neuronal morphology in the african elephant loxodonta
TRANSCRIPT

ORIGINAL ARTICLE
Neuronal morphology in the African elephant(Loxodonta africana) neocortex
Bob Jacobs • Jessica Lubs • Markus Hannan •
Kaeley Anderson • Camilla Butti • Chet C. Sherwood •
Patrick R. Hof • Paul R. Manger
Received: 14 August 2010 / Accepted: 15 October 2010 / Published online: 16 November 2010
� Springer-Verlag 2010
Abstract Virtually nothing is known about the mor-
phology of cortical neurons in the elephant. To this end, the
current study provides the first documentation of neuronal
morphology in frontal and occipital regions of the African
elephant (Loxodonta africana). Cortical tissue from the
perfusion-fixed brains of two free-ranging African ele-
phants was stained with a modified Golgi technique.
Neurons of different types (N = 75), with a focus on
superficial (i.e., layers II–III) pyramidal neurons, were
quantified on a computer-assisted microscopy system using
Neurolucida software. Qualitatively, elephant neocortex
exhibited large, complex spiny neurons, many of which
differed in morphology/orientation from typical primate
and rodent pyramidal neurons. Elephant cortex exhibited a
V-shaped arrangement of bifurcating apical dendritic
bundles. Quantitatively, the dendrites of superficial pyra-
midal neurons in elephant frontal cortex were more com-
plex than in occipital cortex. In comparison to human
supragranular pyramidal neurons, elephant superficial
pyramidal neurons exhibited similar overall basilar den-
dritic length, but the dendritic segments tended to be longer
in the elephant with less intricate branching. Finally, ele-
phant aspiny interneurons appeared to be morphologically
consistent with other eutherian mammals. The current
results thus elaborate on the evolutionary roots of Afro-
therian brain organization and highlight unique aspects of
neural architecture in elephants.
Keywords Dendrite � Morphometry � Dendritic spine �Golgi method � Brain evolution
Introduction
In recent years, elephants have received considerable
attention because of their complex social structure and
cognitive abilities (Poole and Moss 2008; Hart and Hart, in
press). Nevertheless, despite elephants’ status as the largest
living terrestrial mammal, very little is known about the
brain of this species. Prior to 2001, only a few original
articles had specifically focused on the elephant central
nervous system (Cozzi et al. 2001). Although the gross
anatomy of the *5 kg adult elephant brain has been
investigated in the last decade (Kupsky et al. 2001; Hakeem
et al. 2005, 2009; Shoshani et al. 2006; Manger et al. 2010),
virtually nothing is known about the microstructural mor-
phology of its neurons. Indeed, until recently, the only
apparent insight into elephant cortical neuromorphology
was a single camera lucida drawing from an Indian elephant
(Elephas maximus; Barasa and Shochatovitz 1961). Rea-
sons for this poverty of data are many (Cozzi et al. 2001),
but include a propensity for neuroscientists to focus on the
rodent and primate species commonly used in biomedical
B. Jacobs (&) � J. Lubs � M. Hannan � K. Anderson
Laboratory of Quantitative Neuromorphology, Psychology,
The Colorado College, 14 E. Cache La Poudre,
Colorado Springs, CO 80903, USA
e-mail: [email protected]
C. Butti � P. R. Hof
Department of Neuroscience and Friedman Brain Institute,
Mount Sinai School of Medicine, One Gustave L. Levy Place,
New York, NY 10029, USA
C. C. Sherwood
Department of Anthropology, The George Washington
University, 2110 G Street, NW, Washington, DC 20052, USA
P. R. Manger
Faculty of Health Sciences, School of Anatomical Sciences,
University of the Witwatersrand, 7 York Road, Parktown,
Johannesburg 2193, South Africa
123
Brain Struct Funct (2011) 215:273–298
DOI 10.1007/s00429-010-0288-3

research (Manger et al. 2008), and a lack of well-preserved
elephant brain tissue suitable for histological analysis. This
latter issue was recently addressed by Manger et al. (2009),
who were able to fix the brains of wild African elephants
(Loxodonta africana) by postmortem, carotid cannulation-
perfusion. The quality of this tissue facilitated the present
investigation, which qualitatively and quantitatively
explores, for the first time, the neuromorphology of frontal
and occipital cortices in the African elephant.
Elephants belong to the order Proboscidea, which
emerged approximately 60 million years ago from within
the eutherian superorder Afrotheria (Sukumar 2003;
Shoshani and Tassy 2005; Gheerbrant and Tassy 2009;
Fig. 1). Extant afrotherian species (e.g., elephants, mana-
tees, dugongs, hyraxes, aardvarks, golden moles, tenrecs,
and elephant shrews) exhibit significant diversity in terms of
body and brain mass, neural organization, and life history
(Gravett et al. 2009; Pieters et al. 2010). Because the afro-
therian clade represents a major adaptive radiation within
Eutheria, it is in a position to provide insight into mamma-
lian brain evolution, with current evidence suggesting that
Afrotheria contains many neural plesiomorphies in com-
parison with other placental mammal species (Sherwood
et al. 2009). Thus, examination of neuronal morphology in
elephants is essential for understanding broader phyloge-
netic patterns of neuromorphological evolution in Afrothe-
rians and provides data concerning the scaling of neuronal
somatodendritic geometry with increasing body and brain
size (Tower 1954; Haug 1987; Hart et al. 2008).
Both cognitive and sociobehavioral investigations of
elephants reinforce the view that such a large brain is
associated with extensive abilities (Roth and Dicke 2005;
Hart and Hart, in press). The cognitive repertoire of ele-
phants includes elementary tool construction/usage (Hart
et al. 2001), impressive spatial and temporal memory
(Markowitz et al. 1975; Hart et al. 2008), creative problem
solving skills (Bates et al. 2008b), the ability to classify
human ethnic groups by odor and garment color (Bates
et al. 2007a), and the ability to locate out-of-site family
members by smell (Bates et al. 2007b). Social interactions
in elephants are underscored by several documented abil-
ities: ultra-low frequency sound communication over both
local and long distances (Poole 1999; Garstang 2004; Soltis
2009), vocal learning (Poole et al. 2005), long-term care of
their young (Bates et al. 2008b), individual identification of
conspecifics (Bates et al. 2007b), and targeted helping
behavior (Hart et al. 2008). These abilities suggest ele-
phants may be capable of empathy (Bates et al. 2008a, b)
and self-recognition, including theory of mind (Plotnik
et al. 2006), all of which crucially contribute to their
complex social networks (McComb et al. 2000, 2001).
Unfortunately, besides the recent documentation of von
Economo neurons in the anterior insular cortex of elephants
(Hakeem et al. 2009), one can only extrapolate the neural
foundations for these cognitively demanding abilities. For
example, based on the low density of neurons in their
cerebral cortex, it has been suggested that the elephants’
cognitive faculties may be concentrated in long-term pro-
cessing (Hart and Hart 2007; Hart et al. 2008), although fine
sensorimotor integration is certainly required to guide the
trunk (Hoffmann et al. 2004). Apart from such speculation,
there are currently no data that allow for direct examination
of the computational biophysics of elephant cortical neu-
rons. However, in the prototherian echidna, whose ancestry
is close to the root of the mammalian phylogenetic tree
(Fig. 1), a large proportion of pyramidal neurons contain so-
called ‘‘atypical’’ features, such as bifurcated apical den-
drites, inverted somata, poorly developed basilar skirts, and
a lack of terminal bouquets in layer I (Hassiotis et al. 2003,
2005). This suggests that there may be significant phyloge-
netic diversity in the morphology of spiny projection neu-
rons of the cerebral cortex. As such, based on its afrotherian
origin, we expect a variety of spiny neuron morphologies in
the elephant that differs from the euarchontoglire branch of
Fig. 1 Taxonomy of the elephant (Loxodonta africana) and a
sampling of related species mentioned in the present paper. During
the late Cenozoic era, Eutheria (placental mammals) diverged into
two phylogenetic groups: Atlantogenata and Boreoeutheria (Wildman
et al. 2007). Atlantogenata subsequently diverged into two sister
clades, Xenarthra (e.g., sloths, anteaters, and armadillos) and
Afrotheria, whereas Boroeutheria diverged into Laurasiatheria (e.g.,
cetaceans, carnivores, and ungulates) and Euarchontoglires (e.g.,
primates and rodents)
274 Brain Struct Funct (2011) 215:273–298
123

placental mammals, mostly typified by anthropoid primate
and murid rodent models, and thus appear ‘‘atypical’’. In
contrast, the morphological classes for aspiny interneurons
are more uniform across eutherian mammals (Hof et al.
1996, 1999; DeFelipe et al. 2002; Hassiotis and Ashwell
2003; Hof and Sherwood 2005). Thus, we expect aspiny
interneurons in elephants to resemble more closely the types
that are common in the mammalian cerebral cortex.
By extension, neuromorphological diversity in the ele-
phant implies a diverse cytoarchitecture, which would
contradict the long-held, but problematic claim that large
brains—such as in cetaceans—lack histological complexity
(Kesarev 1975; Glezer et al. 1988). In fact, recent studies on
cetacean cerebral cortices have revealed a complex cytoar-
chitecture, comparable to that observed in many mammals,
including primates (Hof et al. 2005; Hof and Van der Gucht
2007; Butti et al. 2009; Butti and Hof 2010). The present
study examines such neuromorphological diversity by
exploring regional variation in neuron morphologies across
all layers of the elephant cortex and by examining quanti-
tative differences in superficial (i.e., layers II–III) pyramidal
neurons between frontal and occipital cortical regions. As is
the case with human supragranular pyramidal neurons, these
superficial pyramidal neurons also tended to stain clearly
with the rapid Golgi technique (Jacobs et al. 2001). Both
functional (Foxe and Simpson 2002) and morphological
data in primates (Elston and Rosa 1997; Jacobs et al. 2001)
suggest regional variation in cortical processing, generally
with successively more complex processing as one moves
rostrally. Greater processing demands on neurons appear to
be reflected in the increasing complexity of their dendritic
arrays (Elston 2003, 2007; Jacobs and Scheibel 2002).
Although it is not possible to know which functional cortical
areas are contained in the frontal and occipital samples in the
present study, examination of dendritic complexity, as
revealed by superficial pyramidal neurons, may indicate
regional variation of integrative functions in the elephant.
Materials and methods
Specimens
Two (LA1 and LA3; Fig. 2a), free ranging, solitary male
African elephants (Loxodonta Africana; approximate age
Fig. 2 a Photograph of elephant LA1; b superior and c inferior views
of the elephant brain illustrating the relative position of sampled
tissue blocks (shaded areas) for the occipital and frontal regions,
respectively. Note that, in c, the right hemisphere’s olfactory bulb has
been removed in order to view the orbital gyri and facilitate removal
of the tissue block. Scale bar 5 cm
c
Brain Struct Funct (2011) 215:273–298 275
123

