T.T.S., 2006. Larval immersion tests with ivermectin inpopulations of the cattle tick Rhipicephalus (Boophilus) microplus (Acari: ixo-didae) from State of Sao Paulo, Brazil. Vet. Parasitol. 142, 386e390.
Logullo, C., da Silva Vaz, I., Sorgine, M., Paiva-Silva, G.O., Faria, F.S., Zingali, R., DeLima, M.F.R., Abreu, L., Fialho Oliveira, E., Alves, E.W., Masuda, H., Gonz�ales, J.C.,Masuda, A., Oliveira, P.L., 1998. Isolation of an aspartic proteinase precursorfrom the egg of a hard tick, Boophilus microplus. Parasitology 116, 525e532.
Logullo, C., Moraes, J., Dansa-Petretski, M., Vaz, I.S., Masuda, A., Sorgine, M.H.F.,Braz, G.R., Masuda, H., Oliveira, P.L., 2002. Binding and storage of heme byvitellin from the cattle tick, Boophilus microplus. Insect biochem. Mol. Biol. 32,1805e1811.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurementwith the Folin phenol reagent. J. Biol. Chem. 193, 265e275.
Maritz-Olivier, C., van Zyl, W., Stutzer, C., 2012. A systematic, functional genomics,and reverse vaccinology approach to the identification of vaccine candidates inthe cattle tick, Rhipicephalus microplus. Ticks Tick-borne Dis. 3, 179e187.
Martins, J.R., Furlong, J., 2001. Avermectin resistance of the cattle tick Boophilusmicroplus in Brazil. Vet. Rec. 149, 64.
Martins, J.R., Evans, D.E., Ceres�er, V.H., Correa, B.L., 2002. Partial strategic tickcontrol within a herd of European breed cattle in the state of Rio Grande do Sul,southern Brazil. Exp. Appl. Acarol. 27, 241e251.
Parizi, L.F., Pohl, P.C., Masuda, A., Vaz Junior, I.D., 2009. New approaches towardanti-Rhipicephalus (Boophilus) microplus tick vaccine. Rev. Bras. Parasitol. Vet 18,1e7.
P�erez de Le�on, A.A., Strickman, D.A., Knowles, D.P., Fish, D., Thacker, E., Fuente, J.,Krause, P.J., Wikel, S.K., Miller, R.S., Wagner, G.G., Almaz�an, C., Hillman, R.,Messenger, M.T., Ugstad, P.O., Duhaime, R.A., Teel, P.D., Ortega-Santos, A.,Hewitt, D.G., Bowers, E.J., Bent, S.J., Cochran, M.H., McElwain, T.F., Scoles, G.A.,Suarez, C.E., Davey, R., Freeman, J.M.H., Lohmeyer, K., Li, A.Y., Guerrero, F.D.,Kammlah, D.M., Phillips, P., Pound, J.M., 2010. One Health approach to identifyresearch needs in bovine and human babesioses; workshop report. Parasit.Vectors 3, 36.
Rachinsky, A., Guerrero, F.D., Scoles, G.A., 2007. Differential protein expression inovaries of uninfected and Babesia-infected southern cattle ticks, Rhipicephalus(Boophilus) microplus. Insect biochem. Mol. Biol. 37, 1291e1308.
Rand, K.N., Moore, T., Sriskantha, A., Spring, K., Tellam, R., Willadsen, P., Cobon, G.S.,1989. Cloning and expression of a protective antigen from the cattle tick Boo-philus microplus. Proc. Natl. Acad. Sci. 86, 9657e9661.
Rodríguez, V.M., 2000. Respuesta inmunol�ogica contra garrapatas. Biotecnol. Apl. 17,215e220.
Rodriguez-Valle, M., Vance, M., Moolhuijzen, P.M., Tao, X., Lew-Tabor, A.E., 2012.Differential recognition by tick-resistant cattle of the recombinantly expressedRhipicephalus microplus serine protease inhibitor-3 (RMS-3). Ticks Tick-borneDis. 3, 159e169.
Rosell-Davis, R., Coons, L.B., 1989. Relationship between feeding, mating, vitello-genin production and vitellogenesis in the tick Dermacentor variabilis. Exp.Appl. Acarol. 7, 95e105.
Rozen, S., Skaletsky, H.J., 2000. Primer3 on the WWW for general users and forbiologist programmers. In: Krawetz, S., Misener, S. (Eds.), BioinformaticsMethods and Protocols: Methods in Molecular Biology. Humana, Totowa,pp. 365e386.
Seixas, A., Oliveira, P., Termignoni, C., Logullo, C., Masuda, A., 2012. V.Jr.I. da Silva,Rhipicephalus (Boophilus) microplus embryo proteins as target for tick vaccine.Vet. Immunol. Immunopathol. 148, 149e156.
Shahein, Y.E., EI-Hakim, A.E., Abouelella, A.M., Hamed, R.R., Allam, S.A., Farid, N.M.,2008. Molecular cloning, expression and characterization of a functional GSTmuclass from the cattle tick Boophilus annulatus. Vet. Parasitol. 152, 116e126.
Taheri, M., Nabian, S., Ranjbar, M., Mazaheri, N.F.R., Gerami, S.A., Sazmand, A., 2014.Study of vitellogenin in Boophilus annulatus tick larvae and its immunologicalaspects. Trop. Biomed. 31, 398e405.
Tellam, R.L., Kemp, D., Riding, G., Briscoe, S., Smith, D., Sharp, P., Irving, D.,Willadsen, P., 2002. Reduced oviposition of Boophilus microplus feeding onsheep vaccinated with vitellin. Vet. Parasitol. 103, 141e156.
Thompson, D.M., Khalil, S.M., Jeffers, L.A., Sonenshine, D.E., Mitchell, R.D.,Osgood, C.J., Roe, R.M., 2007. Sequence and the development and tissue-specificregulation of the first complete vitellogenin messenger RNA from ticksresponsible for heme sequestration. Insect biochem. Mol. Biol. 37, 363e374.
Tirloni, L., Reck, J., Soares, T.R.M., Martins, J.R., Mulenga, A., Sherman, N.E., Fox, J.W.,Yates III, J.R., Termignoni, C., Pinto, A.F.M., da Silva, V. Jr. I., 2014. Proteomicanalysis of cattle tick Rhipicephalus (Boophilus) microplus saliva: a comparisonbetween partially and fully engorged females. PLoS One 9, e94831.
Towbin, H., Staehelien, T., Gordon, J., 1979. Electrophoretic transfer of proteins frompolyacrylamide gels to nitrocellulose sheets: procedure and some applications.Proc. Nat. Acad. Sci. U.S.A. 76, 4350e4354.
Wang, H., Nuttall, P.A., 1999. Immunoglobulin-binding proteins in ticks: new targetfor vaccine development against a blood-feeding parasite. Cell. Mol. Life Sci. 56,286e295.
Willadsen, P., Riding, G.A., McKenna, R.V., Kemp, D.H., Tellam, R.L., Nielsen, J.N.,Lahnstein, J., Cobon, G.S., Gough, J.M., 1989. Immunologic control of a parasiticarthropod. Identification of a protective antigen from Boophilus microplus.J. Immunol. 143, 1346e1351.
P.B. Ramírez Rodríguez et al. / Experimental Parasitology 170 (2016) 227e235 235