molecular cloning and expression of the regulatory (rg~) subunit
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY (CJ 1991 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 266, No. 24, Issue of August 25, pp. 15782-15789.1991 Printed in U.S.A.
Molecular Cloning and Expression of the Regulatory (RG~) Subunit of the Glycogen-associated Protein Phosphatase*
(Received for publication, April 12, 1991)
Pauline M. Tang$, Jeffry A. Bondor, Kristine M. Swidereks, and Anna A. DePaoli-Roachn From the Department of Biochemistry and Molecular Biology, Indiana University School of Medicine, Indianapolis, Indiana 46202-5122
DNA clones encoding the glycogen-binding (Rcl) sub- unit of glycogen-associated protein phosphatase were isolated from rabbit skeletal muscle Xgtll cDNA li- braries. Overlapping clones provided an open reading frame of 3327 nucleotides that predicts a polypeptide of 1109 amino acids with a molecular weight of 124,257. Northern hybridization of rabbit RNA iden- tified a major mRNA transcript of 7.5 kilobases pres- ent in skeletal, diaphragm, and cardiac muscle, but not in brain, kidney, liver, and lung. Southern analysis of rabbit genomic DNA digested with various restriction endonucleases gave rise to a single hybridizing frag- ment, suggesting that a single gene is present. Expres- sion of the complete Rcl subunit coding sequence in Escherichia coli generated a protein of apparent mo- lecular weight on sodium dodecyl sulfate-polyacryl- amide gel electrophoresis of approximately 160,000, similar to the size of the polypeptide detected by West- ern immunoblot in rabbit skeletal muscle extracts. The Rcl subunit shares significant homology with the Sac- charomyces cerevisiae GACl gene product which is involved in activation of glycogen synthase and gly- cogen accumulation. The homology with GACl sub- stantiates the role of this enzyme in control of glycogen metabolism. Hydropathy analysis of the Rc, subunit amino acid sequence revealed the presence of a hydro- phobic region in the COOH terminus, suggesting a potential association with membrane. This result sug- gests that the same phosphatase regulatory component may be involved in targeting the enzyme both to mem- branes and to glycogen.
Protein phosphorylation is a major mechanism by which many cellular functions are regulated. The co-ordinated con-
* This work was supported in part by National Institutes of Health Grants DK36569 and PO1 HL06308proj.7 and by the Grace M. Foundation. The oligonucleotides used in this study were synthesized in the Biochemistry Biotechnology Facility in our department, partly supported by Indiana University Diabetes Research and Training Center Grant DK20542. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequence($ reported in this paper has been submitted to the GenBank““/EMBL Data Bank with accession numbeds) M65 109.
$ Partially supported by a postdoctoral fellowship from the Amer- ican Heart Association, Indiana Affiliate, Inc.
Present address: Beckman Research Institute of City of Hope, Division of Immunology, 1450 E. Duarte Rd., Duarte, CA 91010-0269.
1 Recipient of Research Career Development Award DK01690. To whom correspondence should be addressed: Dept. of Biochemistry and Molecular Biology, Indiana University, School of Medicine, 635 Barnhill Drive, Indianapolis, Indiana 46202-5122.
trol of both types of interconverting enzymes, protein kinases and protein phosphatases, determines the phosphorylation and hence modulation of the activity of intracellular target proteins. Regulation of protein kinases has been extensively studied and for some forms it is reasonably well understood (1-6), whereas knowledge and understanding of the potential control of protein phosphatases is incomplete. The major cellular serine/threonine protein phosphatases have been classified into two categories, type 1 and type 2, according to their substrate specificities and their sensitivities to inhibitor proteins (7, 8). The cDNA coding the catalytic subunits of four forms (type 1, 2A, 2B, and 2C) included in this classifi- cation have been cloned (9-16), and with the exception of the 2C protein phosphatase, all appear to be derived from the same gene family. At least four forms of type 1 phosphatases differing in the associated regulatory subunit have been iden- tified (a) the ATP-Mg-dependent phosphatase (17-19), ( b ) the glycogen-associated phosphatase (20), (c) the sarco- plasmic reticulum-associated phosphatase (21, 22), and ( d ) the myosin-associated phosphatase (23, 24). There is also evidence for nuclear associated type 1 phosphatases (25, 26). They all appear to share a similar 37-kDa catalytic subunit complexed to different regulatory subunits that are responsi- ble for regulation and/or targeting of the enzymes to specific cellular locales (8,9). These regulatory subunits are (a ) inhib- itor-2, ( b ) glycogen-binding or RG, subunit,’ (c) sarcoplasmic reticulum-binding subunit, and ( d ) a putative myosin-binding subunit. A recent report has shown that the properties of the sarcoplasmic reticulum-associatedphosphatase in rabbit skel- etal muscle are similar to those of the glycogen-bound phos- phatase, suggesting that the &I subunit might play a dual role in targeting type 1 phosphatase to two different subcel- lular locations, glycogen and membranes (22).
Up to 60% of rabbit skeletal muscle phosphorylase phos- phatase is associated with glycogen (27, 28). This glycogen- bound protein phosphatase was first identified and purified as a 137-kDa heterodimer, consisting of a 37-kDa catalytic subunit and a 103-kDa regulatory component, the &I subunit (20). However, the Rcl subunit is extremely sensitive to pro- teolysis. More recent analysis by Western immunoblotting
The abbreviations used are: Rcl, glycogen-binding regulatory sub- unit of type 1 protein phosphatase. This protein had previously been designated “G subunit,” causing some confusion with GTP binding proteins. We propose the present nomenclature as a means to avoid this confusion. In addition, other type 1 phosphatase regulatory subunits can be named in a parallel fashion, RSR and R M ~ for the sarcoplasmic reticulum and myosin associated forms, respectively, and RI.* for inhibitor-2. The other abbreviations used are: HX, random hexamer-primed DT, oligo(dT)-primed SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; kb, kilobase(s); IPTG, isopropyl-(3-thio-galactopyranoside; TLCK, 1-chloro-3-tosy- lamido-7-amino-2-heptanone; EGTA, [ethylenebis(oxyethylene- nitri1o)tetraacetic acid.
15782

Protein Phosphatase 15783
indicated that the intact subunit has a M, of 160,000- 170,000 (28, 29). Its function appears to be targeting the phosphatase to the glycogen particle, where several of the enzymes, such as glycogen synthase and phosphorylase ki- nase, involved in glycogen metabolism are located. The &l
subunit is phosphorylated in vitro by the CAMP-dependent protein kinase at two sites (29). Phosphorylation of site 2 has been proposed to cause dissociation of the catalytic subunit (30). Studies from our (31) and Cohen's (32) laboratories also revealed a complex multisite phosphorylation of the Rcl sub- unit. Phosphorylation by CAMP-dependent protein kinase formed the recognition sites for other protein kinases such as glycogen synthase kinase-3 and casein kinase 11.' The CAMP- dependent protein kinase sites and one of the glycogen syn- thase kinase-3 sites (33) have been shown to be phosphoryl- ated in vivo. Epinephrine is reported to enhance significantly the phosphate content of site 2 (33), whereas insulin leads to increased phosphorylation of site 1 (34).
To investigate the native structure and the role of the &I
subunit in the regulation of the glycogen-associated phospha- tase we have undertaken the molecular cloning of cDNAs coding for this subunit. This paper reports the first isolation and characterization of cDNA clones encoding the Rcl sub- unit. The deduced amino acid sequence of the entire trans- lated region, the tissue specific distribution, and the expres- sion of the coding sequences in Escherichia coli are presented.
EXPERIMENTAL PROEDURES
Materials-The bacteriophage T7 polymerase expression system was generously provided by Dr. F. W. Studier (Brookhaven National Laboratories). Oligonucleotides were synthesized in an Applied Bio- systems DNA synthesizer model 380A. Restriction and other DNA modifying enzymes, M13 vectors, and agarose were purchased from Bethesda Research Laboratories. Genescribe-Z vectors and random- primed DNA labeling kits were obtained from United States Bio- chemicals Corp. Deoxy- and dideoxy nucleotide triphosphates were from Pharmacia LKB Biotechnology Inc. Radionucleotides were pur- chased from Du Pont-New England Nuclear, and 1251-protein A was from ICN. Other general reagents were from Bethesda Research Laboratories, Sigma, and Boehringer Mannheim.