20–30 years) scheduled for population management culling
were humanely killed as described in Manger et al. (2009).
In situ perfusion-fixation of the brains was conducted by
removal of the head, flushing of the cranium with cold
saline, and intra-carotid perfusion with 4% paraformalde-
hyde in 0.1 M phosphate buffer, resulting in relatively
short autolysis times (LA1: 160 min; LA3: 110 min). The
brains were then removed from the skull, placed in the
same cold fixative, weighed (LA1: 5,145 g, LA3: 4,835 g),
and stored in 4% paraformaldehyde in 0.1 M phosphate
buffer for 72 h. Small tissue blocks containing the cortical
regions of interest were removed and stored in 0.1%
sodium azide in 0.1 M phosphate buffer saline at 4�C for
8 months before staining. The present research protocol
was approved by the Colorado College Institutional
Review Board (#H94-004) and by the University of the
Witwatersrand Animal Ethics Committee (Manger et al.
2009).
Tissue selection
Frontal and occipital tissue blocks (1–2 cm, perpendicular
to the long axis of the sampled gyri) were removed from
the right hemisphere. Because no cytoarchitectural criteria
exist in the elephant for identification of these cortical
regions, localization was based solely on gross examination
of elephant brains (Haug 1970; Hakeem et al. 2005;
Shoshani et al. 2006) and on reports from other species
(e.g., giraffes: Badlangana et al. 2007; cetaceans: Hof and
Van der Gucht 2007). Occipital tissue was removed from
the parieto-occipital contour close to the midline, approx-
imately 3–5 cm immediately anterior to the cerebellum,
although it should be noted that the elephant occipital lobe
is not as clearly demarcated as in humans (Shoshani et al.
2006; Fig. 2b). Frontal tissue was removed from the
anterior portion of what appeared to be the orbital gyri,
3–5 cm from the midline, in a position superior to the
anterior third of the olfactory bulb (Fig. 2c).
Tissue blocks were trimmed to 3–5 mm in thickness,
coded to prevent experimenter bias, and processed by a
modified rapid Golgi technique (Scheibel and Scheibel
1978b). To be consistent with previous studies (Jacobs
et al. 1997, 2001), processed tissue was serially sectioned
at 120 lm with a vibratome. Tissue blocks adjacent to
those removed for Golgi analysis were examined with a
routine Nissl stain to establish laminar and cortical depth.
Thirty depth measurements for each layer were averaged
across 12 sections from each cortical region.
Neuron selection and quantification
Analyses qualitatively and quantitatively characterized
elephant neocortical neurons (N = 75), with a particular
focus on superficial (i.e., layers II–III) pyramidal neurons
(mean soma depth = 794 ± 131 lm) in the two cortical
regions. Because the Golgi impregnation was not uniform,
more neurons appropriate for quantification were sampled
from LA1 (n = 46) than from LA3 (n = 29). A total of 40
superficial pyramidal neurons were traced in the occipital
cortex (15 from LA1; 5 from LA3) and in the frontal cortex
(20 from LA1); of these, 16 had relatively complete apical
dendrites, which were analyzed only descriptively.
Although pooling data across animals was not an optimal
solution, an ANOVA comparing occipital total dendritic
length in superficial neurons between LA3 and LA1 found
no significant difference, suggesting this as an acceptable
compromise for subsequent analyses. A broader sampling
of neuron morphology resulted in 16 additional, deep layer
III to layer VI pyramidal neurons being traced in occipital
cortex (10 from LA1; 6 from LA3), and 19 in frontal cortex
(1 from LA1; 18 from LA3).
Previously established selection criteria for human su-
pragranular pyramidal neurons (Jacobs et al. 1997, 2001;
Anderson et al. 2009) were adapted in the present study.
Briefly, selected neurons were required to be relatively
isolated and unobscured, to appear fully impregnated with
a soma roughly centered within the 120 lm-thick section,
and to have as complete dendritic arbors as possible. In
order to create a relatively homogeneous cell population
for superficial pyramidal neurons, a running average of
soma depth from the pial surface was maintained and cells
were chosen to preserve similar averages across both cor-
tical areas.
Cells were quantified along x-, y-, and z-coordinates
using an Olympus BH-2 microscope under a Planachromat
409 (NA = 0.70) dry objective interfaced with a Neu-
rolucida software system (MBF Bioscience, Williston, VT,
USA). The system utilized a MicroFire Digital CCD 2-
Megapixel camera (Optronics, Goleta, CA, USA), which
was mounted on a trinocular head. The camera image was
viewed on a Dell E248WFP 24-inch LCD monitor with
1,920 9 1,200 resolution. First, the soma was traced at its
widest point in the two-dimensional plane in order to
provide an estimate of its cross-sectional area. Dendrites
were then traced somatofugally in their entirety, account-
ing for dendritic diameter and quantifying all visible
spines, regardless of spine type or differences in spine
length. Dendritic processes were not followed into adjacent
sections; as such, broken tips and unclear terminations
were identified as incomplete endings.
Neurons were traced by three observers (BJ, JL, and
KA). Intrarater reliability was assessed by having each
rater trace the same dendritic branch, with somata and
spines, 10 times. Tracings varied little, as indicated by
the relatively low coefficient of variation for soma size
(2.8%), total dendritic length (TDL 2.8%), and dendritic
276 Brain Struct Funct (2011) 215:273–298
123

spine number (DSN 3.9%). Intrarater reliability was also
tested with a split plot design (a = 0.05), which revealed
no significant difference within tracers between the first
five tracings and the last five tracings of a dendritic
system. For interrater reliability, tracers were normed by
comparing their tracings of 10 dendritic systems to the
same tracings completed by the primary investigator (BJ).
Pearson’s product correlations across soma size, TDL,
and DSN (defined below) averaged 0.97, 0.96, and
0.96, respectively. An analysis of variance (ANOVA;
a = 0.05) indicated that there was no significant differ-
ence among tracers in the three measures. Finally, the
primary investigator reexamined all final tracings to
ensure accuracy.
Cell descriptions and dependent dendritic/spine
measures
Deep pyramidal neurons were qualitatively described
according to nomenclature found in previous neuro-
morphological research (e.g., Ngowyang 1932; Ferrer et al.
1986a, b). Descriptions for each neuron used qualitative
criteria, such as cortical location, size, dendritic field
patterns, presence of spines, and soma characteristics.
Nevertheless, because axons were not visible and because
intermediates between different neuronal types exist, these
designations remain tentative.
For quantitative measures, a centrifugal nomenclature
was used (Bok 1959; Uylings et al. 1986): dendritic
branches arising from the soma are first-order segments
until they bifurcate into second-order segments, which
branch into third-order segments, and so on. In addition to
noting soma size (as apparent surface area in lm2) and
soma depth from the pial surface, five previously estab-
lished measures (Jacobs et al. 2001) were used to analyze
each neuron. Total dendritic length (TDL, lm) refers to
the summed length of all dendritic segments. Mean seg-
ment length (MSL, lm) is the average length of each
dendritic segment. Dendritic segment count (DSC) rep-
resents the number of dendritic segments. Dendritic spine
number (DSN) refers to the total number of spines on
dendritic segments. Dendritic spine density (DSD) is the
average number of spines per micron of dendritic length.
Additionally, dendritic diameter throughout the cell was
determined, thus providing a sixth measure: dendritic
volume (Vol, lm3). Although tested independently, many
of these measures are inherently interrelated. Finally,
dendritic branching patterns of all traced neurons were
examined using Sholl analysis (Sholl 1953), which
quantified the number of dendritic intersections at 20-lm
intervals radiating centrifugally from the soma. This
analysis assisted in the characterization of apical and
basilar dendrites.
Independent variables and statistical analyses
For superficial pyramidal neuron comparisons, only basilar
dendrites were examined inferentially because apical den-
drites were often incomplete. Basilar dendritic data were
aggregated by neuron (CELL), and brain region (AREA).
The resulting dataset was analyzed using a one-way anal-
ysis of variance (ANOVA; a = 0.05) to investigate the
effects of area (i.e., frontal and occipital) on each depen-
dent measure for the basilar skirt.
Results
Overview
Nissl-stained sections indicated that the frontal cortex was
slightly thicker than the occipital cortex, with both regions
exhibiting a relatively thick layer III and lacking a distinct
layer IV (Fig. 3; Table 1). The majority of cells traced
resided in layers III and V. In Golgi preparations, the
dominant characteristic across both occipital and frontal
Fig. 3 Photomicrographs of Nissl-stained cortex from frontal (a) and
occipital (b) cortex with labeled layers. Note that the elephant appears
to lack a well-developed layer IV. Scale bar 1 mm
Table 1 Laminar and cortical depth from the pial surface in elephant
cortex (lm)
Frontal region Occipital region
Layer I/II junction 560 ± 14 519 ± 20
Layer II/III junction 730 ± 13 675 ± 19
Layer III/V junction 1,495 ± 14 1,643 ± 38
Layer V/VI junction 2,034 ± 21 2,098 ± 42
Gray/white matter junction 2,828 ± 41 2,737 ± 55
Values represent mean ± SEM
Brain Struct Funct (2011) 215:273–298 277
123

regions was the vast heterogeneity of spiny cell types
(Fig. 4). Generally, superficial layers contained many
small, bifurcating variations of the pyramidal cell with
thick apical dendrites that ramified as they ascended toward
the pial surface. Deeper neurons possessed broad basilar
skirts and often extended multiple widely diverging apical
dendrites. In some cells, a prominent basilar dendrite or
taproot penetrated the white matter. Aspiny neurons tended
to possess long dendrites that bifurcated near the soma or
not at all, and were located in upper layer III or lower layer
V. Tracings of individual cell types are provided in Figs. 5,
6, and 7 with quantitative data provided in Table 2. It
should be noted that laminar boundaries are not indicated
in Figs. 5, 6, and 7 because, given the considerable vari-
ability across individual sections, neurons of similar depth
may not necessarily be located in the same cortical layer
across different sections. These descriptions, based on
examination of all Golgi-stained sections as well as the
quantified neurons themselves, are presented below with
superficial pyramidal neurons being considered separately
because they were more extensively analyzed.
Spiny neurons
Deeper pyramidal neurons with ascending apical dendrites
exhibited several variations: magnopyramidal (with and
without a taproot), multiapical, and fork neurons. Magno-
pyramidal neurons (n = 3, Fig. 6F–H), located at a depth
of approximately 1,500 lm in layer III of the occipital
lobe, projected an ascending, typically bifurcating, apical
process, often exceeding 1,200 lm in length. Their somata
varied in size (Table 2) and shape, from small and trian-
gular to very large and fusiform. The basilar skirt extended
in all directions to produce a circular receptive area. The
magnopyramidal cells were generally less spiny than their
superficial counterparts, with a DSD between 0.33 and 0.47
(Table 2). Sholl analysis revealed a high density of basilar
dendrites similar in size and distribution to those of
superficial pyramidal cells, although the apical dendrite
was always longer (Fig. 8a).
An apparent variation of the above was the magnopy-
ramidal-taproot neurons (n = 4, Figs. 4D, 5I–L, 9), which
were located in layers III and V of the frontal lobe. Because
of their prominent appearance in Golgi-stained sections
and their trunk-like taproot, we named these ‘‘matriarch’’
neurons (referring to the matriarch’s prominent role in
elephant society). They exhibited extensive dendritic
branching, which suggests a broad sampling of cortical
information (Brown et al. 2008). The matriarch neuron’s
fusiform soma extended an apical process for up to 900 lm
that often bifurcated during its ascent to the pial surface. A
basilar skirt projected densely and uniformly in all direc-
tions. Unique to this neuron was the addition of a complex,
descending taproot that tended to bifurcate widely into
smaller segments while the main branch extended for long
distances (up to 1 mm) toward the underlying white matter.
Fig. 4 Low magnification photomicrograph of elephant frontal
cortex illustrating a wide diversity of neuronal morphologies: inverted
pyramidal neurons (A, C); superficial pyramidal neurons (B, E);
magnopyramidal-taproot or matriarch neuron (D, also seen in
Fig. 5J); and fork neuron (F, also seen in Fig. 5F). Pial surface is
at the top of the photomicrograph. Scale bar 500 lm
278 Brain Struct Funct (2011) 215:273–298
123

DSD ranged from 0.32 to 0.54 (Table 2). In the Sholl
analysis, the matriarch neuron (Fig. 8c) was second only to
the neurogliaform cell (Fig. 7L) in terms of overall den-
dritic density.
Multiapical pyramidal neurons (n = 3, Figs. 5M, 6K–L),
present both in frontal and in occipital cortex, were among
the deepest of the spiny cells, located predominantly in
layer V at a depth of 1,500–1,700 lm (Table 2). Although
often incomplete due to sectioning, multiple, prominent
apical dendrites extended from a spherical cell body to
ascend symmetrically toward the pial surface. Basilar
dendrites extended with some variability from the soma,
either favoring radial processes that descended toward the
white matter or more lateral, tufted projections. Dendrites
possessed a relatively low density of spines, with a DSD
ranging from 0.30 to 0.36 (Table 2). Sholl analysis
revealed two apical dendritic peaks, with the second peak
being somewhat larger (Fig. 8e). Basilar dendritic density
peaked closer to the soma than did the apical dendrite.
A fork neuron (or Gabelzelle; n = 1, Figs. 4F, 5F),
originally described by Ngowyang (1932) in the human
frontoinsular cortex, was found at the boundary between
layers III and V of the frontal cortex. It possessed a split
soma that tapered into two thick, ascending apical den-
drites 600 lm in length. Basilar projections extended
obliquely as fine, parallel dendritic processes. DSD was
relatively high at 0.58 (Table 2). Its axon emerged proxi-
mally from a basilar dendrite and appeared to descend. The
fork neuron was unusual in that, like the horizontal pyram-
idal cell, its apical and basilar branches were similar in
length (Fig. 8g), although the apical dendrites were clearly
truncated by sectioning.
Greater variation in apical dendritic orientation was
observed with horizontal and inverted pyramidal neurons.
Horizontal pyramidal neurons (n = 2, Fig. 6I–J), found in
layer III of the occipital lobe, possessed a narrow, elon-
gated soma from which a quickly ramifying apical dendrite
extended laterally or obliquely (\45�). Basilar dendrites
Fig. 5 Neurolucida tracings of spiny neurons in the elephant frontal
cortex presented to indicate their relative soma depths from the pial
surface (in lm): crab-like neuron (A); superficial pyramidal neurons
(B–E); fork neuron (F); inverted pyramidal neuron (G, H);
magnopyramidal-taproot or matriarch neurons (I–L); multiapical
pyramidal neuron (M). The quantitative dependent measures for all
neurons are provided in Table 2. Scale bar 100 lm
Brain Struct Funct (2011) 215:273–298 279
123

branched into distinct, parallel collaterals or extended
numerous secondary and tertiary dendrites. DSD was
between 0.44 and 0.47 for the two cells (Table 2). In the
Sholl analysis, the apical and basilar dendrites were
roughly equal in density and length (Fig. 8f). Inverted
pyramidal neurons (n = 3, Figs. 4A, C, 5G–H, 6A), found
in layer III of frontal and occipital cortices, possessed
apical dendrites that descended and bifurcated from a
roughly triangular soma. In the frontal lobe, the length of
the apical process varied from 600 to 1,100 lm and pos-
sessed few collaterals. The basilar skirt projected toward
the pial surface, producing a circular receptive area. The
smallest and most superficial of these cells (Fig. 6A) pos-
sessed a short apical shaft that descended obliquely for less
than 400 lm in length, and an axon that emerged from the
base of the soma and bifurcated, one branch ascending and
the other descending. DSD ranged from 0.39 to 0.46
(Table 2). Sholl analyses confirmed the inverted pyramidal
neuron’s (Fig. 8b) structural similarity to the magnopyra-
midal neurons.
Finally, two ‘‘atypical’’ spiny neuron variants appeared
to have horizontally oriented dendritic arbors: flattened
pyramidal and ‘‘crab-like’’ neurons. Flattened pyramidal
neurons (n = 3, Fig. 6M–N, P) were found in deep layer
III and upper layer VI of the occipital cortex, where they
appeared to be fairly common. Flattened pyramidal neu-
rons sent multiple, thick apical dendrites from opposing
ends of the soma to ascend outward and then gradually
toward the pial surface. Somata varied from elongated to
globular. Dendrites extended further laterally than verti-
cally, with basilar dendrites often descending toward the
white matter alongside apical dendrite collaterals. Their
thick dendrites made the cells, by volume, among the
largest in the present sample. Their DSD was similar to
other large pyramidal cells, ranging from 0.37 to 0.41
(Table 2). Sholl analysis revealed the maximum density of
apical dendrites to be further from the soma than that of the
basilar dendrites (Fig. 8d). Crab-like neurons (n = 3,
Figs. 5A, 6B, O) were so designated because their dendritic
branches emerged from opposite ends of a rounded soma to
ramify symmetrically, producing claw-like shapes. These
were located in layers II and III and at the junction between
layer V and VI of the frontal and occipital lobes. DSD
varied greatly, ranging from 0.39 to 0.67 (Table 2). Sholl
analysis indicated that crab-like neurons (Fig. 8j) exhibited
a relatively small dendritic receptive area.
Fig. 6 Neurolucida tracings of spiny neurons in the elephant
occipital cortex presented to indicate their relative soma depths
from the pial surface (in lm): inverted pyramidal neuron (A); crab-like
neurons (B, O); superficial pyramidal neurons (C–E); magnopyramidal
neurons (F–H); horizontal pyramidal neurons (I–J); multiapical pyrami-
dal neurons (K–L); flattened pyramidal neurons (M–N, P). The quanti-
tative dependent measures for all neurons are provided in Table 2. Scalebar 100 lm
280 Brain Struct Funct (2011) 215:273–298
123
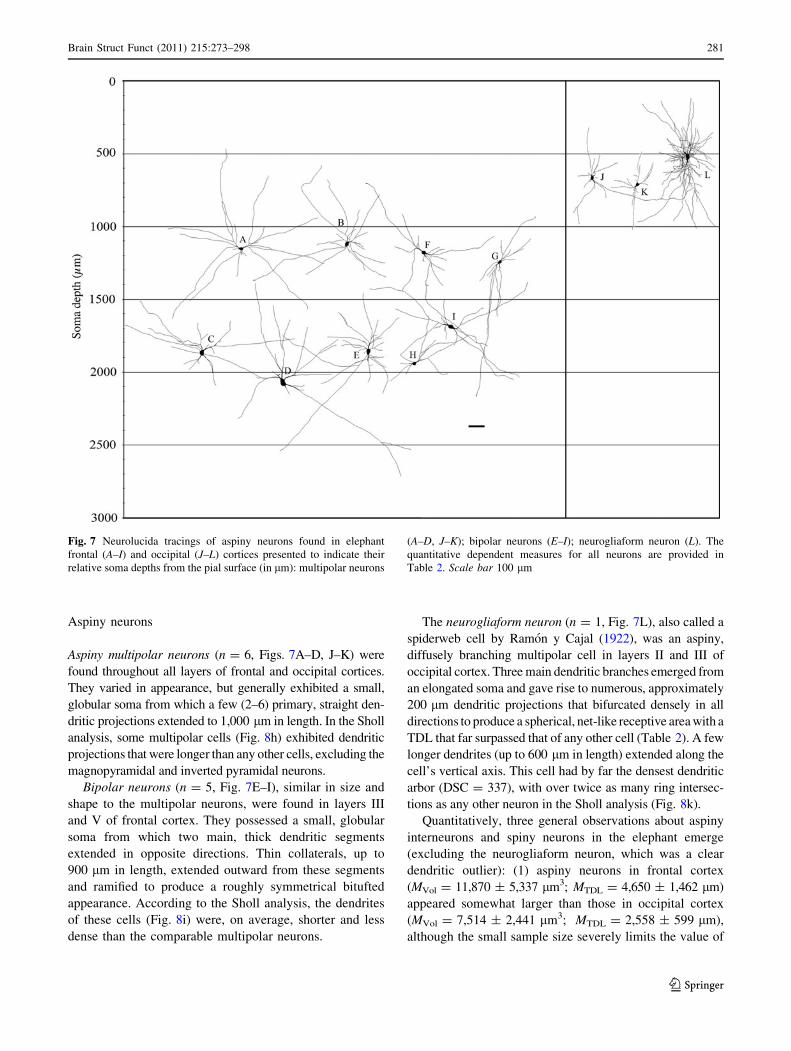
Aspiny neurons
Aspiny multipolar neurons (n = 6, Figs. 7A–D, J–K) were
found throughout all layers of frontal and occipital cortices.
They varied in appearance, but generally exhibited a small,
globular soma from which a few (2–6) primary, straight den-
dritic projections extended to 1,000 lm in length. In the Sholl
analysis, some multipolar cells (Fig. 8h) exhibited dendritic
projections that were longer than any other cells, excluding the
magnopyramidal and inverted pyramidal neurons.
Bipolar neurons (n = 5, Fig. 7E–I), similar in size and
shape to the multipolar neurons, were found in layers III
and V of frontal cortex. They possessed a small, globular
soma from which two main, thick dendritic segments
extended in opposite directions. Thin collaterals, up to
900 lm in length, extended outward from these segments
and ramified to produce a roughly symmetrical bitufted
appearance. According to the Sholl analysis, the dendrites
of these cells (Fig. 8i) were, on average, shorter and less
dense than the comparable multipolar neurons.
The neurogliaform neuron (n = 1, Fig. 7L), also called a
spiderweb cell by Ramon y Cajal (1922), was an aspiny,
diffusely branching multipolar cell in layers II and III of
occipital cortex. Three main dendritic branches emerged from
an elongated soma and gave rise to numerous, approximately
200 lm dendritic projections that bifurcated densely in all
directions to produce a spherical, net-like receptive area with a
TDL that far surpassed that of any other cell (Table 2). A few
longer dendrites (up to 600 lm in length) extended along the
cell’s vertical axis. This cell had by far the densest dendritic
arbor (DSC = 337), with over twice as many ring intersec-
tions as any other neuron in the Sholl analysis (Fig. 8k).
Quantitatively, three general observations about aspiny
interneurons and spiny neurons in the elephant emerge
(excluding the neurogliaform neuron, which was a clear
dendritic outlier): (1) aspiny neurons in frontal cortex
(MVol = 11,870 ± 5,337 lm3; MTDL = 4,650 ± 1,462 lm)
appeared somewhat larger than those in occipital cortex
(MVol = 7,514 ± 2,441 lm3; MTDL = 2,558 ± 599 lm),
although the small sample size severely limits the value of
Fig. 7 Neurolucida tracings of aspiny neurons found in elephant
frontal (A–I) and occipital (J–L) cortices presented to indicate their
relative soma depths from the pial surface (in lm): multipolar neurons
(A–D, J–K); bipolar neurons (E–I); neurogliaform neuron (L). The
quantitative dependent measures for all neurons are provided in
Table 2. Scale bar 100 lm
Brain Struct Funct (2011) 215:273–298 281
123

Table 2 Summary statistics for elephant neurons in frontal and occipital cortex
Type Cella Vol.b TDLc MSLc DSCd DSNe DSDf Soma sizeg Soma depthh
Frontal
Crab-like 5A 5,732 4,249 83 51 2,863 0.67 439 424
Superficial pyramidal 5B 8,233 4,574 82 56 2,036 0.45 286 741
Superficial pyramidal 5C 17,880 6,740 95 71 4,162 0.62 478 691
Superficial pyramidal 5D 30,300 7,603 104 98 4,839 0.64 479 818
Superficial pyramidal 5E 17,250 8,233 106 78 5,379 0.65 506 875
Fork 5F 55,256 8,050 117 69 4,685 0.58 1,154 1,436
Inverted pyramidal 5G 13,937 5,634 95 59 2,608 0.46 681 1,101
Inverted pyramidal 5H 18,055 7,370 93 79 3,012 0.41 706 1,203
Matriarch 5I 27,149 7,716 100 77 2,489 0.32 1,061 1,066
Matriarch 5J 33,500 10,838 111 97 5,856 0.54 766 1,299
Matriarch 5K 43,904 8,269 115 72 3,977 0.48 973 1,317
Matriarch 5L 28,360 10,377 113 92 5,430 0.52 799 1,423
Multiapical pyramidal 5M 54,261 6,742 98 69 2,340 0.35 1,552 1,662
Multipolar 7A 10,843 6,879 186 37 – – 387 1,124
Multipolar 7B 9,311 5,152 152 34 – – 611 1,069
Multipolar 7C 11,905 3,778 114 33 – – 801 1,811
Multipolar 7D 24,569 5,839 195 30 – – 1,375 2,027
Bipolar 7E 13,497 5,883 115 51 – – 757 1,811
Bipolar 7F 9,186 4,129 118 35 193 0.05 494 1,132
Bipolar 7G 8,510 3,996 125 32 – – 351 1,203
Bipolar 7H 5,851 1,946 97 20 – – 411 1,893
Bipolar 7I 13,160 4,248 112 38 – – 737 1,641
Occipital
Inverted pyramidal 6A 5,173 3,465 96 36 1,346 0.39 398 823
Crab-like 6B 4,596 2,500 104 24 1,214 0.49 407 806
Superficial pyramidal 6C 10,897 5,921 82 72 3,346 0.57 505 722
Superficial pyramidal 6D 16,264 5,509 115 48 3,630 0.66 643 746
Superficial pyramidal 6E 11,312 6,752 83 81 3,914 0.58 256 650
Magnopyramidal 6F 18,924 8,752 115 76 4,094 0.47 780 1,470
Magnopyramidal 6G 76,591 7,925 113 70 3,148 0.40 2,015 1,487
Magnopyramidal 6H 46,090 6,895 90 77 2,296 0.33 2,004 1,525
Horizontal pyramidal 6I 29,131 7,554 113 67 3,521 0.47 988 1,571
Horizontal pyramidal 6J 28,811 6,411 88 73 2,853 0.44 941 1,090
Multiapical pyramidal 6K 35,055 4,428 84 53 1,311 0.30 1,562 1,536
Multiapical pyramidal 6L 32,114 6,375 133 48 2,291 0.36 1,284 1,734
Flattened pyramidal 6M 58,109 6,187 115 54 2,293 0.37 1,694 1,456
Flattened pyramidal 6N 36,137 4,346 92 47 1,706 0.39 1,222 1,407
Crab-like 6O 13,679 4,938 60 83 1,949 0.39 689 2,003
Flattened pyramidal 6P 57,494 6,476 93 70 2,674 0.41 1,092 2,233
Multipolar 7J 9,241 2,981 110 27 210 0.07 479 672
Multipolar 7K 5,788 2,134 71 30 34 0.02 423 704
Neurogliaform 7L 26,959 14,818 44 337 – – 857 528
a Refers to tracings of individual cells as indentified in Figs. 5, 6, 7b Volume in lm3
c Length in lmd Number of segments per neurone Number of spines per neuronf Number of spines per lm of dendritic lengthg Soma size in lm2
h Soma depth in lm from the pial surface
282 Brain Struct Funct (2011) 215:273–298
123