Isolation and Sequence Determination of cDNA Clones-Two rabbit skeletal muscle random hexamer-primed Xgtll cDNA libraries con- taining, respectively, lo6 and 6 X lo6 independent recombinants and one rabbit muscle oligo(dT)-primed Xgtll cDNA library containing lo7 independent recombinants were constructed (35). To isolate cDNA clones encoding the Rcl subunit, two oligonucleotide probes were synthesized, corresponding to the amino acid sequences around the CAMP-dependent protein kinase phosphorylation sites: G- COMP1,GGCGGTGTGCACGTACACCTCCTCandG-COMP2,TT- (G/A)AA(G/C)CC(G/A)AA(G/A)TT(G/A)TCGGCGAA. The choice of the codons was based on the most frequent codon usage for rabbit (36). The two oligonucleotides, 5'-end-labeled with T4 polynucleotide kinase and [T-~~PJATP, were used to screen on duplicate filters approximately 160,000 recombinants from the rabbit skeletal muscle random hexamer-primed unamplified library. Hybridization was per- formed at 50 "C in a solution containing 10 X Denhardt's (0.2% (w/ v) each of Ficoll, polyvinylpyrrolidone, and gelatin), 6 X SSPE (1 X SSPE: 0.15 M sodium chloride, 10 mM sodium phosphate, and 1 mM EDTA, pH 7.4), 0.05% NaPPi, 0.1% SDS, and the radiolabeled probe (2 X lo6 cpm/ml). Nitrocellulose filters were washed in 6 X SSC (1 X SSC: 0.15 M sodium chloride and 15 mM sodium citrate, pH 7) at room temperature for 10 min twice followed by 20 min at 50 "C. Positive clones were plaque-purified by consecutive screening. DNA from positive recombinant phages was isolated (37) and the cDNA inserts were subcloned into the GeneScribe-Z vector, pTZ19U, for restriction endonuclease analysis and into M13 vectors for DNA sequencing (38). To obtain the entire coding sequences, confirmed cDNA fragments labeled by the random hexamer priming method (39) were used to screen additional 310,000 recombinant phages from
V. Cerovsky, K. M. Swiderek, J. H. Haseman, C. J. Fiol, R. W. Roeske, P. J. Roach, and A. A. DePaoli-Roach, manuscript in prep- aration.
the random hexamer-primed library and 160,000 recombinants from the oligo(dT)-primed library. Hybridization of the filters was carried out at 55 "C in the solution described above. Phages yielding positive signals were isolated and the cDNA inserts subcloned in pTZ19U for double-stranded DNA sequencing by the dideoxy chain termination method (40) utilizing vector- and cDNA-specific oligonucleotide primers.
Isolation of Genomic DNA Clones-A rabbit genomic library con- structed in X phage Charon 4A (41), kindly provided by Dr. R. C. Hardison (The Pennsylvania State University), was screened (3.8 X lo5 plaque-forming units) with the cDNA fragment (995 bp) obtained from one clone (HX 1-1) and an oligonucleotide corresponding to nucleotides 61-79. One positive clone was identified. The DNA was isolated, digested with restriction enzymes, and analyzed by a South- ern blot (42). Fragments hybridizing with the cDNA probe were subcloned into pTZ19U vector for sequencing, utilizing vector- and cDNA-specific oligonucleotide primers.
Northern Blot Analysis-Total RNA from rabbit brain, kidney, liver, lung and skeletal, diaphragm, and heart muscle was isolated by the method of Chirgwin et al. (43). RNA samples (15 pg) were electrophoresed through a 0.8% agarose/formaldehyde gel and trans- ferred to nitrocellulose membrane in 10 X SSC (44). Prehybridization was carried out at 55 "C in a solution containing 10 X Denhardt's, 6 X SSPE, 0.05% sodium pyrophosphate, 0.1% SDS, and 0.1 mg/ml Torula RNA. The membrane was hybridized at 55 "C with two 32P- labeled cDNA fragments (2 X lo6 cpm/ml) comprising a total of 3854 bp. After hybridization, the filter was washed two times for 10 min at room temperature followed by 30 min at 68 "C. Autoradiography was performed utilizing Du Pont Quanta 111 intensifying screens.
Primer Extension-A 19-mer synthetic oligonucleotide comple- mentary to residues 61-79 of rabbit Rcl subunit cDNA was used as a primer. The "P-end-labeled oligonucleotide (1.5 X lo6 cpm) was annealed to 25 pg of rabbit skeletal muscle, 30 yg of diaphragm or lung total RNA. The extension reaction was carried at 37 "C for 1 h in 50-pl volume containing first strand buffer (50 mM Tris-HC1, pH 8.3, 75 mM, KCI, 3 mM MgC12, and 10 mM dithiothreitol), 2.5 mM dNTPs, 500 units of mouse Moloney leukemia virus reverse transcrip- tase (Bethesda Research Laboratories), and 625 pg of actinomycin D. The resulting cDNA was analyzed and compared with the sequence of the 6.6-kb SphI genomic DNA fragment on a 6% polyacrylamide sequencing gel.
Southern Analysis of Genomic DNA-Rabbit genomic DNA (20 pg) digested with the restriction enzymes EcoRI, HindIII, and XbaI was separated on a 1% agarose gel and transferred to a nitrocellulose membrane. Following prehybridization at 68 "C in a solution contain- ing 10 X Denhardt's, 6 X SSPE, 0.05% sodium pyrophosphate, 0.1% SDS, and 0.1 mg/ml Torula RNA, the membrane was hybridized with the 995-bp cDNA insert of clone HX 1-1 labeled by the nick trans- lation method (1 X lo6 cpm/ml)(45). The membrane was washed twice for 10 min at room temperature and once for 40 min at 68 "C in 6 X SSC, 0.1% SDS, and 0.05% sodium pyrophosphate followed by a further wash at 68 "C in 2 X SSC for 30 min.
Construction of G.pET-8C Expression Vector-Three cDNA clones, HX 1-1 (995 bp), HX 5-1 (1412 bp), and HX 1-2 (2951 bp), were used to assemble the entire coding sequence. The 341-bp EcoRI-BglII portion of clone HX 1-1 was ligated with the 1102-bp BglII-EcoRI portion of clone HX 5-1 at the BglII site to generate a DNA fragment containing the 5'-most sequences. The resulting DNA fragment was digested with either RsaI and BglII or with SphI and BgZlI to produce a 340-bp RsaI-BglII and a 746-bp BglII-SphI DNA fragment. Two complementary oligonucleotides (18-mer and 14-mer), corresponding to the sequences from the start ATG to the RsaI site at nucleotide 28, were synthesized and annealed to generate an adaptor containing a cohesive NcoI site at the 5'-end and a RsaI site at the 3'-end. This adaptor was ligated to the RsaI-BglII which was subsequently joined to the BglII-SphI fragment. The resulting NcoI-SphI fragment was further ligated to a SphI-BamHI fragment derived from the HX 1-2 clone, giving rise to a 3872-bp NcoI-BamHI fragment, which was then inserted into the PET-8c expression vector (46, 47) cleaved at NcoI and BamHI sites. This plasmid will be referred to as G.pET-8c. The nucleotide sequences in the promoter region and the adaptor oligo- nucleotides were confirmed by sequencing.