this comparison; (2) the aspiny interneurons (MVol =
11,078 ± 5,147 lm3; MTDL = 4,270 ± 1,569 lm) exhib-
ited substantially less overall dendritic extent than the
spiny neurons (MVol = 28,765 ± 11,078 lm3; MTDL =
6,580 ± 1,906 lm); and (3) the aspiny interneurons
exhibited considerably longer (MMSL = 127 ± 37 lm),
but fewer (MDSC = 33 ± 8) dendritic branches than did the
spiny neurons (MMSL = 100 ± 15 lm; MDSC = 67 ± 17).
Superficial pyramidal neurons
Superficial pyramidal neurons (n = 40; Figs. 4B, E, 5B–E,
6C–E) were well impregnated across both occipital and
frontal cortices (Fig. 10), facilitating a more detailed
quantitative analysis. These neurons were sampled pri-
marily from upper cortical layer III with minimal variation
of soma depth across cortical areas (frontal 818 ± 108 lm;
occipital 769 ± 149 lm). Measurements of soma size
indicated minimal variation between cortical areas (frontal
515 ± 80 lm2; occipital 517 ± 156 lm2). There was a
significant positive correlation between soma depth and
soma size (r(40) = 0.451, p \ 0.01), as well as between
soma size and volume (r(40) = 0.353, p \ 0.05), which
underscored the importance of maintaining similar soma
depths between areas for subsequent comparisons.
In terms of appearance, superficial pyramidal neurons
exhibited typical basilar skirts, which projected radially
for a short distance from the soma. They were charac-
terized by apical dendrites that generally bifurcated
around 50 lm from the soma to form two, ascending
apical shafts (Figs. 5D–E, 6C, E, 10a, d), or by two
diverging apical dendrites (Figs. 5C, 6D) that emerged
directly from the soma. These resulting apical trunks
traveled obliquely to the pial surface, thus creating a large
apical arbor for each cell and a crisscross or ‘‘V’’
appearance in the upper layers (Fig. 11b–c). Generally,
these branches appeared to bundle with other branches
traveling at similar angles toward the pial surface
(Fig. 11a, d).
Dendritic morphology-dependent measures
Selected dependent measures for basilar data were first
analyzed using a MANOVA test statistic. Due to direct
Fig. 8 Sholl analyses of 11 cell types assessing the relative
complexity of basilar, apical, and total dendritic branching patterns.
Concentric rings, separated by 20 lm increments and centered on the
soma, were used to measure dendritic intersections. Neurons a–g were
spiny. Neurons h–k were aspiny. a–c exhibited relatively long apical
dendrites, d–g had shorter apical dendrites. For cells with basilar
dendrites, the density of basilar intersections peaked before 200 lm,
as was the case for the total dendritic density of all aspiny neurons.
The apical dendrite either tended to extend for a much larger distance
than the basilar dendrites (a, b) or was nearly equal to them (c–g).
Aspiny neurons (h–j) generally had low dendritic densities, the
notable exception being the neurogliaform neuron (k), which had the
highest dendritic density of all neurons. Asterisks represents that the
y-axis for the neurogliaform graph (k) is set to a larger scale than
other cells to compensate for its particularly high dendritic density
Brain Struct Funct (2011) 215:273–298 283
123

associations between DSD and DSC with other measures,
these variables were not used in the MANOVA. The
MANOVA was significant (F(4, 35) = 3.125, p \ 0.05,
gp2 = 0.263). Post hoc analyses, consisting of six one-way
ANOVAs, were then conducted for each dependent vari-
able. All dependent measures, except MSL and DSD, were
significantly higher in frontal cortex, indicating the pres-
ence of more complex basilar dendrites. Although there
were not enough complete apical branches to warrant sta-
tistical tests, descriptive data indicated a similar pattern for
apical dendrites. The differences in complexity can be seen
visually in Fig. 12, which contains representative tracings
from each region.
Volume
Basilar dendritic volume was significantly greater (F(1, 38)
= 5.80, p \ 0.05, gp2 = 0.132) in frontal than in occipital
cortex (by 25.2%; Fig. 13a). On average, neurons from
frontal cortex contained 29.5% thicker apical dendrites and
23.3% more overall volume than neurons from occipital
cortex.
Dendritic length
Basilar dendrites in frontal cortex exhibited significantly
greater TDL (F(1, 38) = 7.15, p \ 0.05, gp2 = 0.158) than
basilar dendrites in occipital cortex (Fig. 13b). The trend
was in the opposite direction for MSL, but was not sig-
nificant (Fig. 13c). Both TDL (by 19.0%) and MSL (by
5.0%) were greater for apical dendrites in the frontal cortex
than in the occipital cortex. Descriptive data from complete
neurons (i.e., basilar and apical dendrites combined)
showed that, overall, TDL was greater (by 13.5%) and
MSL was shorter (by 9.2%) in the frontal cortex than in the
occipital cortex.
Dendritic segment count
There was a significant difference in basilar DSC across
both areas (F(1, 38) = 10.93, p \ 0.05, gp2 = 0.223), with
basilar dendrites in frontal cortex exhibiting 25.1% higher
DSC than occipital cortex (Fig. 13d). Apical dendrites also
appeared to have higher DSC in frontal cortex (by 14.8%),
and complete neurons in frontal cortex had 20.7% higher
DSC than those in occipital cortex.
Dendritic spine number and density
Basilar dendrites of neurons in the frontal cortex displayed
a significantly higher DSN (by 25.1%; Fig. 13e) than
neurons in the occipital cortex (F(1, 38) = 7.95, p \ 0.01,
gp2 = 0.173). DSD was not significant (Fig. 13f). Apical
dendrites in frontal cortex contained 29.4% more spines
and had a 13.4% higher DSD than those in occipital cortex.
Evidence from complete neurons indicated that frontal
Fig. 9 Photomicrographs of two magnopyramidal-taproot or matri-
arch neurons (a, b) with higher magnification insets of their spine-rich
ascending apical (a) and descending taproot (t) dendrites. Neurolucida
tracings can be seen in Fig. 5L for a, and in Fig. 5I for b. Scale bar100 lm for a and b, and 25 lm for the insets
284 Brain Struct Funct (2011) 215:273–298
123

Fig. 10 Photomicrographs of Golgi-impregnated superficial pyrami-
dal neurons in occipital (a) and frontal (d) cortices. Associated
photomicrographs of apical (occipital: b; frontal f) and basilar
(occipital: c; frontal e) dendrites are also shown at higher
magnifications. Note in both cortical regions the presence of
bifurcating apical dendrites. For a, b, d, f scale bars 50 lm. For c,
e scale bars 25 lm
Brain Struct Funct (2011) 215:273–298 285
123

superficial neurons contained 20.2% more spines and had a
7.6% higher DSD than did occipital neurons.
Sholl analysis
Frontal and occipital regions had similar shapes in Sholl
analysis profiles, but frontal results were slightly
higher on average (Fig. 14a, b). Basilar dendrites in both
areas reached their maximum complexity (frontal
16.45 ± 0.99 intersections; occipital 12.80 ± 0.86 inter-
sections) 120 lm from the soma, and apical dendritic
complexity peaked around 200 lm from the soma
(frontal 4.88 ± 0.50 intersections; occipital 4.10 ± 0.48
intersections).
Comparison to human supragranular pyramidal neurons
Elephant superficial pyramidal neurons were compared to
supragranular pyramidal neuron data gathered with similar
methodology in human inferior frontal (specifically, area
11, from Jacobs et al. 2001) and occipital regions (area 18,
from Jacobs et al. 1997). These regions were chosen to be a
relatively close topographic match for those examined in
the elephant. The human supragranular pyramidal neurons
Fig. 11 Photomicrographs of V-apical bundling within elephant
occipital cortex. At lower magnifications (b and c), a V-shaped
crossing pattern resulting from bifurcating apical dendrites is
apparent. At higher magnifications (a and d), apical dendrites appear
to bundle together as they obliquely approach the pial surface. For
a and b scale bars 50 lm. For c scale bar 100 lm. For d scale bar25 lm
286 Brain Struct Funct (2011) 215:273–298
123

(n = 116) were traced in 6 neurologically normal subjects
(4 men, 23–69 years of age; 2 women, 32 and 34 years of
age).
Dendritic length and segment count
On average, elephant superficial pyramidal neurons dem-
onstrated only slightly higher TDL than human neurons
both in frontal (by 6.8%) and in occipital (by 2.9%) cor-
tices (Fig. 15a). However, TDL was manifested differently
in humans and elephants, as revealed by MSL and DSC
values. Basilar dendrites in the elephant exhibited greater
MSL both in frontal (by 23.3%) and in occipital cortices
(by 35.6%; Fig. 15b) than they did in humans. In contrast,
elephant DSC was considerably lower than human DSC
(by 16.0% in frontal cortex, and by 33.5% in occipital
cortex; Fig. 15c).
Dendritic spines
Elephant superficial pyramidal neurons exhibited greater
values for DSN and DSD than did human neurons. In
elephant frontal cortex, basilar dendrites exhibited 63.6%
higher DSN and 61.7% higher DSD than in humans. In
elephant occipital cortex, basilar dendrites exhibited 52.3%
higher DSN and 49.9% higher DSD than in humans.
Sholl analysis
Although Sholl analysis profiles were similar across both
area 11 and area 18 in human, they varied considerably
from those in the elephants (Fig. 14). On average, elephant
basilar dendrites extended 760 lm from the soma in frontal
cortex and 740 lm from the soma in occipital cortex. In
contrast, human basilar dendrites extended 380 lm from
the soma in the frontal cortex and 400 lm in occipital
cortex. At the high point of dendritic intersections, human
supragranular pyramidal neurons contained more intersec-
tions than did elephant superficial pyramidal neurons. In
frontal cortex, human basilar dendrites peaked at 22.90 ±
0.90 intersections, whereas elephant dendrites peaked at
16.45 ± 0.99 intersections. In occipital cortex, human
basilar dendrites peaked at 21.47 ± 0.65 intersections,
whereas elephant dendrites peaked at 12.80 ± 0.86 inter-
sections. Overall, the Sholl analysis profiles confirm that
human supragranular pyramidal neurons were more con-
densed and contained more branch points than the elephant
superficial pyramidal neurons.
Fig. 12 Sample tracings of superficial pyramidal neurons, and their
corresponding dendritic measurements, from frontal (cells a–d) and
occipital (cells e–h) cortices. Neurons are presented to indicate their
relative soma depths from the pial surface (in lm). Within each
cortical area, the two neurons with relatively complete apical
dendrites appear on the left. Note also the bifurcation of all apical
dendrites. In general, frontal neurons appeared more dendritically
complex than occipital neurons. Scale bars 100 lm
Brain Struct Funct (2011) 215:273–298 287
123

Discussion
The present study is the first to document neuronal
morphology in the African elephant cortex. In terms of
cytoarchitecture, the elephant neocortex is characterized
by the presence of only five cortical layers as it altogether
lacks a visible layer IV, which is quite different from
what is observed in primates. Although it has not been
studied in much detail to date, especially in terms of
regional differences, we can make several generalizations.
Layer I appears to be relatively thick and more cellular
than in most terrestrial species. Layer II is clearly visible,
densely packed, and displays cellular clustering in many
cortical regions, particularly in the insula (Hakeem et al.
2009), as has been reported in cetaceans (Manger et al.
1998; Hof and Van der Gucht 2007). Layer II also con-
tains occasional large pyramid-like neurons. Layer III is
relatively thick and populated by large pyramidal neurons,
the density and size of which clearly vary among different
cortical domains; this fact deserves a detailed investiga-
tion. Layer V presents as a relatively thin row of very
large pyramidal cells usually distributed in small groups,
presumably representing principal efferent neurons, and a
thicker deep portion containing smaller pyramidal cells.
Layer VI is of variable thickness among cortical regions
and contains a variety of pyramidal and non-pyramidal
neurons as seen in many other mammalian species. This
cortical lamination pattern with lack of an internal gran-
ular layer IV may reflect a particular organization of
cortical connectivity in elephants, perhaps comparable to
that in cetaceans, where agranularity is also a neocortical
characteristic (Hof et al. 2005; Hof and Van der Gucht
2007). Although, we are currently exploring the cytoar-
chitecture of elephant cortex in detail, it remains clear
that complex neocortical gyrification and distinct laminar
organization represent remarkable features of the elephant
brain.
In terms of neuromorphology, elephant cortex featured a
great diversity of large, complex neurons, with consider-
able variety exhibited by ‘‘atypical’’ spiny neurons. A
prominent characteristic in elephant cortex was the
V-shaped arrangement of bifurcating apical dendrites.
Quantitatively, the dendrites of superficial pyramidal neu-
rons in the elephant frontal cortex were more complex than
those in occipital cortex. In comparison to humans, ele-
phant superficial pyramidal neurons exhibited similar
overall basilar dendritic length, but individual dendrites
tended to be longer in the elephant with less intricate
branching. Before considering the implications of these
findings, methodological issues must be addressed.
Fig. 13 Bar graphs of relative volume (a), total dendritic length (b),
mean segment length (c), dendritic segment count (d), dendritic spine
number (e), and dendritic spine density (f) of superficial pyramidal
neuron basilar dendrites in elephant frontal and occipital cortex. The
basilar dendrites in frontal cortex had significantly greater volume,
TDL, DSC, DSC, and DSN than did those in occipital cortex. Errorbars represent SEM
288 Brain Struct Funct (2011) 215:273–298
123

Methodological considerations
In general, the same constraints that apply to Golgi-stained
human tissue also apply here (Jacobs and Scheibel 2002):
(1) a small sample size in terms of subjects and sampled
neurons (Jacobs and Scheibel 1993); (2) lack of historical
information on the subjects (Jacobs et al. 1993); (3)
underestimation of spines in light microscopy (Horner and
Arbuthnott 1991); and (4) inherent issues with Golgi
impregnations (Braak and Braak 1985). These general
limitations are accepted and we focus here on three broader
issues.
Functional classification of elephant cortex and regional
specialization
Functional characteristics of the sampled regions could not
be determined. The frontal cortex, although located in what
appear to be anterior orbital gyri, could subserve prefrontal
executive functions (Barbas 1995) or, equally likely,
premotor activities. The function of the occipital cortex is
even more problematic, as this region could support visual
or even auditory processes. As such, it is impossible to
correlate dendritic/spine measure with the functional
attributes of these regions, as has been done in primates
(Elston and Rosa 1998a, b; Jacobs et al. 2001). Moreover,
it is not possible to make definitive statements about the
regional distribution of traced neurons in the present study
despite observing that some neuron types appeared only in
one cortical region.
Neuromorphological nomenclature
Unambiguously classifying neurons is problematic when
the criteria are not clearly defined (Bota and Swanson
2007), are too restrictive (Germroth et al. 1989), or when
cellular morphologies form a continuum rather than distinct
categories, as may be the case for extraverted telencephalic
neurons (Sanides and Sanides 1972; Ferrer and Perera
1988). Further classification issues emerge when definitions
Fig. 14 Graphic representation of Sholl analysis results for elephant
superficial and human supragranular pyramidal neurons, showing
mean numbers of dendritic intersections in elephant frontal (a),
elephant occipital (b), human frontal (c), and human occipital
(d) cortices. Although dendritic extent between areas and species is
similar, elephant dendrites are distributed across a greater distance
from the soma and exhibit fewer intersections at the peak than
observed in humans. Error bars represent SEM
Brain Struct Funct (2011) 215:273–298 289
123

change over time, as typified by the term ‘‘pyramidal,’’
which now applies to a much broader category of neurons
than it did originally (Masland 2004). Compounding the
classification problem is an essentially Euarchontoglires-
centric nomenclature in the literature, whereby the
(anthropoid) primate together with the (murid) rodent brain
constitutes the template against which all other brains are
compared, thus dictating what is ‘‘typical’’ versus ‘‘atypi-
cal’’ (Manger et al. 2008). In the present study, we have
attempted to follow existing nomenclature where possible
or, when appropriate, named cells based loosely on their
dendritic morphology (e.g., crab-like neuron). Although we
observed many examples of the presented neurons in our
examination of the Golgi-stained sections, we only traced
those that were relatively complete and unobscured, which
limits our overall conclusions regarding neuronal types.
Finally, the present study was also limited because it
described neurons based only on somatodendritic measures.
Comparison of human and elephant dendritic/spine
measures
With regard to dendritic measures, only portions captured
in the 120-lm thick section could be compared. Because
the basilar dendrites in elephants exhibited greater TDL
than in humans, the observed TDL difference between the
two species was probably attenuated (Jacobs et al. 1997).
Nevertheless, sampling within an equal volumetric ‘‘slice’’
provides useful relative measures for comparison. With
regard to spine measures, several issues prohibit a
Fig. 15 Bar graphs displaying the differences in relative total
dendritic length (a), mean segment length (b), and dendritic segment
count (c) for basilar dendrites in elephant superficial pyramidal
neurons and human supragranular pyramidal neurons across frontal
and occipital cortex. Sample tracings of elephant superficial and
human supragranular pyramidal neurons are also provided. Although
elephant and human pyramidal neurons show similar TDL values,
human neurons contain shorter segments and more intricate branch-
ing. All human data were obtained from Jacobs et al. (1997) for
occipital cortex (specifically area 18), and Jacobs et al. (2001) for
frontal cortex (specifically area 11). Error bars represent SEM
290 Brain Struct Funct (2011) 215:273–298
123

meaningful interpretation of the relative DSN and DSD
values. First, the human brains were immersion-fixed,
whereas the elephant brains were perfusion-fixed, the latter
revealing greater detail (i.e., spines; Morest and Morest
2005). Second, autolysis time in the humans was longer
(*14 h) than in the elephants (*2 h), which potentially
decreased the number of impregnated spines in the human
samples (de Ruiter 1983). Finally, although data were
collected in a methodologically similar manner, human
neurons were traced using a Neurolucida Lucivid system,
whereas elephant neurons were traced on a Neurolucida
camera system. It was recently discovered that the higher
magnification in the camera system increases the number
of spines identified by about 17% (Anderson et al. 2010),
clearly making accurate spine comparisons between the
two species problematic.
Spiny neurons
The elephant cortex is characterized by an even greater
variety of spiny neurons than the morphologically hetero-
geneous inferior temporal cortex of macaque monkeys and
rats (Germroth et al. 1989; De Lima et al. 1990). Although
several of these neuron types have been observed in other
eutherian species, what remains noteworthy in the elephant
is their arrangement, which represents a striking departure
from the vertical apical dendrites and cortical columns that
have been deemed the fundamental, if not canonical
building block of the cerebral cortex (Mountcastle 1997;
Innocenti and Vercelli 2010).
These spiny cortical neurons exist along a continuum
from those that appear more pyramid-like to those that
radically differ from the ‘‘typical’’ pyramidal neuron in
terms of morphology and/or orientation. In the elephant,
three neuron types, in addition to superficial pyramidal
neurons, approximate general pyramidal neuron morphol-
ogy: magnopyramidal, multiapical, and fork neurons. The
only exact comparison point in the literature is the single
Indian elephant neuron depicted in Barasa and Shochato-
vitz (1961), which is consistent with the present layer V
magnopyramidal neurons. Although pyramidal neurons in
the cat, dog, pig, and sheep resemble those in the elephant,
the most striking similarities are found in the cow, horse,
two-toed sloth, and anteaters (Barasa 1960; Ferrer et al.
1986b; Sherwood et al. 2009), which exhibit prominent,
bifurcating apical dendrites much the same as those
observed in elephant magnopyramidal and multiapical
neurons. Unfortunately, size comparisons with cow and
horse neurons are not possible because Barasa (1960) did
not provide scale bars. The fork neuron has been observed
in the insula and hippocampus of humans, chimpanzees,
and orangutans (Ngowyang 1936; de Crinis 1934).
Although originally considered a unique type of neuron,
subsequent investigations in several mammals (e.g., pri-
mates, carnivores, ungulates, and rodents) have concluded
that it is a type of enveloping neuron (or Umfassungszelle)
and thus is merely a variant of pyramidal neurons (Juba
1934; Syring 1956). Finally, elephant magnopyramidal,
multiapical, and fork neurons appear at least as dendritic-
ally complex as infragranular pyramidal neurons in the cat
parietal cortex (Yamamoto et al. 1987), human cingulate,
and motor cortices (Meyer 1987; Schlaug et al. 1993).
Within a more polymorphic subgroup of pyramidal
neurons are the horizontal and inverted pyramidal neurons
(Ramon y Cajal 1891; Van der Loos 1965). In terms of
morphology, these are similar to magnopyramidal and
multiapical neurons, but are slightly smaller in dendritic
extent and differ in their specific orientations. Although
horizontal pyramidal neurons have typically been noted in
superficial and deep cortical layers in primates and rodents
(Meyer 1987; Miller 1988), elephant horizontal pyramidal
neurons were located in layer III. These strongly resembled
one variant of the asymmetrical pyramidal neurons in
inferior temporal cortex (Fig. 8e of De Lima et al. 1990),
although one could argue that what De Lima et al. refer to
as a prominent, laterally oriented, basilar dendrite is actu-
ally the apical dendrite. The elephant variant appears to be
a more dendritically complex version than what has been
noted in layer VI of both lissencephalic and gyrencephalic
brains (Ferrer et al. 1986a, b, 1987). Although the function
of these neurons remains unclear, it is possible that they
contribute to the lateral integration of synaptic input (Van
Brederode et al. 2000), which has been suggested as a
major step in neocortical evolution (Ferrer et al. 1986b).
Much the same as horizontal pyramidal neurons, elephant
inverted pyramidal neurons were located in layer III. In
human temporal cortex, they have been documented in
layer V (Ong and Garey 1990). In chimpanzees, inverted
pyramidal neurons are located in layers III, V and espe-
cially VI of sensorimotor cortices, constituting a small
percentage (\1%) of all pyramidal neurons (Qi et al. 1999).
In rats, rabbits, cats, sheep, sloths, anteaters, rock hyrax,
and elephant shrews, they also tend to be located in in-
fragranular layers and constitute a small (1–8.5%) per-
centage of all neurons in the cortex (Parnavelas et al. 1977;
Mendizabal-Zubiaga et al. 2007; Sherwood et al. 2009).
These excitatory neurons have been shown to project to
ipsi- and contra-lateral cortical regions, as well as to the
claustrum and striatum (Bueno-Lopez et al. 1991; Mendi-
zabal-Zubiaga et al. 2007). In the elephant, however, the
extent to which inverted pyramidal neurons share connec-
tional and/or functional attributes with their counterparts in
other species remains unclear.
One type of elephant neuron that does not have a clear
counterpart in the literature is the magnopyramidal-taproot
or ‘‘matriarch’’ neuron of the frontal cortex. Although they
Brain Struct Funct (2011) 215:273–298 291
123

resemble, because of the descending taproot, some of the
layer VI vertical fusiform projection and asymmetric
pyramidal neurons described in macaque inferior temporal
gyrus (De Lima et al. 1990) and some of the layer VI
pyramidal neurons in human motor cortex (Meyer 1987),
they exhibited a much more extensive dendritic system.
Indeed, these were among the most complex neurons in the
present sample, approaching the dendritic extent of human
Betz or Meynert neurons. Although morphologically very
idiosyncratic, some Betz cells, like matriarch neurons,
possess a bifurcating apical shaft with branches that spread
laterally (Braak and Braak 1976), and may also exhibit a
long, descending taproot dendrite (Scheibel and Scheibel
1978a). However, unlike Betz cells, matriarch neurons
were consistently very spiny along both thin and thick
dendrites, including the taproot. Although the function of
matriarch neurons remains unknown, they may represent
extreme adaptations of pyramidal neurons, much the same
as other large cortical neurons in primates (e.g., Betz,
Meynert, and von Economo’s spindle cells; Nimchinsky
et al. 1995; Sherwood et al. 2003; Wittenberg and Wang
2007). Their morphology and potential regional localiza-
tion in the frontal cortex suggests that, like Betz cells,
matriarch neurons may sample a wide range of cortical
input and exert modulatory field effects in the surrounding
neuropil, potentially contributing to associative cognitive
processes (Kaiserman-Abramof and Peters 1972; Scheibel
and Scheibel 1978a).
Among the more ‘‘atypical’’ spiny neurons in elephant
cortex were the horizontally oriented crab-like neurons and
the flattened pyramidal neurons. One could also describe
the three crab-like neurons as horizontally oriented bitufted
neurons; similarly, one cannot rule out that the deeper of
these, in layer VI of occipital cortex (Fig. 6O), may even
have been an unusual, apical-less variant of the solitary cell
of Meynert or of the Meynert–Cajal neuron (von Economo
and Koskinas 1925). Both the crab-like and flattened
pyramidal neurons appear to be much larger variants of
neurons described in the dog and sheep as pyramidal
neurons with multiple horizontal collaterals or horizontal
pyramidal neurons (Figs. 6 and 9 of Ferrer et al. 1986b).
The flattened pyramidal neuron is also strongly reminiscent
of the large, widely bifurcating neuron observed in the cow
(Fig. 7 in Barasa 1960), but with a more pronounced lateral
extension of the apical dendrites (up to 1 mm, Fig. 6M).
The horizontal nature of these two neuronal types may
argue against a strict vertical, columnar arrangement of
cortex (Lubke et al. 2000, 2003), and again suggests a role
for lateral integration of information, perhaps related to the
low density of elephant cortical neurons (Hart et al. 2008).
What remains unclear, however, is why flattened pyramidal
neurons appeared to be more prominent in the occipital
cortex.
Aspiny neurons
Classification of aspiny interneurons typically incorporates
axonal morphology (Lund and Lewis 1993; Petilla Inter-
neuron Nomenclature Group 2008; Druga 2009), which
can be further supplemented with electrophysiological and
molecular properties (DeFelipe 1997; Zeitsev et al. 2009).
Aspiny interneurons in the present study, however, could
only be described according to their general somatoden-
dritic characteristics because axonal arbors were not
revealed by the Golgi stain. Despite this limitation, it
appeared that frontal aspiny neurons exhibited greater
dendritic spread than did those in occipital cortex. Apart
from this size difference, all elephant aspiny interneurons
could be categorized as either multipolar, bipolar, or neu-
rogliaform neurons. With the exception of one aspiny
interneuron (Fig. 7G), which had vertically oriented den-
dritic branches, all other aspiny interneurons contained a
sparse number of laterally or radially extending dendrites.
Dendritic spread in elephant aspiny interneurons, at least
in frontal cortex (with a radius of up to 1,000 lm from the
soma) was extensive. Comparatively, dendritic radii for
local circuit neurons have been reported to be approxi-
mately 50–400 lm in the rat (Kawaguchi 1995) and the cat
(Peters and Regidor 1981; Somogyi et al. 1983),
100–300 lm in the striped dolphin (Stenella coeruleoalba,
Ferrer and Perera 1988), 100–400 lm in the echidna
(Hassiotis and Ashwell 2003), and up to 500 lm in
macaque monkey (Lund and Lewis 1993) and humans
(Kisvarday et al. 1990), although aspiny interneurons in
human motor cortex have been reported with radii of up to
800 lm (Meyer 1987). Similarly, the elephant’s neuro-
gliaform neuron was morphologically similar to what has
been reported in other species, but was approximately 2–3
times larger in terms of its dendritic field (Jones 1984;
Povysheva et al. 2007). Thus, although aspiny interneuron
morphology appears to be highly conserved among
eutherian mammals (Ferrer 1986; Sherwood et al. 2009)
and in the monotreme echidna (Hassiotis et al. 2005), the
elephant variants, at least in the frontal lobe, appear to be
relatively large in terms of dendritic extent. This large size
may be required for these interneurons to exert the wide-
spread inhibitory influences required to shape the temporal
flow of information during cortical processing (Constan-
tinidis et al. 2002).
Elephant superficial pyramidal neurons and human
supragranular pyramidal neurons
Among the most spiny of cortical neurons in the elephant,
superficial pyramidal neurons were ‘‘atypical’’ insofar as
they possessed an apical dendrite with a primary bifurca-
tion at or near the soma, resulting in two obliquely
292 Brain Struct Funct (2011) 215:273–298
123