DNA was transfected into E. coli strain BL21(DE3), which contained Expression of the RCL Subunit in E. coli-The plasmid G.pET-8c
the T7 RNA polymerase gene integrated in the chromosome under the control of lacUV5 promoter. Cultures were grown at 37 "C in 2 X YT medium containing ampicillin (50 pg/ml). When the culture reached an AeO0 of 0.85 units, the T7 RNA polymerase was induced

15784 Protein Phosphatase
1 3339 3876bp Bel11 RsaI
~ s ; l l WIII ACZIRS~I sph~ ~ s ; l l Ann Accl R5;u Xbai Beln ka Hind111 LI I II I I I I I I I I I I
FIG, 1. Rcl subunit DNA clones. A partial restriction map and all the clones used for DNA sequence analysis are shown. H X denotes clones isolated from a random hexamer-primed rabbit skele- tal muscle Xgtll cDNA library and DT clones obtained from an oligo(dT)- primed library. GG10-1 was a clone iso- lated from a genomic library. The black - bar indicates the coding region; thick lines indicate regions that were se- quenced in both directions; thin lines indicate regions sequenced in one direc- tion; dashed lines indicate regions not sequenced; and the dotted line indicates a portion of an intron in the genomic DNA clone.
Hxl-1 (.935kb)
M13-2 (3.m kb)
Hxl-2 (2.95 kb)
Hx11-1 (1.Wkb)
-1- - _-. GGlO-1 (6.6kb)
HX51 (1.41 kb) DTB-1 (1.05kb)
" - - - " HXl8-1 (0.92 kb) HXlB-1 (1.14kb)
" n x s 1 (0.74 kb) ""- Hx1&2(1.11 hb)
"""- HXll-2(0.Wkb) "- HX21-1 (.75Y)
013-1 (39 hb) - HxlBB1 (.53hb)
0
by adding 0.5 or 1.0 mM isopropyl-0-D-thiogalactopyranoside (IPTG), and the cell growth was continued for 3 h a t 30 "C. Cells were harvested by centrifugation a t 7,000 X g for 15 min and resuspended in 10 volumes of buffer containing 50 mM Tris-HC1, pH 7.5, 1 mM EDTA, 0.5 mM phenylmethylsulfonyl fluoride, 0.1 mM TLCK, 2 mM benzamidine, 10 pg/ml leupeptin, 50 mM 0-mercaptoethanol, and 1% Triton X-100. After freezing at -80 "C overnight, the cells were thawed and sonicated for 20 s twice. The lysate was centrifuged at 9,000 X g for 20 min. A sample of 1.4 p1 each of the whole cell lysate and the Triton-soluble fraction were analyzed by SDS-PAGE accord- ing to Laemmli (48) and by Western immunoblotting (49).
Western Blot Analysis-A synthetic peptide KPGFSPQPS- RRGSESSEEVYV surrounding the CAMP-dependent protein kinase phosphorylation site 1 on the Rcl subunit was synthesized and used to raise antibodies (anti-Rcl) in guinea pigs. The antibodies were affinity-purified on Sepharose 4B coupled to the peptide. Rabbit skeletal muscle was homogenized in 3 volumes of 50 mM Tris-HC1, p H 7.5, 1 mM EDTA, 1 mM EGTA, 0.5 mM phenylmethylsulfonyl fluoride, 2 mM benzamidine, 0.1 mM TLCK, 10 pg/ml leupeptin and centrifuged for 20 min a t 10,000 X g. A 1-pl sample of the rabbit skeletal muscle soluble extract and samples of E. coli extract prepared as described above were subjected to 7.5% SDS-PAGE. For immu- noblotting, gels were equilibrated for 20 min in 20 mM Tris, 190 mM glycine, 20% methanol, sandwiched with nitrocellulose membrane, and subjected to ice-cooled transverse electrophoresis a t 100 V for 2 h. The nitrocellulose filter was blocked overnight a t room temperature in 5% powder milk in PBS-T (20 mM sodium phosphate, pH 7.4, 115 mM sodium chloride, and 0.1% Tween 20) and then incubated for 2 h with anti-Rcl antibody (5 pg/ml in PBS-T). Bound antibodies were detected by incubating the filter for 1 h in PBS-T containing 0.2 &i/ ml of "'1-protein A. After removal of unreacted protein A the filter was subjected to autoradiography.
RESULTS
Isolation and Characterization of cDNA and Genomic DNA Clones-The initial screening of the rabbit skeletal muscle random hexamer-primed Xgtll cDNA library identified two positive clones that hybridized with both oligonucleotide probes G-COMP1 and G-COMP2. Nucleotide sequencing confirmed that clone HX 1-1 (995 bp) coded for the available amino acid sequences (50) of the R G ~ subunit polypeptide. This labeled cDNA fragment was used to rescreen the original filters and additional 160,000 plaque-forming units. Clones isolated from this screen were used for subsequent screening of another random hexamer-primed and an oligo(dT)-primed cDNA libraries. A total of 630,000 independent recombinants were screened, and 74 positive clones were identified, out of which 23 were characterized. Complete nucleotide sequences were determined from six clones (HX 1-1, 1-2, 5-1, 11-1, 13-
1 2 3 4kb
2, and DT 6-1), whereas partial sequences were obtained from other clones including HX 11-2, 15-1, 16-1,18-1, 18-2,18B-1, 21-1, and DT 3-1 (Fig. 1). These overlapping clones provided a combined sequence with an open reading frame of 3317 nucleotides and 537 nucleotides downstream of a stop codon. However, no in-frame ATG codon upstream of the known amino acid sequence was found. To search for the transla- tional start site, the HX 1-1 cDNA (995 bp), containing the most 5"sequences and a synthetic oligonucleotide from nu- cleotide 61 to 79, were used to screen a rabbit genomic library. Southern blot analysis of the single positive clone identified a 6.6-kb SphI genomic fragment which hybridized with the 995-bp cDNA fragment. Sequence analysis revealed that 108 bp of an intron were present at the 3'-end of the SphI fragment, which continued upstream in the coding region at nucleotide 799. By utilizing as primer an oligonucleotide cor- responding to nucleotide 61-79, sequences were extended at the 5'-end by 291 bp. In this region were several stop codons in all reading frames and no intronlexon boundary consensus sequence (51). Only two ATG codons were found, both in the same reading frame. The first was followed by an in-frame stop codon and the second was located 10 bp upstream from the cDNA 5'-end. The GCCCAATGG sequence around the second ATG is in reasonable agreement with the Kozak's consensus sequence (52). Thus, it seems likely that this ATG is the translational initiation codon. The COOH terminus is defined by a TAA codon which is followed by several other stop codons in all reading frames. No polyadenylation signal was found in the 537 bp of the 3"untranslated region. Se- quence data from all the clones establish an open reading frame of 3327 bases (Fig. 2) encoding a protein of 1109 amino acids with a M , of 124,257. The amino acid sequences of six peptides obtained from purified rabbit skeletal muscle RGI subunit, provided by Dr. Philip Cohen (University of Dundee, Dundee, Scotland), were present in the deduced sequences (Fig. 2) with only two mismatches, cysteine residues at posi- tions 26 and 183 for tyrosine and serine, respectively. Hy- dropathy analysis by the method of Rao and Argos (53) indicated a region, at the COOH terminus between residues 1063 ando 1097, rich in hydrophobic residues and which predicts a transmembrane helix.