ascending secondary branches. In the literature, there
appears to be little agreement on whether these bifurcating
apical branches are characteristic of bitufted (Bartesaghi
et al. 2003), multiapical (Ferrer et al. 1986a, b), or even
extraverted pyramidal neurons (Fitzpatrick and Henson
1994). Although resembling layer II extraverted pyramidal
neurons in several other species (hedgehog, bat, and
opossum: Sanides and Sanides 1972; dolphin: Glezer and
Morgane 1990; quokka: Tyler et al. 1998; humpback
whale: Hof and Van der Gucht 2007; giant elephant shrew,
giant and lesser anteater: Sherwood et al. 2009), elephant
superficial pyramidal neurons did not meet the strict defi-
nition of extraversion, whereby apical dendritic extent
exceeds that of basilar dendrites (Sanides and Sanides
1972). In general, the elephant superficial pyramidal neu-
rons possessed a very well-developed basilar skirt, the TDL
of which exceeded that of the apical dendrite in 35 of the
40 traced neurons. Because the basilar array is considered
to be the most progressive feature of the pyramidal neuron
(Sanides and Sanides 1972), it is possible that elephant
superficial pyramidal neurons represent an intermediary
between extraverted neurons and ‘‘typical’’ primate pyramidal
neurons. This could be an adaptation that accompanies a large
brain as an extensive basilar skirt could compensate for the
relatively low neuron density in elephant cerebral cortex
(Tower 1954) by increasing the neuron’s receptive surface
area.
Similarly, the bifurcating apical dendrite greatly
increases the lateral spread of apical branches and could
represent an adaptation to the cytoarchitecture of the ele-
phant neocortex which, as in the hedgehog (Valverde and
Facal-Valverde 1986), bat (Ferrer 1987), and cetaceans
(Garey et al. 1985), does not contain a developed layer IV.
Such laminar organization is associated with an increase in
thalamic inputs into the molecular layer over the deeper
layers (Ferrer and Perera 1988; Glezer and Morgane 1990).
Bifurcating apical dendrites formed V-shaped apical bun-
dles, which is consistent with the description of small-
medium pyramidal cells in cows and horses (Barasa 1960),
but differs substantially from the canonical vertical apical
bundle-type architecture that typifies primate and rodent
neocortex (Fleischhauer et al. 1972). The resulting apical
dendritic bundling, variations of which have been observed
in other species (cat: Fleischhauer 1974; rat: Wyss et al.
1990; tenrec, red-eared pond turtle: Schmolke and Kunzle
1997; echidna: Hassiotis et al. 2003, 2005), raises inter-
esting questions regarding neocortical computation in
cognitively sophisticated species such as the elephant.
Although dendritic bundles appear to help synchronize
signals involved in cortical processing (Scheibel and
Scheibel 1975; Roney et al. 1979; Geisler et al. 2000), their
exact functions remain elusive. It is possible that this V-
shaped arrangement represents a variation of the vertical
column or serves to facilitate communication across wider
domains of cortex, either independent of columns or
between columns that are more widely spaced than in other
species, as has been noted in whales (Hof and Van der
Gucht 2007).
The regional differences observed in elephant superficial
pyramidal neurons, with frontal neurons exhibiting more
complex dendritic systems than occipital neurons, are
generally consistent with findings in human supragranular
pyramidal neurons, where basilar dendrites in area 10 were
found to be more complex than those in area 18 (Jacobs
et al. 1997), and where greater dendritic/spine complexity
has been documented in heteromodal (areas 6b, 39) and
supramodal (areas 10, 11) cortices than in primary (areas
3-1-2, 4) and unimodal (areas 22, 44) cortices (Jacobs et al.
2001). Nevertheless, further research is required in the
elephant to determine if there are progressive increases in
basilar dendritic complexity along hierarchically arranged
pathways, as has been demonstrated in non-human primates
(Elston et al. 1996; Elston and Rosa 1997, 1998a, b), or if
regional differences in basilar dendritic extent are due to
intrinsic properties within each cortical region, as appears to
be the case in the mouse (Benavides-Piccione et al. 2006).
Finally, although TDL was similar between elephants
and humans, there was a difference in how this overall
length was distributed within the neuron. Human supra-
granular pyramidal neurons appeared smaller and denser,
with more complex branching patterns, whereas elephant
superficial pyramidal neurons appeared larger and more
expansive in terms of dendritic spread. Thus, although the
elephant brain is *3 times larger than the human brain,
elephant cortical neurons are not simply ‘‘scaled up’’ ver-
sions of a general mammalian type. As such, simple
models of neuron scaling and computations across phy-
logeny may be problematic (Wittenberg and Wang 2007).
Each species has its own phylogenetic heritage and func-
tional design principles (Harrison et al. 2002). Elephant
neuromorphology may reflect a particular phylogenetic
heritage and/or may be associated with the lower cell
packing density in the cortex. Indeed, the elephant cortex
appears to have a lower neuronal density than dolphins and
primates (Haug 1987; Lyck et al. 2009). These data,
combined with calculations of total volume of cortex
dedicated to somatic processing (Hofman 1982), suggest
that elephants have approximately 10 billion non-somatic
neurons spread across a large area of neocortex, compared
to 6.5 billion tightly packed neurons for chimpanzees and
20 billion tightly packed neurons for humans (Hart et al.
2008). As such, it has been suggested that elephant neo-
cortex may favor long-distance connections over the more
compartmentalized primate model, a possibility tentatively
supported by the longer basilar dendritic systems docu-
mented here.
Brain Struct Funct (2011) 215:273–298 293
123

Evolutionary and cognitive considerations
The present results confirm that the African elephant
cerebral cortex contains a rich variety of large neurons with
expansive, spine-rich dendritic systems and, presumably,
long axonal projections to interconnect these relatively
sparsely packed neurons (Wang et al. 2008). These findings
reinforce the high metabolic expense of large brains
(Grande 1980), and appear consistent with evidence in the
elephant neocortex for high glia-neuron ratios (Haug
1987), high neocortical white–gray matter ratios (Zhang
and Sejnowski 2000), and evolutionary selection on aero-
bic energy metabolism genes (Goodman et al. 2009).
Reasons for the extensive dendritic spread of elephant
neurons, as opposed to the more compact, branched pri-
mate model, remain to be explained.
Perhaps, the most distinguishing characteristic of the
elephant cortex is the presence of widely bifurcating apical
dendrites that branch close to the soma. This seems con-
sistent with the closest living relatives of elephants,
including other afrotherians and especially xenarthrans
(e.g., the giant anteater), but contrasts with findings in the
rock hyrax, where pyramidal neurons have single apical
dendrites resembling those in rodents and primates (Sher-
wood et al. 2009; Bianchi et al, in review). Variations of
bifurcating apical dendrites also appear in cetaceans (Ferrer
and Perera 1988; Hof et al. 1992). More generally, bifur-
cating apical branches appear in caniforms and perisso-
dactyls (Barasa 1960), which is consistent with the close
association of odd-toed ungulates and carnivores, and the
possible assemblage of carnivores and cetartiodactyls
together into superclade Ferungulata (Hou et al. 2009).
Thus, it is possible that bifurcating apical dendrites may be
a feature of animals with large bodies/brains, but to the
general exclusion of primates due to their position within
the euarchontoglire lineage. This raises the interesting
possibility that geometric effects on neocortical neuron
scaling are constrained by phylogenetic history.
Although the arrangement and morphology of cortical
neurons crucially interact with an elephant’s cognitive and
sociobehavioral repertoire, the relationship between corti-
cal architecture and cognition remains unclear. The pres-
ence of V-shaped apical bundles, high synaptic density,
and neurons with a wide, horizontal dendritic spread col-
lectively illustrate that neocortical neurons in elephants are
designed to synthesize a large number of diverse inputs.
Nevertheless, much more research on elephant neocortex is
required before any functional conclusions can be sup-
ported. Certainly, the elephant’s integrative cortical cir-
cuitry seems consistent with its complex spatial and social
abilities, as well as its behaviors indicating empathy and
theory-of-mind. These cortical characteristics may also
contribute to the elephant’s ability to synthesize a variety
of information over an extended period of time. In con-
clusion, although the present study provides an initial
window into the complex neuromorphology of the elephant
cortex, it also highlights the need for more comparative
research to elucidate the multiplicity of evolutionary
options for constructing large, elaborate brains.
Acknowledgments Partial support for this work was provided by the
Thomas M. McKee Professorship in the Natural Sciences (B.J.), The
Colorado College’s divisional research funds (B.J.), the James S. Mc-
Donnell Foundation (grant 22002078, to C.C.S., P.R.H.), and the South
African National Research Foundation (P.R.M.; FA2005033100004).
We would also like to thank Dr. Hilary Madzikanda of the Zimbabwe
Parks and Wildlife Management Authority, and Dr. Bruce Fivaz and the
team at the Malilangwe Trust, Zimbabwe.
References
Anderson K, Bones B, Robinson B, Hass C, Lee H, Ford K, Roberts
T-A, Jacobs B (2009) The morphology of supragranular
pyramidal neurons in the human insular cortex: a quantitative
Golgi study. Cereb Cortex 19:2131–2144
Anderson K, Yamamoto E, Kaplan J, Hannan M, Jacobs B (2010)
Neurolucida lucivid versus Neurolucida camera: a quantitative
and qualitative comparison of three-dimensional neuronal
reconstructions. J Neurosci Methods 186:209–214
Badlangana NL, Bhagwandin A, Fuxe K, Manger PR (2007)
Observations on the giraffe central nervous system related to
the corticospinal tract, motor cortex and spinal cord: what
difference does a long neck make? Neuroscience 148:522–534
Barasa A (1960) Forma, grandezza e densita dei neuroni della
corteccia cerebrale in mammiferi di grandezza corporea differ-
ente. Z Zellforsch 53:69–89
Barasa A, Shochatovitz A (1961) Grandezza e densita della cellule
nervose della corteccia cerebrale de Elephas indicus. Rendiconti
Accad Naz Lincei 30:246–249
Barbas H (1995) Anatomic basis of cognitive-emotional interactions
in the primate prefrontal cortex. Neurosci Biobehav Rev
19:499–510
Bartesaghi R, Severo S, Guidi S (2003) Effects of early environment
on pyramidal neuron morphology in field CA1 of the guinea-pig.
Neuroscience 116:715–732
Bates LA, Sayialel KN, Njiraini NW, Moss CJ, Poole JH, Byrne RW
(2007a) Elephants classify human ethnic groups by odor and
garment color. Curr Biol 17:1938–1942
Bates LA, Sayialel KN, Njiraini NW, Poole JH, Moss CJ, Byrne RW
(2007b) African elephants have expectations about the locations
of out-of-site family members. Biol Lett 4:34–36
Bates LA, Lee PC, Njiraini NW, Poole JH, Sayialel KN, Sayialel S,
Sayialel S, Moss CJ, Byrne RW (2008a) Do elephants show
empathy? J Consc Stud 15:204–225
Bates LA, Poole JH, Byrne RW (2008b) Elephant cognition. Curr
Biol 18:R544–R546
Benavides-Piccione R, Hamzei-Sichani F, Ballesteros-Yanez I,
DeFelipe J, Yuste R (2006) Dendritic size of pyramidal neurons
differs among mouse cortical regions. Cereb Cortex 16:990–
1001
Bianchi S, Bauernfeind AL, Stimpson CD, Bonar CJ, Manger PR, Hof
PR, Jacobs B, Sherwood CC Neuronal diversity in Afrotheria: a
Golgi study of the rock hyrax (Procavia capensis) neocortex and
comparison with the African elephant (Loxodonta africana). NY
Acad Sci (in review)
294 Brain Struct Funct (2011) 215:273–298
123

Bok ST (1959) Histonomy of the cerebral cortex. Elsevier,
Amsterdam
Bota M, Swanson LW (2007) The neuron classification problem.
Brain Res Rev 56:79–88
Braak H, Braak E (1976) The pyramidal cells of Betz within the
cingulate and precentral gigantopyramidal field in the human
brain: a Golgi and pigmentarchitectonic study. Cell Tissue Res
172:103–119
Braak H, Braak E (1985) Golgi preparations as a tool in neuro-
pathology with particular reference to investigations of the
human telencephalic cortex. Prog Neurobiol 25:93–139
Brown KM, Gillette TA, Ascoli GA (2008) Quantifying neuronal
size: summing up trees and splitting the branch difference.
Semin Cell Dev Biol 19:485–493
Bueno-Lopez JL, Reblet C, Lopez-Medina A, Gomez-Urquijo
Grandes P, Gondra J, Hennequet L (1991) Targets and laminar
distribution of projection neurons with ‘‘inverted’’ morphology
in rabbit cortex. Eur J Neurosci 3:415–430
Butti C, Hof PR (2010) The insular cortex: a comparative perspective.
Brain Struct Func 214:477–493
Butti C, Sherwood CC, Hakeem AY, Allman JM, Hof PR (2009)
Total number and volume of von Economo neurons in the
cerebral cortex of cetaceans. J Comp Neurol 515:243–259
Constantinidis C, Williams GV, Goldman-Rakic PS (2002) A role for
inhibition in shaping the temporal flow of information in
prefrontal cortex. Nat Neurosci 5:175–180
Cozzi B, Spagnoli S, Bruno L (2001) An overview of the central
nervous system of the elephant through a critical appraisal of the
literature published in the XIX and XX centuries. Brain Res Bull
54:219–227
de Crinis M (1934) Uber die Spezialzellen in der menschlichen
Grosshirnrinde. J Psych Neurol. 45:439–449
de Lima AD, Voigt T, Morrison JH (1990) Morphology of the cells
within the inferior temporal gyrus that project to the prefrontal
cortex in the macaque monkey. J Comp Neurol 296:159–172
de Ruiter JP (1983) The influence of post-mortem fixation delay on
the reliability of the Golgi silver impregnation. Brain Res
266:143–147
DeFelipe J (1997) Types of neurons, synaptic connections and
chemical characteristics of cells immunoreactive for calbindin—
D28K, parvalbumin and calretinin in the neocortex. J Chem
Neuroanat 14:1–19
DeFelipe J, Alonso-Nanclares L, Arellano JI (2002) Microstructure of
the neocortex: comparative aspects. J Neurocytol 31:299–316
Druga R (2009) Neocortical inhibitory system. Folia Biol (Prague)
55:201–217
Elston GN (2003) Cortex, cognition and the cell: new insights into the
pyramidal neuron and prefrontal function. Cereb Cortex
13:1124–1138
Elston GN (2007) Specialization of the neocortical pyramidal cell
during primate evolution. In: Kass JH, Preuss TM (eds)
Evolution of nervous systems: a comprehensive reference, vol
4. Elsevier, New York, pp 191–242
Elston GN, Rosa MGP (1997) The occipitoparietal pathway of the
macaque monkey: comparison of pyramidal cell morphology in
layer III of functionally related cortical visual areas. Cereb
Cortex 7:432–452
Elston GN, Rosa MGP (1998a) Complex dendritic fields of pyramidal
cells in the frontal eye field of the macaque monkey: comparison
with parietal areas 7a and LIP. NeuroReport 9:127–131
Elston GN, Rosa MGP (1998b) Morphological variation of layer III
pyramidal neurones in the occipitotemporal pathway of the
macaque monkey visual cortex. Cereb Cortex 8:278–294
Elston GN, Rosa MGP, Calford MB (1996) Comparison of dendritic
fields of layer III pyramidal neurons in striate and extrastriate
visual areas of the marmoset: a Lucifer Yellow intracellular
injection study. Cereb Cortex 6:807–813
Ferrer I (1986) Golgi study of the isocortex in an insectivore: the
common European mole (Talpa europaea). Brain Behav Evol
29:105–114
Ferrer I (1987) The basic structure of the neocortex in insectivorous
bats (Miniopterus sthreibersi and Pipistrellus pipistrellus).
J Hirnforsch 28:237–243
Ferrer I, Perera M (1988) Structure and nerve cell organisation in the
cerebral cortex of the dolphin Stenella coeruleoalba a Golgi
study: with special attention to the primary auditory area. Anat
Embryol (Berl) 178:161–173
Ferrer I, Fabregues I, Condom E (1986a) A Golgi study of the sixth
layer of the cerebral cortex I. The lissencephalic brain of
Rodentia, Lagomorpha, Insectivora and Chiroptera. J Anat
145:217–234
Ferrer I, Fabregues I, Condom E (1986b) A Golgi study of the sixth
layer of the cerebral cortex II. The gyrencephalic brain of
Carnivora, Artiodactyla and primates. J Anat 146:87–104
Ferrer I, Fabregues I, Condom E (1987) A Golgi study of the sixth
layer of the cerebral cortex III. Neuronal changes during normal
and abnormal cortical folding. J Anat 152:71–82
Fitzpatrick DC, Henson OW (1994) Cell types in the mustached bat
auditory cortex. Brain Behav Evol 43:79–91
Fleischhauer K (1974) On different patterns of dendritic bundling in
the cerebral cortex of the cat. Z Anat Entwickl-Gesch
143:115–126
Fleischhauer K, Petsche H, Wittkowski W (1972) Vertical bundles of
dendrites in the neocortex. Anat Embryol 136:213–223
Foxe JJ, Simpson GV (2002) Flow of activation from V1 to frontal
cortex in humans. Exp Brain Res 142:139–150
Garey LJ, Winkelmann E, Brauer K (1985) Golgi and Nissl studies of
the visual cortex of the bottlenose dolphin. J Comp Neurol
240:305–321
Garstang M (2004) Long-distance, low-frequency elephant commu-
nication. J Comp Physiol A 190:791–805
Geisler HC, Ijkema-Paassen J, Westerga J, Gramsbergen A (2000)
Vestibular deprivation and the development of dendritic bundles
in the rat. Neural Plast 7:193–203
Germroth P, Schwerdtfeger WK, Buhl EH (1989) Morphology of
identified entorhinal neurons projecting to the hippocampus A
light microscopical study combining retrograde tracing and
intracellular injection. Neuroscience 30:683–691
Gheerbrant E, Tassy P (2009) L’origine et l’evolution des elephants.
C R Palevol 8:281–294
Glezer II, Morgane PJ (1990) Ultrastructure of synapses and Golgi
analysis of neurons in neocortex of the lateral gyrus (visual
cortex) of the dolphin and pilot hale. Brain Res Bull 24:401–
427
Glezer II, Jacobs MS, Morgane PJ (1988) Implications of the ‘‘initial
brain’’ concept for brain evolution in Cetacea. Behav Brain Sci
11:75–116
Goodman M, Sterner KN, Islam M, Uddin M, Sherwood CC, Hof PR,
Hou Z-C, Lipovich L, Jia H, Grossman LI, Wildman DE (2009)
Phylogenomic analyses reveal convergent patterns of adaptive
evolution in elephant and human ancestries. Proc Natl Acad Sci
USA 106:20824–20829
Grande F (1980) Energy expenditure of organs and tissues. In: Kinney
JM (ed) Assessment of energy metabolism in health and disease:
report of the first Ross conference on medical research. Ross
Laboratories, Columbus, pp 88–92
Gravett N, Bhagwandin A, Fuxe K, Manger PR (2009) Nuclear
organization and morphology of cholinergic, putative catechol-
aminergic and serotonergic neurons in the brain of the rock
hyrax, Procavia capensis. J Chem Neuroanat 38:57–74
Brain Struct Funct (2011) 215:273–298 295
123

Group Petilla Interneuron Nomenclature (2008) Petilla terminology:
nomenclature of features of GABAergic interneurons of the
cerebral cortex. Nat Rev Neurosci 9:557–568
Hakeem AY, Hof PR, Sherwood CC, Switzer RC, Rasmussen LRL,
Allman JM (2005) Brain of the African elephant (Loxodontaafricana): neuroanatomy from magnetic resonance images. Anat
Rec 287A:1117–1127
Hakeem AY, Sherwood CC, Bonar CJ, Butti C, Hof PR, Allman JM
(2009) Von Economo neurons in the elephant brain. Anat Rec
292:242–248
Harrison KH, Hof PR, Wang SS-H (2002) Scaling laws in the
mammalian neocortex: does form provide blues to function? J
Neurocytol 31:289–298
Hart BL, Hart LA (2007) Evolution of the elephant brain: a paradox
between brain size and cognitive behavior. In: Kaas JH,
Krubitzer LA (eds) Evolution of nervous systems: a compre-
hensive reference. Vol 3: Mammals. Elsevier, Amsterdam (NL),
pp 491–497
Hart BL, Hart LA (2010) Unique attributes of the elephant mind offer
perspectives on the human mind. In: Smith J, Mitchell RW (eds)
Experiencing animals: encounters between animals and human
minds. Columbia University Press, New York (in press)
Hart BL, Hart LA, McCoy M, Sarath CR (2001) Cognitive behaviour
in Asian elephants: use and modification of branches for fly
switching. Anim Behav 62:839–847
Hart BL, Hart LA, Pinter-Wollman N (2008) Large brains and
cognition: where do elephants fit in? Neurosci Biobehav Rev
32:86–98
Hassiotis M, Ashwell KWS (2003) Neuronal classes in the isocortex
of a monotreme, the Australian echidna (Tachyglossus aculea-tus). Brain Behav Evol 61:6–27
Hassiotis M, Paxinos G, Ashwell KWS (2003) The anatomy of the
cerebral cortex of the echidna (Tachyglossus aculeatus). Comp
Biochem Physiol Part A 136:827–850
Hassiotis M, Paxinos G, Ashwell KWS (2005) Cyto- and chemoar-
chitecture of the cerebral cortex of an echidna (Tachyglossusaculeatus) II. Laminar organization and synaptic density.
J Comp Neurol 482:94–122
Haug H (1970) Die makrospkoschie Aufbau des Großhirns: qualita-
tive und quantitative Untersuchungen an den Gehirnen des
Menschen der Delphinoideae und des Elefanten. Springer, Berlin
Haug H (1987) Brain sizes, surfaces, and neuronal sizes of the cortex
cerebri: a stereological investigation of man and his variability
and a comparison with some mammals (primates, whales,
marsupials, insectivores, and one elephant). Am J Anat
180:126–142
Hof PR, Sherwood CC (2005) Morphomolecular neuronal phenotypes
in the neocortex reflect phylogenetic relationships among certain
mammalian orders. Anat Rec 287A:1153–1163
Hof PR, Van der Gucht E (2007) Structure of the cerebral cortex of
the humpback whale, Megaptera novaeangliae (Cetacea, Mys-
ticeti, Balaeopteridae). Anat Rec 290:1–31
Hof PR, Glezer II, Archin N, Janssen WG, Morgane PJ, Morrison JH
(1992) The primary auditory cortex in cetacean and human
brain: a comparative analysis of neurofilament protein-contain-
ing pyramidal neurons. Neurosci Lett 146:91–95
Hof PR, Bogaert YE, Rosenthal RE, Fiskum G (1996) Distribution of
neuronal populations containing neurofilament protein and
calcium-binding proteins in the canine neocortex: regional
analysis and cell typology. J Chem Neuroanat 11:81–98
Hof PR, Glezer II, Conde F, Flagg RA, Rubin MB, Nimchinsky EA,
Vogt Weisenhorn DM (1999) Cellular distribution of the
calcium-binding proteins parvalbumin, calbindin, and calretinin
in the neocortex of mammals: phylogenetic and developmental
patterns. J Chem Neuroanat 16:77–116
Hof PR, Chanis R, Marino L (2005) Cortical complexity in cetacean
brains. Anat Rec 287A:1142–1152
Hoffmann JN, Montag AG, Dominy NJ (2004) Meissner corpuscles
and somatosensory acuity: the prehensile appendages of primates
and elephants. Anat Rec 281A:1138–1147
Hofman MA (1982) Encephalization in mammals in relation to the
size of the cerebral cortex. Brain Behav Evol 20:84–96
Horner CH, Arbuthnott E (1991) Methods of estimation of spine
density—are spines evenly distributed throughout the dendritic
field? J Anat 177:179–184
Hou ZC, Romero R, Wildman DE (2009) Phylogeny of the
Ferungulata (Mammalia: Laurasiatheria) as determined from
phylogenomic data. Mol Phylogenet Evol 52:660–664
Innocenti GM, Vercelli A (2010) Dendritic bundles, minicolumns,
columns, and cortical output units. Front Neuroanat 4:1–7
Jacobs B, Scheibel AB (1993) A quantitative dendritic analysis of
Wernicke’s area in humans I. Lifespan changes. J Comp Neurol
327:83–96
Jacobs B, Scheibel AB (2002) Regional dendritic variation in primate
cortical pyramidal cells. In: Schuz A, Miller R (eds) Cortical
areas: unity and diversity (Conceptual advances in brain research
series). Taylor & Francis, London, pp 111–131
Jacobs B, Schall M, Scheibel AB (1993) A quantitative dendritic
analysis of Wernicke’s area in humans II. Gender, hemispheric,
and environmental factors. J Comp Neurol 327:97–111
Jacobs B, Driscoll L, Schall M (1997) Life-span dendritic and spine
changes in areas 10 and 18 of human cortex: a quantitative Golgi
study. J Comp Neurol 386:661–680
Jacobs B, Schall M, Prather M, Kapler E, Driscoll L, Baca S (2001)
Regional dendritic and spine variation in human cerebral cortex:
a quantitative Golgi study. Cereb Cortex 11:558–571
Jones EG (1984) Neurogliaform or spiderweb cells. In: Peters A,
Jones EG (eds) Cerebral cortex. Plenum Press, New York and
London, pp 409–418
Juba A (1934) Uber seltenere Ganglienzellformen der Grosshirnrinde.
Z Zellforsch 21:441–447
Kaiserman-Abramof IR, Peters A (1972) Some aspects of the
morphology of Betz cells in the cerebral cortex of the cat. Brain
Res 43:527–546
Kawaguchi Y (1995) Physiological subgroups of nonpyramidal cells
with specific morphological characteristics in layer II/III of rat
frontal cortex. J Neurosci 15:2638–2665
Kesarev VS (1975) Homologization of the cerebral neocortex in
cetaceans. Arkh Anat Gistol Embriol 68:5–13
Kisvarday ZF, Gulyas A, Beroukas D, North JB, Chub IW, Somogyi P
(1990) Synapses, axonal and dendritic patterns of GABA-immu-
noreactive neurons in human cerebral cortex. Brain 113:793–812
Kupsky WJ, Marchant GH, Cook K, Shoshani J (2001) Morphologic
analysis of the hippocampal formation in Elephas maximus and
Loxodonta africana with comparison to that of human. In:
Cavarretta G, Gioia P, Mussi M, Palombo MR (eds) Proceedings
of the first international congress of La Terra degli Elefanti, the
world of elephants. Consiglio Nazionale delle Ricerche, Rome
(IT), pp 643–647
Lubke J, Egger V, Sakmann B, Feldmeyer D (2000) Columnar
organization of dendrites and axons of single and synaptically
coupled excitatory spiny neurons in layer 4 of the rat barrel
cortex. J Neurosci 20:5300–5311
Lubke J, Roth A, Feldmeyer D, Sakmann B (2003) Morphometric
analysis of the columnar innervation domain of neurons
connecting layer 4 and layer 2/3 of juvenile rat barrel cortex.
Cereb Cortex 13:1051–1063
Lund JS, Lewis DA (1993) Local circuit neurons of developing
mature macaque prefrontal cortex: Golgi and immunocytochem-
ical characteristics. J Comp Neurol 328:282–312
296 Brain Struct Funct (2011) 215:273–298
123