Examination of the nucleotide sequences of all the cDNA clones characterized indicated the existence of two groups, clones HX 13-2, HX 1-2, HX 11-2, and HX 16-1 in one group and clones HX 1-1, HX 11-1, HX 5-1, and HX 15-1 in the

Protein Phosphatase 15785 CCAGAGAGCCCAATGGAGCCTTCTGMGTACCTGGTCAGAACAGCAAAGATMTTTTTTAGMGTTCCTMTTTGTCTGATTCTCTTTGTGMGATGMGMGTTMAGCTATTTTCMA 120
M E P S E V P G Q N S K D N F L E V P N L S D S 1 . C E D E E V K A T F K 36 CCTGGCTTCTCCCCTCMCCGAGCAGACGAGGTTCTGMTCTTCTGMGAGGTCTACGTGCACACCGCATCCTCAGGTG~AGMGAGTTTCGTTTGCTGACAACTTTGGATTCAATCTT 240
P G F S P O P S R R G S E S S . E E V Y V H T ~ F A D N F G F N ~ 7 6
GTGTCTGTGAAAGAGTTTGATACCTGGGMTTACCGAGTGTGTCCACCACCTTTGMTTAGGAMGGATGCTTTCCAGACAGMGMTATGTTTTATCTCCACTGTTTGACTTGCCTGCC 360 ~ S V K E F D T Y E L P S V S T T F E L G K D A F O T E ~ Y V I , S P I . F D I , P A 116 TCMAGGAAGATCTTATGCAACMCTACMGTTCAGAAAGCMTGCTGGAGTCMCTGAATATGTTCCT~TTCTACMGTATGMGGGTATTATTCGAG~TTGMTATTTCTTTTGAG 480
~ E D L ~ Q Q L Q V Q K A M L E S T E Y V P G S T S M K G I I R V L N I S F E 156 MGTTAGTATATGTGAGMTGTCCTTAGATGACTGGCAAACCCATTATGACATTTTGGCAGMTATGTTCCTMCTCGTGTGATGGTG~CCGATCAGTTCTCCTTCMGATTTCATTG 600
K L V Y V R ~ ~ L D D Y O T H Y D I L A E Y ~ P N ~ ~ D G E T D ~ F ~ F K I ~ L ~ ~ ~
V P P Y Q K D C S K V E F C I R Y E T S V G T F W S N N N G T N Y T L V C Q K K 236 GTTCCTCCTTATCAAAMGATGGCAGTAAAGTAMGTTGAGTTTTGTATACGTTACGAAACTTCTGTTGGTACATTTTGGTCMATMTMTGGCACAAATTATACACTGGTTTGTCAAAAGMG 720
GMCCTGAGCCAGAGCCTGGAAAGCCATTGGAAGMGCTCCTAGTAAACM~MGGCTGCTTGAAGGT~TCMGTMGGMGMTCATCAGMACATCAGAAGMMTAACTTT 840
GAAAATTCAAAGATTGCAGATACCTATATCCCMCAATTGTTTGTTCTCATGAGG~GGMGACTTGMMGCAGTTATCAAAATGT~GATGTAMCACGGAACATGATGMCAT 960 E P E P E P G K P L E E A P S K Q K K G C L K V K S S K E E S S E T S E E N N F 276
MTGAAlUAGMTTAGAGTTGATGATMATCAACGCTTMTMGMCTAGATGTGCTGCTTCTGMTATGG~GMCACATTGTCATCAGATCCATCCMTATTCCAAATAAACCTGAG 1080 E N S K I A D T Y I P T I V C S H E E K E D L K S S Y Q N V K D V N T E H D E H 316
GAGTTACAGMGMCCAAAGCCACAGTGM~ATGCACTGACTTGTCCCMAGGCTTTTGTCTCCAGGTTCATCAGCAGAAAGTTCCTTMAGGGAGATT~TACCACACTGMAAATAT 1200 N E K E L E L M I N Q R L I R T R C A A S E Y G K N T L S S D P S N I P N K P E 356
TCCTCAGGAAATGAGTCCAGTCATCAGCCTTCAGATATGGGAGAMTCMCCCCTCATTGGGAGGTACTACCAGTGATGGATCAGTGCMTTACACATCA~AGTMAGMATCCTGGAT 1320 E L Q K N Q S H S E A C T D L S Q R L L S P G S S A E S S L K G D F Y H T E K Y 396
GATAACGCTMTCCAGCCCATGGGAGTGGCAGAGGGGAAATATCTTGCTCCTTCCCAGGTCMCTTMGGCMGTMCCT~TAAAMGTATGAAGGAGGGGCTGAGMCAGTGAAATG 1440 S S G N E S S H Q P S D U G E I N P S L G G T T S D G S V Q L H I S S K E I L D 436
AAAGATTGCGMTGTTTACCMGAGATGTCCACTTG~GCATCGGACTATTTCA~ATCAACAG~CAGACCCTCCGAGGMGATTATGGGACTAGTAAGGATMTMGGAAAM 1560 D N A N P A H G S G R G E I S C S F P G Q L K A S N L N K K Y E G G A E N S E M 476
AGMTACAGTTAGATGTTGATGM~CMGCAAAMTTTTCGATCMTCTTCTATGACCMGAAAGAAATGTAGGCCACCTTGMATMCTGTGGMGGGATTGAAGCCAGTGACAGA 1680 K D C E C L P R D V H L K A S D Y F K K S T E N R P S E E D Y G T S K D N K E K 516
R I Q L D V D E K T S K N F R S I F Y D Q E R K V G H L E I T V E G I E A S D B 556 GACTTMCTTCTCTGCCMCCAAAGATACCACMTTCCMCTTGGGCMTCATGGMGATACATTTCACTCATCAAGGACACCTTTAGGTAGGGAAGMGCTGTGTTMCAACCCCAGM 1800 D L T S L P T K D T T T P T W A T M E D T F H S S R T P L G R E E A V L T T P E 596 CATGATCTCTCTTCTAGTGAAGGCACTATTTTAGGAGGGCTMCTGGAGGGGTTTGTTCACCAAGMATGGAAATGTTTTGMGMTGATTATCTTTTCCMGTTGA~AGAAAATCC 1920 H D L S S S E G T I L G G L T G G V C S P R N G N V L K B D Y L F O V E K R K S 636
GGTTGGATTMTCCTGMGATCAGMTAAGGACACACAGCATCAACAMGTTGGAATGTTCTGGAAAGTCAGGMAAAGCMGAGGGAGTAAGAC~TATAGCAGAGCAGATCAAAGM 2040 G W I N P E D Q N K D T Q H Q Q S W N V L E S Q E K A R G S K T N I A E Q I K E 676 CAAGTAGACTGTGMGACATGTGGGAAAAAAGAGATMCACAGGGAGTTTGAAAGCTACTCCTGCTGMGCATTGTTTACCTGCCMGMGCAGAGCACTGTGMCTGTCTCCTCTAGCT 2160 Q V D C E D M W E K R D N T G S L K A T P A E A L F T C Q E A E H C E L S P L A 716 GATCATGGTATTCCTGGGMAGCAGMGCAGGTACAGCTTATATMTTMGACMCATCAGMACTACCCCAGMAGCATGTCTGCTGGAG~AGCGATMTTGCTMGCTACCTCM 2280 D H C I P G K A E A G T A Y I I K T T S E T T P E S M S A G E K A I I A K L P Q 756
GAGACAGCGCGAAGTGACAGGCCCATGGAGGTAMGGAAACAGCGTTTGATCCACACGAAGGGAGMATGATGATTCACATTATACCCTTTGTCAACGAGACACAGTCGGTGTAATCGAT 2400 E T A R S D R P M E V K E T A F D P H E G R N D D S H Y T L C Q R D T V G V I D 796
GACAACGGTGTTGAAAAGGAATCCCATTTAGATATTTGTAATGCTCGTCTAGATGAAATGCGGMGGAGGAAGCCATGTCTATGCACAGCCCTGGGMGATGCGTGACAGGGAGAAACTC 2520 D N G V E K E S H L D I C N A R L D E M R K E E A M S U H S P G K M R D R E K L 836
GGCATTGGAMCATCACATCTGTAGMGMTCCTCACAGGTCATTGCAAATMTGMMAGCAACTTCMGGCTGGATTTACATTTGG~TGCCATCAGCAGACM~ATATTTCCA 2640 G I G N I T S V E E S S O V I A N N E K A T S R L D L H L E U P S A D K K I F P 876
GAAAATAGAGATCTTGGGCAGGTTCMGMTTATCAMGAAAACCGACATAGATAACACTGTTCACTCTGCCTTTMCTCAGACACGMCAGAGCTTCTCGGGATGACTCTCTTCTTTCC 2760 E N R D L G Q V Q E L S K K T D I D N T V H S A F N S D T N R A S R D D S L L S 916
AGTCACCATACAGAAACTTCAGTGCTGTCTTGTGMCAGGCAMTGCTGTAMGAATACAGTTACTACCACTGCTCTCCAGACTTCTGCTACTGAGTCAGMTATMTTGTAGTCCMCA 2880 S H H T E T S V L S C E Q A N A V K N T V T T T A L O T S A T E S E Y N C S P T 956
AGGGAAACCCAGGGTCMCCTGCATCTAAACCTGMGAAGTTTCCAGAGGTTCMGMGAGTGACATCAGAGACTAGGAAAGA~ATGCGTAGGCCAGATGTTCCAGTCAGGAGMTGC 3000 R E T Q G Q P A S K P E E V S R G S R R V T S E T R K E K C V G Q M F Q S G E C 996
MTGTGGAAATGTCGCMGGGCCAATGATTTTAGTCAGTGAATCTCGTGAGMCGTAGAMGGGAAAGGCATGAAAATGAAMTGAAGGACTGATTAACTCAGGTGACAAGGAATTTGAGAGCTCT 3120 N V E M S Q G P M I L V S E S R E N V E R E R H E N E G L I N S G D K E F E S S l O 3 6
GCTTCTTCTAGTCTACCTGTGCAGGAMCTCAAGATCAAAGCAATGAATCTCTTCTTTCAAMTACACCMCTCT~TACCTTATTTCCTTTTGTTTCTGATGTTTCTTGTAACCGTC 3240 A S S S L P V Q E T Q D Q S N E S L L S K Y T N S K I _ P Y F L L F J , " ~ J L V _ T - ~ 1 0 7 6
TACCACTATGACCTCATGATAGGCTTGGCATTCTACCTTTTCTCATTGTATTGGCTATACTGGGMGAGGGCAGACAAAMGAGTCTGTCA~GMGTMCCTCAGCACTATTATTAT 3360
TAAAAGATMGCTGTTTAGCTCCAMCATTTGGATTGGTGMGGAGACTATTCATTGTTCAAAGAGCCAGTGCAGTTTTTCTCTTGAAGGATCATTTMAAAGGAATGCCTATGMGTTT 3480 1109
GCTCCTTCATATMGTMTTATTCTATATAGGACCATTATGTTTGGATCATTAAATACCTATATGMTATGAGATCTGAAGCACGTCMGTTGMATTAGGTACAGCTGTTGCTCCTTAG 3600 CAGGCTATGAAGTTGCMTGCTTCACGTCTCTTCACTACTT~GTGCTATTTCTTGTGTTCATTTCTTTTGCAGTAAAGCTTCATTTTTTTCCCCCTGAGCACATCTTTCCCTCTATGG 3720 TTTTTAAAMTAGATAMACATGGACMTGGCAGMGATTTTCTTCCTTTTTTTTTGTCTTTTAGGATTGACMTGAAATTTTCATCTACCACTGTATCATTTATTAGCACATAATGATA 3840 GATCAACTATTTCAACTCATATTTCATAGTTTTAAG 3876
~ H Y D L M I G L A F Y L F S L Y W L Y W E E G R Q K E S V K K K ~
FIG. 2. Nucleotide and deduced amino acid sequences of rabbit skeletal muscle Rcl subunit. The complete sequence was obtained by combining all the clones shown in Fig, 1. A single underline indicates peptide sequences derived from the RGI subunit polypeptide and a double underline the amino acid sequences used for synthesis of the oligonucleotide probes employed in the screening. The dotted l ine indicates the region rich in hydrophobic residues.
second. The two groups differed in six nucleotides, at positions 703, 744, 1176, 1302, 944, and 1251. The last two caused changes in amino acid sequences at residue 311 from threo- nine to methionine and at residue 413 from asparagine to lysine, respectively. The other four differences were silent. Most likely this discrepancy is due to allelic variations.
Determination of the Transcriptional Start Site-Primer extension analysis was carried out in order to map the 5'-end of the rabbit Rcl subunit transcript. Utilizing total RNA prepared from rabbit skeletal muscle, a major transcriptional start point at a C located 12 nucleotides upstream from the translational start codon ATG was identified (Fig. 3). This putative cap site was observed also in RNA prepared from rabbit diaphragm and cardiac muscle but not from lung (data
not shown) in which no homologous RGl subunit appears to be expressed.
Tissue Distribution of the Rcl Subunit mRNA and Southern Analysis of Genomic DNA-The tissue distribution and the complexity of the Rcl subunit mRNA was investigated by Northern analyses. Total RNA from various rabbit tissues was hybridized with the labeled cDNA inserts from clone HX 1-1 (995 bp) and from clone HX 1-2 (2.95 bp) as probes. A major hybridizing mRNA species at 7.5 kb was observed in skeletal, diaphragm, and cardiac muscle (Fig. 4). Minor spe- cies of approximately 3.5 kb were also observed at low strin- gency. The level of the mRNA appeared to be higher in skeletal than in cardiac muscle. However, none of the mRNA species was present in brain, kidney, liver, and lung, although staining of the gel with ethidium bromide indicated that
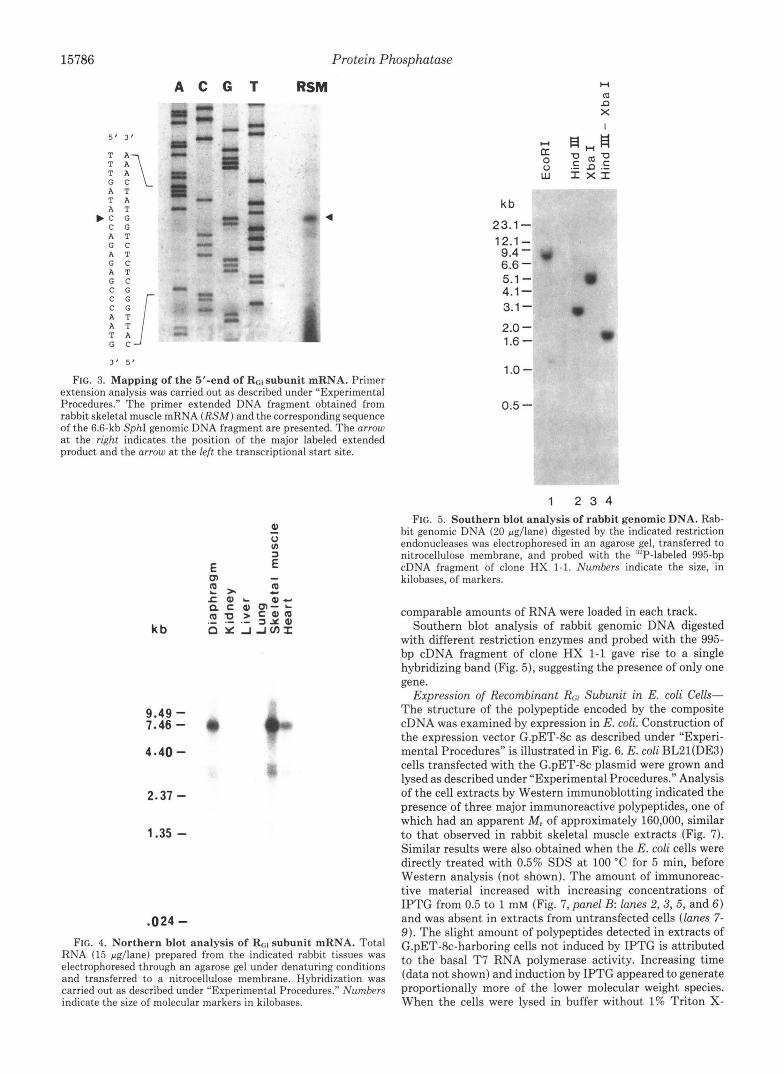
15786 Protein Phosphatase
A C G T RSM ” -
5’ 3’
T A
G C A T T A
b c G A T
C G A T G C A T G C A T G C
A T T A G C “?r 3 ‘ 5’
FIG. 3. Mapping of the 5’-end of Rcl subunit mRNA. Primer extension analysis was carried out as described under “Experimental Procedures.” The primer extended DNA fragment obtained from rabbit skeletal muscle mRNA ( R S M ) and the corresponding sequence of the 6.6-kb SphI genomic DNA fragment are presented. The arrow at the right indicates the position of the major labeled extended product and the arrow at the left the transcriptional start site.
kb
9.49 - 7.46 - 4-40 -
2.37 -
1.35 -
.024 -
Q Y J,rcnI
FIG. 4. Northern blot analysis of RGl subunit mRNA. Total RNA (15 &lane) prepared from the indicated rabbit tissues was electrophoresed through an agarose gel under denaturing conditions and transferred to a nitrocellulose membrane. Hybridization was carried out as described under “Experimental Procedures.” Numbers indicate the size of molecular markers in kilobases.
kb
23.1 - 12.1 - 9.4 - 6.6 - 5.1 - 4.1 - 3.1 - 2.0 - 1.6 - 1.0 -
0.5 -
1 2 3 4 FIG. 5. Southern blot analysis of rabbit genomic DNA. Rab-
bit genomic DNA (20 pgllane) digested by the indicated restriction endonucleases was electrophoresed in an agarose gel, transferred to nitrocellulose membrane, and probed with the ‘lP-labeled 995-bp cDNA fragment of clone HX 1-1. Numbers indicate the size, in kilobases. of markers.
comparable amounts of RNA were loaded in each track. Southern blot analysis of rabbit genomic DNA digested
with different restriction enzymes and probed with the 995- bp cDNA fragment of clone HX 1-1 gave rise to a single hybridizing band (Fig. 5), suggesting the presence of only one gene.
Expression of Recombinant Rc, Subunit in E. coli Cells- The structure of the polypeptide encoded by the composite cDNA was examined by expression in E. coli. Construction of the expression vector G.pET-8c as described under “Experi- mental Procedures” is illustrated in Fig. 6. E. coli BL21(DE3) cells transfected with the G.pET-8c plasmid were grown and lysed as described under “Experimental Procedures.” Analysis of the cell extracts by Western immunoblotting indicated the presence of three major immunoreactive polypeptides, one of which had an apparent M , of approximately 160,000, similar to that observed in rabbit skeletal muscle extracts (Fig. 7). Similar results were also obtained when the E. coli cells were directly treated with 0.5% SDS at 100 “C for 5 min, before Western analysis (not shown). The amount of immunoreac- tive material increased with increasing concentrations of IPTG from 0.5 to 1 mM (Fig. 7, panel B: lunes 2 , 3 , 5 , and 6 ) and was absent in extracts from untransfected cells (lunes 7- 9). The slight amount of polypeptides detected in extracts of G.pET-8c-harboring cells not induced by IPTG is attributed to the basal T7 RNA polymerase activity. Increasing time (data not shown) and induction by IPTG appeared to generate proportionally more of the lower molecular weight species. When the cells were lysed in buffer without 1% Triton X-
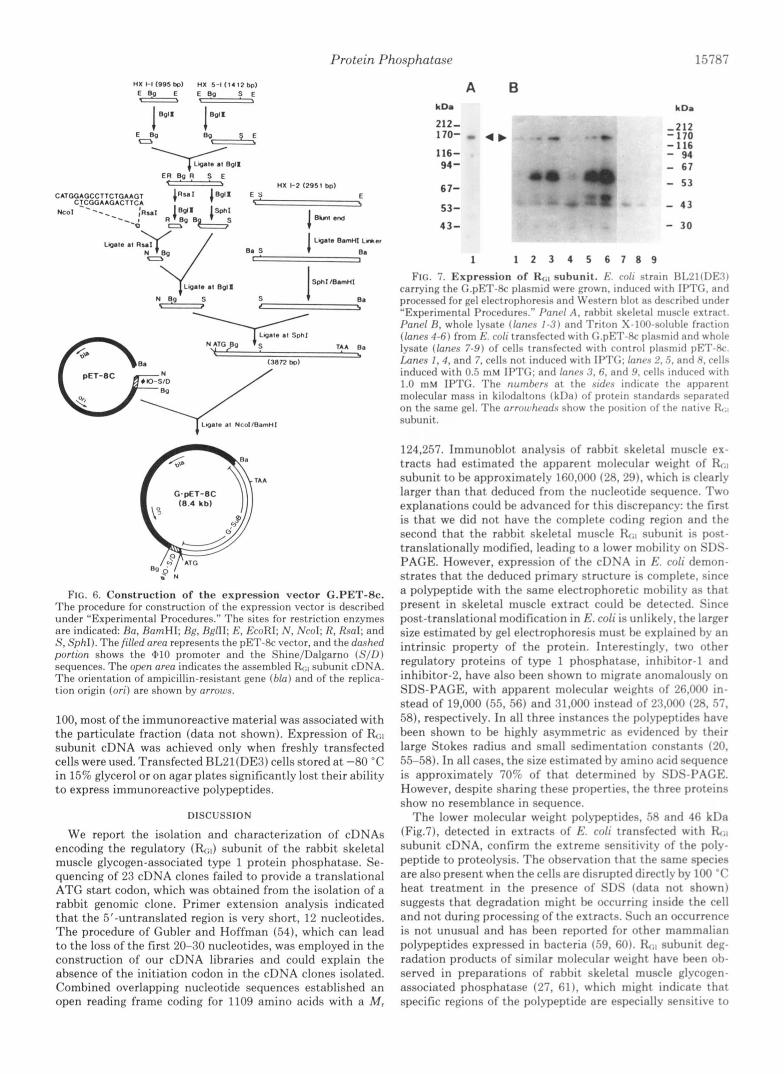
Protein Phosphatase 15787
E R B g k S E -
(8.4 kbl (8.4 kbl
T I A
FIG. 6. Construction of the expression vector G.PET-8c. The procedure for construction of the expression vector is descrihed under “Experimental I’rocedures.” The sites for restriction enzymes are indicated: Ro, RamHI; Hp, RgflI; E, E~cJRI; N , Ncol: R, Rsnl; and S , Sphl). The filled oren represents the PET-8c vector, and the dashrd portton shows the $110 promoter and the Shine/Dalgarno ( S / I ) ) sequences. The open own indicates t,he assemhled R;l suhunit cDNA. The orientation of ampicillin-resistant gene (bln) and of the replica- tion origin (or i ) are shown hy nrrours.
100, most of the immunoreactive material was associated with the particulate fraction (data not shown). Expression of E;, subunit cDNA was achieved only when freshly transfected cells were used. Transfected RL21(DE3) cells stored at -80 “C in 15% glycerol or on agar plates significantly lost their ability to express immunoreactive polypeptides.
DISCUSSION
We report the isolation and characterization of cDNAs encoding the regulatory (E;,) suhunit of the rahhit skeletal muscle glycogen-associated type 1 protein phosphatase. Se- quencing of 23 cDNA clones failed to provide a translational ATG start codon, which was obtained from the isolation of a rabbit genomic clone. Primer extension analysis indicated that the 5’-untranslated region is very short, 12 nucleotides. The procedure of Gubler and Hoffman (54), which can lead t o t h e loss of the f irst 20-30 nucleotides, was employed in the construction of our cDNA libraries and could explain the absence of the initiation codon in the cDNA clones isolated. Combined overlapping nucleotide sequences established an open reading frame coding for 1109 amino acids with a M,
A B kD8 kDm
212- 170- - 4 b 116- 94-
67-
53-
43-
- 170 -212
- 116 - 94 - 67
- 53
- 4 3
- 30
1 1 2 3 4 5 6 7 0 9
FIG. 7. Expression of RCil subunit. E , ro/i strain F 3 1 2 1 ~ D F 3 ~ carrying the (;.pE’l‘-Hc plasmid were grown. induced with II’TG, and processed for gel electrophoresis and li’estern hlnt as descril)etl under “Experimental Procedures.” I’anrl A , rahhit skeletal muscle extract. P o n d H. whole lysate (lnnrs 1 - 3 ) and Tri ton X-lf~O-soluhle fraction (lanes 4 - 6 ) from b;. coli transfected with G.pF:T-Rc plasmid and whole lysate (lnnrs 7-9) of cells transfected with control plasmid p F T - H c . Imnrs I , 4 , and 7. cells not induced with IPTG; /ones 2. 5 , and 8. cells induced with 0.5 mM II’TG: antl /nnrs 3, 6. antl 9. cells intfuced w i t h 1.0 mM II’TG. The numbers at the sidrs indicate the apparent molecular mass in kiloclaltons (kI)a) of protein standards separatc.d on the same gel. The arrowhcods show t h e position of t hP native It<,) suhunit.
124,257. lmmunohlot analysis of rahhit skeletal muscle ex- tracts had estimated the apparent molecular weight of R; l suhunit to he approximatelv 160,000 (28, 29), which is clearly larger than that deduced from the nucleotide sequence. Two explanations could he advanced for this discrepancy: the first is tha t we did not have the complete coding region and the second that the rahhit skeletal muscle &;I subunit is post- translationally modified, leading to a lower mohility on SDS- PAGE. However, expression of the cDNA in E . coli demon- strates that the deduced primary structure is complete, since a polypeptide with the same electrophoretic mohilitv as that present in skeletal muscle extract could he detected. Since post-translational modification in E. coli is unlikely, the larger size estimated by gel electrophoresis must he explained bv an intrinsic property of the protein. Interestinglv, two other regulatory proteins of t-ype 1 phosphatase, inhihitor-1 and inhihitor-2, have also heen shown to migrate anomalouslv on SDS-PAGE, with apparent molecular weights of 26.000 in- stead of 19,000 (55, 56) and 31,000 instead of 23,000 (28. 57, 58), respectively. In all three instances the pol-ypeptides have heen shown to he highlv asymmetric as evidenced hv their large Stokes radius and small sedimentation constants ( 2 0 . 55-58). In all cases, the size estimated hv amino acid sequence is approximately 70% of that determined hv SI>S-I’AGF:. However, despite sharing these properties. the three proteins show no resernhlance in sequence.
T h e lower molecular weight polypeptides, 58 and 46 kDa (Fig.7), detected in extracts of E . coli transfected with suhunit cDNA, confirm the extreme sensitivitv of the polv- peptide to proteolysis. The observation that the same species are also present when the cells are disrupted directly hy 100 “ C heat treatment in the presence of SIX (data not shown) suggests that degradation might he occurring inside the cell and not during processing of the extracts. Such an occurrence is not unusual and has heen reported for other mammalian polypeptides expressed in hacteria (.59, 60). RiI suhunit deg- radation products of similar molecular weight hnve heen oh- served in preparations of rahhit skeletal muscle glvcogen- associated phosphatase (27, 61), which might indicate that specific regions of the pol-ypeptide are especiallv sensitive to

15788 Protein Phosphatase
proteolytic cleavage. We can exclude that the 58- and 46-kDa forms are generated by initiation of translation at a down- stream ATG, since the antibodies used for the detection were raised to the region corresponding to residues 37 to 56 and no other ATGs are present in this NHz-terminal region.
Recently Hubbard et al. (22) reported that a sarcoplasmic reticulum-associated phosphatase contains a polypeptide sim- ilar, if not identical, to the RG, subunit. Hydrophathy analysis of the deduced amino acid sequence reported here indicated a potential transmembrane region between residues 1063 and 1097 (Fig. 2), which could be responsible for anchoring the protein to the membrane, Thus, the same regulatory subunit might function to target the phosphatase to membranes and glycogen. Several lines of evidence argue against the existence of distinct muscle isoforms. The low molecular weight poly- peptides observed in glycogen-associated phosphatase purified from rabbit skeletal muscle are not always detected by West- ern immunoblotting analysis (Fig. 7) and Northern hybridi- zation, utilizing 3,850 bp of cDNA sequence, indicated one major mRNA species, The minor differences observed in the two groups of cDNA clones, four silent nucleotide substitu- tions and two changing the amino acid residues could be explained by allelic variations. Southern analysis also sug- gested a single gene. Studies in progress, with mutant protein in which the hydrophobic region has been deleted, should prove useful in addressing the question of how the same polypeptide might be directed to different cellular compart- ments.
Analysis of the tissue distribution of the RG, subunit mRNA (Fig. 4) supports previous observations by Western immuno- blot (28); which indicated that the polypeptide is specifically expressed in skeletal and cardiac muscle, but not in other tissues examined. This also suggests that the polypeptide responsible for targeting the phosphatase to glycogen in liver is not homologous to the muscle form, although their molec- ular weights appear to be very similar and both interact with a highly conserved catalytic subunit (62).
The rabbit skeletal muscle glycogen-associated phosphatase undergoes in vivo and in vitro phosphorylation at several sites all of which are located near the NH2 terminus. The CAMP- dependent protein kinase sites are at residues 48 and 67, respectively, and the glycogen synthase kinase-3 sites at po- sitions 40 and 44. Other potential recognition sites for CAMP- dependent protein kinase are threonine 498 and 978 and serine 636. Since purification of intact R c l subunit has been difficult and some of the site identifications have been carried out on proteolyzed species (50), it is possible that additional phosphorylation sites might have been missed. Threonine 978, in the motif -Arg-Arg-Val-Thr-, is an especially strong can- didate. Phosphorylation of this residue, similarly to serine 48, could form a recognition site for glycogen synthase kinase-3 (31).
Search of the Swiss protein (release 13) and the EMBL (release 21) data bases with PCGENE utilizing the FASTN and FASTP programs (63) revealed no significant homology between the Rc, subunit and other known sequences. How- ever, a search of the protein data base assembled by Dr. Mark Goebl, at Indiana University, indicated significant homology with the product of the yeast gene, GACl, isolated by Dr. Kelly Tatchell at North Carolina State Uni~ersity.~ Over a segment of 144 residues, the identity is 27% and the homology
' K. M. Swiderek and A. A. DePaoli-Roach, unpublished results. K. Tatchell, personal communication.
38% if conservative replacements are taken into acc~un t .~ Utilizing the algorithm of Lipman and Pearson (63), the optimal alignment score between R G 1 and GACl amino acid sequence is 13 standard deviations over the mean of the optimal score of 100 random shufflings of the GAG1 sequence. It is of significance that the similarity lies within the 40-kDa NHz-terminal portion of the protein, which is able to interact with glycogen and with the catalytic subunit of type 1 phosphatase (61). The GACl protein appears to be involved in activation of glycogen synthase and glycogen accumulation. Both of these functions are consistent with the GACl gene product being the yeast homologue of the R G ~ subunit. Gene replacement in yeast should allow us to address this question. In addition site specific and deletion mutagenesis will provide a powerful tool to elucidate the physiological role and regu- lation of the glycogen-associated protein phosphatase.
Acknoufedgments-We are grateful to Dr. P. Cohen for making available amino acid sequences from RGI subunit peptides, to Dr. F. W. Studier for the generous gift of the T7 RNA polymerase expression system, to Drs. R. C. Hardison and T. Maniatis for the rabbit genomic library, to Dr. K. Tatchell for allowing comparative analysis with the GACl gene product, and to Dr. M. Goebl for carrying out the analysis. We also thank Dr. P. J. Roach for his advice and criticism in t,he preparation of the manuscript.
I.
2. 3.
4. 5. 6.
7. 8. 9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19. 20.
21.
22.
23.
REFERENCES Edelman, A. M., Blumenthal, D. K., and Krebs, E. G. (1987)
Annu. Rev. Biochem. 56,567-613 Hunter, T. (1987) Cell 50,823-829 Stull, J. T., Nunnally, M. H., and Michnoff, C. H. (1986)in The
Enzymes (Boyer, P. D.) 3rd Ed., Voi. 17, pp. 114-166, Academic Press, Orlando, FL
Taylor, S. S. (1989) J. Biol. Chem. 264,8443-8446 Soderling, T. R. (1990) J. Biol. Chem. 265,1823-1826 Kemp, B. E., and Pearson, R. B. (1990) Trends Biochem. Sei.
Ingebritsen, T. S., and Cohen, P. (1983) Science 221,331-338 Cohen, P. (1989) Annu. Rev. Biochem. 58,453-508 Cohen, P., and Cohen, P. T. W. (1989) J . Biol. Chem. 2 6 4 ,
21435-21438 Berndt, N., Campbell, D. G., Caudwell, F. B., Cohen, P., da Cruz
e Silva, E. F., da Cruz e Silva, 0. B., and Cohen, P. T. W. (1987) FEBS Lett. 223,340-346
Bai, G., Zhang, Z., Amin, J., Deans-Zirattu, S. A., and Lee, E. Y.
Arino, J., Woon, C. W., Brautigan, D. L., Miller, Y. B. Jr., and Johnson, G. L. (1988) Proc. Natl. Acad. Sci. U. S. A. 85,4252- 4256
Green, D. D., Yang, %-I., and Mumby, M. C. (1987) Proc. Natl. Acad. Sci. U. S. A. 84,4880-4884
Da Cruz e Silva, 0. B., Alemany, S., Campbell, D. G., and Cohen,
Kincaid, R. L., Nightingale, M. S., and Martin, B. M. (1988) Proc. Natl. Acad. Sci. U. S. A. 85,8983-8987
Tamura, S., Lynch, K. R., Larner, J., Fox, J., Yasui, A., Kikuchi, K., Suzuki, Y., and Tsuiki, S. (1989) Proc. Natl. Acad. Sci.
Yang, S. D., Vandenheede, J. R., Goris, J., and Merievede, W.
Ballou, L. M., Brautigan, D. L., andFisher, E. H. (1983) Biochem-
DePaoli-Roach, A. A. (1985) Adu. Prot. Phosphatases 1,59- 72 Stralfors, P.. Hirana. A., and Cohen, P. (1985) Eur. J. Biochem.
177,342-346
C. (1988) FASEB J. 2,3010-3016
P. T. W. (1987) FEBS Lett. 221,415-422
U. S. A. 86, 1796-1800
(1980) J . Bwl. Chem. 2 5 5 , 11759-11767
istry 22,3393-3399
I . . 149,295-303
Villa-Moruzzi. E.. and Heilmever, L. M. G.. Jr. (1987) Eur. J. - . ~. Bwchem. 169,659-667
J. Biochem. 189,243-249 Hubbard, M. J., Dent, P., Smythe, C., and Cohen, P. (1990) Eur.
Chisholm, A. A. K., and Cohen, P. (1988) Biochim. Biophys. Acta 968,392-400
The following groupings of amino acids were considered homolo- gous: Val-Ile-Leu; Phe-Tyr; Ala-Ser-Thr; Asp-Glu; Asn-Gin; Arg- Lys.

Protein Phosphatase 15789
24. Schlender, K. K., Wang, W., and Wilson, S. E. (1989) Biochem. 43. Chirgwin, J. M., Przybyla, A. E., MacDonald, R. J., and Rutter,
25. Kuret, J., Bell, H., and Cohen, P. (1986) FEBS Lett. 203, 197- 44. Thomas, P. S. (1980) Proc. Natl. Acad. Sci. U. S. A. 77, 5201-
26. Jakes, S., Merlgren, R. L., and Schlender, K. K. (1986) Biochim. 45. Rigby, P. W., Dieckmann, M., Rhodes, C., and Berg, P. (1977) J.
27. Hiraga, A,, and Cohen, P. (1986) Eur. J. Biochm. 161, 763-769 46. Studier, F. W., and Moffatt, B. A. (1986) J. Mol. Biol. 189, 113- 28. DePaoli-Roach, A. A. (1989) Adu. Protein Phosphatases 5, 479- 130
500 47. Studier, F. W., Rosenberg, A. H., Dunn, J. J., and Dubendorff, J. 29. Hubbard, M. J., and Cohen, P. (1989) Eur. J . Biochem. 180, W. (1990) Methods Enzymol. 185, 60-89
30. Hubbard, M. J., and Cohen, P. (1989) Eur. J . Biochem. 186, 49. Burnette, W. N. (1981) Anal. Biochem. 112, 195-203
31. Fiol, C. J., Haseman, J. H., Wang, Y., Roach, P. J., Roeske, R. 194,85-90
Biophys. Res. Commun. 159, 72-78 W. J. (1979) Biochemistry 18, 5294-5299
202 5205
Biophys. Acta 888, 135-142 Mol. Biol. 113, 237-251
457-465 48. Laemmli, U. K. (1970) Nature 227, 680-685
701-709 50. Caudwell, F. B., Hiraga, A,, and Cohen, P. (1986) FEBS Lett.
W., Kowalczuk, M., and DePaoli-Roach, A. A. (1988) Arch. 51. Nevins, J. R. (1983) Annu. Reu. Biochem. 52,441-466 Biochem. Biophys. 267, 797-802 52. Kozak, M. (1987) J. Mol. Biol. 196, 947-950
32. Dent, P., Campbell, D. G., Hubbard, M. J., and Cohen, P. (1989) 53. Rao, M. J. K., and Argos, P. (1986) Biochim. Biophys. Acta 869, FEBS Lett. 248,67-72 197-214
33. Dent, P., Campbell, D. G., Caudwell, F. B., and Cohen, P. (1990) 54. Gubler, U., and Hoffman, B. J. (1983) Gene (Amst.) 25, 263-269 FEBS Lett. 259,281-285 55. Nimmo, G. A., and Cohen, P. (1978) Eur. J. Biochem. 87, 341-
34. Dent, P., Lavoinne, A., Nakielny, S., Caudwell, F. B., Watt, P., 351 and Cohen, P. (1990) Nature 348, 302-307 56. Aitken, A., Bilham, T., and Cohen, P. (1982) Eur. J . Biochem.
35. Zhang, W., Browner, M. F., Fletterick, R. J., DePaoli-Roach, A. 126,235-246 A., and Roach, P. J. (1989) FASEB J. 3, 2532-2536 57. Holmes, C. F. B., Campbell, D. G., Caudwell, F. B., Aitken, A.,
36. Maruyama, T., Gojobori, T., Aota, A.-I., and Ikemura, T. (1986) and Cohen, P. (1986) Eur. J. Biochem. 155, 173-182 Nucleic Acids Res. 14, (suppl.) 151-197 58. Foulkes, J. G., and Cohen, P. (1980) Eur. J . Biochem. 105, 195-
37. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular 203 cloning: A laboratory Manual, Cold Spring Harbor Laboratory, 59. Goeddel, D. V., Heyneker, H. L., Hozumi, T., Arentzen, R., Cold Spring Harbor, NY Itakura, K., Yansura, D. G., Ross, M. J., Mizzari, G., Crea, R.,
38. Messing, J. (1983) Methods Enzymol. 101, 20-78 and Seeburg, P. H. (1979) Nature 281, 544-548 39. Feinberg, A. P., and Vogelstein, B. (1983) Anal. Biochem 132, 60. Goeddel, D. V. (1990) Methods Enzymol. 185, 3-7
40. Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl. 163,253-258
41. Maniatis, T., Hardison, R. C., Lacy, E., Lauer, J., O’Connel, C., 266,339-345
42. Southern, E. M. (1975) J. Mol. Biol. 98, 503-517 1441
6-13 61. Hiraga, A., Kemp, B. E., and Cohen, P. (1987) Eur. J . Biochem.
Acad. Sci. U. S. A. 74,5463-5467 62. Wera, S., Bollen, M., and Stalmans, W. (1991) J. Biol. Chem.
and Quon, D. (1978) Cell 15, 687-701 63. Lipman, D. J., and Pearson, W. R. (1985) Science 227, 1435-