Lyck L, Dalmau Santamaria I, Pakkenberg B, Chemnitz J, Daa
Schrøder H, Finsen B, Gundersen HJG (2009) An empirical
analysis of the precision of estimating the numbers of neurons
and glia in human neocortex using a fractionator-design with
sub-sampling. J Neurosci Methods 182:143–156
Manger PR, Sum M, Szymanski M, Ridgway S, Krubitzer L (1998)
Modular subdivisions of dolphin insular cortex: does evolution-
ary history repeat itself? J Cogn Neurosci 10:153–166
Manger PR, Cort J, Ebrahim N, Goodman A, Henning J, Karolia M,
Rodrigues S-L, Strkalj G (2008) Is 21st century neuroscience too
focused on the rat/mouse model of the brain function and
dysfunction? Front Neuroanat 2:1–7
Manger PR, Pillay P, Madeko BC, Bhagwandin A, Gravett N, Moon
D-J, Jillani N, Hemingway J (2009) Acquisition of brains from
the African elephant (Loxodonta africana): Perfusion-fixation
and dissection. J Neurosci Methods 179:16–21
Manger PR, Hemingway J, Haagensen M, Gilissen E (2010) Cross-
sectional area of the elephant corpus callosum: comparison to
other eutherian mammals. Neuroscience 167:815–824
Markowitz H, Schmidt M, Nadal L, Squire L (1975) Do elephants
ever forget? J Appl Behav Anal 8:333–335
Masland RH (2004) Neuronal cell types. Curr Biol 14:R497–R500
McComb K, Moss C, Sayialel S, Baker L (2000) Unusually extensive
networks of vocal recognition in African elephants. Anim Behav
59:103–1109
McComb K, Moss C, Durant SM, Baker L, Sayialel S (2001)
Matriarchs as repositories of social knowledge in African
elephants. Science 292:491–494
Mendizabal-Zubiaga JL, Reblet C, Bueno-Lopez JL (2007) The
underside of the cerebral cortex: layer V/VI spiny inverted
neurons. J Anat 211:223–236
Meyer G (1987) Forms and spatial arrangement of neurons in the
primary motor cortex of man. J Comp Neurol 262:402–428
Miller MW (1988) Maturation of rat visual cortex: IV. The
generation, migration, morphogenesis and connectivity of atyp-
ically oriented pyramidal cells. J Comp Neurol 274:387–405
Morest DK, Morest RR (2005) Perfusion-fixation of the brain with
chrome-osmium solutions for the rapid Golgi method. Am J
Anat 118:811–831
Mountcastle VB (1997) The columnar organization of the neocortex.
Brain 120:701–722
Ngowyang G (1932) Beschreibung einer Art von Specialzellen in der
Inselrinde. J Psychol Neurol 44:671–674
Ngowyang G (1936) Neuere Befunde uber die Gabelzellen.
Z Zellforsch 25:236–239
Nimchinsky EA, Vogt BA, Morrison JH, Hof PR (1995) Spindle
neurons of the human anterior cingulate cortex. J Comp Neurol
355:27–37
Ong WY, Garey LJ (1990) Neuronal architecture of the human
temporal cortex. Anat Embryol 181:351–364
Parnavelas JG, Lieberman AR, Webster KE (1977) Organization of
neurons in the visual cortex, area 17, of the rat. J Anat 124:
305–322
Peters A, Regidor J (1981) A reassessment of the forms of
nonpyramidal neurons in area 17 of cat visual cortex. J Comp
Neurol 203:685–716
Pieters R, Gravett N, Fuxe K, Manger PR (2010) Nuclear organiza-
tion of cholinergic, putative catecholaminergic and serotonergic
nuclei in the brain of the eastern rock elephant shrew,
Elephantulus myurus. J Chem Neuroanat 39:175–188
Plotnik JM, de Waal FBM, Reiss D (2006) Self-recognition in an
Asian elephant. Proc Natl Acad Sci USA 103:17053–17057
Poole JH (1999) Signals and assessment in African elephants:
evidence from playback experiments. Anim Behav 58:185–193
Poole J, Moss CJ (2008) Elephant sociality and complexity: the
scientific evidence. In: Wemmer CC (ed) Elephants and ethics:
towards a morality of coexistence. John Hopkins University
Press, Baltimore, pp 69–98
Poole JH, Tyack PL, Stoeger-Horwath AS, Watwood S (2005)
Elephants are capable of vocal learning: two animals coin
unexpected sounds as a surprising form of social communica-
tion. Nature 434:455–456
Povysheva NV, Zaitsev AV, Kroner S, Krimer OA, Rotaru DC,
Gonzalez-Burgos G, Lewis DA, Krimer LS (2007) Electrophys-
iological differences between neurogliaform cells from monkey
and rat prefrontal cortex. J Neurophysiol 97:1030–1039
Qi H-X, Jain N, Preuss TM, Kaas JH (1999) Inverted pyramidal
neurons in chimpanzee sensorimotor cortex are revealed by
immunostaining with monoclonal antibody SMI-32. Somatosens
Mot Res 16:49–56
Ramon y, Cajal S (1891) On the structure of the cerebral cortex of
certain mammals. (La Cellule 7:125–176). In: DeFelipe J, Jones
EG (trans. and eds) Cajal on the cerebral cortex: an anno-
tated translation on the complete writings (History of neurosci-
ence; No 1) (1988) Oxford University Press, New York,
pp 23–54
Ramon y Cajal S (1922) Studien uber die Sehrinde der Katze.
J Psychol Neurol 29:161–181
Roney KJ, Scheibel AB, Shaw GL (1979) Dendritic bundles: survey
of anatomical experiments and physiological theories. Brain Res
Rev 1:225–271
Roth G, Dicke U (2005) Evolution of the brain and intelligence.
Trends Cogn Sci 9:250–257
Sanides F, Sanides D (1972) The ‘‘extraverted neurons’’ of the
mammalian cerebral cortex. Z Anat Entwickl-Gesch
136:272–293
Scheibel ME, Scheibel AB (1975) Dendrite bundles, central programs
and the olfactory bulb. Brain Res 95:407–421
Scheibel ME, Scheibel AB (1978a) The dendritic structure of the
human Betz cell. In: Brazier MAB, Petsche H (eds) Architec-tonics of the cerebral cortex (IBRO) monographic series, vol 3.
Raven Press, New York, pp 43–57
Scheibel ME, Scheibel AB (1978b) The methods of Golgi. In:
Robertson RT (ed) Neuroanatomical research techniques. Aca-
demic Press, New York, pp 89–114
Schlaug G, Armstrong E, Schleicher A, Zilles K (1993) Layer V
pyramidal cells in the adult human cingulate cortex: a quanti-
tative Golgi-study. Anat Embryol 187:515–522
Schmolke C, Kunzle H (1997) On the presence of dendrite bundles in
the cerebral cortex of the Madagascan lesser hedgehog tenrec
and the red-eared pond turtle. Anat Embryol 196:195–213
Sherwood CC, Lee PWH, Rivara C-B, Holloway RL, Gilissen EPE,
Simmons RMT, Hakeem A, Allman JM, Erwin JM, Hof PR
(2003) Evolution of specialized pyramidal neurons in primate
visual and motor cortex. Brain Behav Evol 61:28–44
Sherwood CC, Stimpson CD, Butti C, Bonar CJ, Newton AL, Allman
JM, Hof PR (2009) Neocortical neuron types in Xenarthra and
Afrotheria: implications for brain evolution in mammals. Brain
Struct Funct 213:301–328
Sholl DA (1953) Dendritic organization of the neurons of the visual
and motor cortices of the cat. J Anat 87:387–406
Shoshani J, Tassy P (2005) Advances in proboscidean taxonomy and
classification, anatomy and physiology, and ecology and behav-
ior. Quat Int 126–128:5–20
Shoshani J, Kupsky WJ, Marchant GH (2006) Elephant brain part I:
gross morphology, functions, comparative anatomy, and evolu-
tion. Brain Res Bull 70:124–157
Soltis J (2009) Vocal communication in African elephants (Loxodontaafricana). Zoo Biol 28:1–18
Somogyi P, Kisvarday ZF, Martin KAC, Whitteridge D (1983)
Synaptic connections of morphologically characterized large
basket cells in the striate cortex of cat. Neurosci 10:261–294
Brain Struct Funct (2011) 215:273–298 297
123

Sukumar R (2003) The living elephants: evolutionary ecology
behavior, and conservation. Oxford University Press, New York
Syring A (1956) Die Verbreitung von Spezialzellen in der Gross-
hirnrinde verschiedener Saugertiergruppen. Z Zellforsch
45:399–434
Tower DB (1954) Structural and functional organization of mamma-
lian cerebral cortex; the correlation of neurone density with brain
size; cortical neurone density in the fin whale (Balaenopteraphysalus L.) with a note on the cortical neurone density in the
Indian elephant. J Comp Neurol 101:19–51
Tyler CJ, Dunlop SA, Lund RD, Harman AM, Dann JF, Beazley LD
(1998) Anatomical comparison of the macaque and marsupial
visual cortex: common features that may reflect retention of
essential cortical elements. J Comp Neurol 400:449–468
Uylings HBM, Ruiz-Marcos A, van Pelt J (1986) The metric analysis
of three-dimensional dendritic tree patterns: a methodological
review. J Neurosci Methods 18:127–151
Valverde F, Facal-Valverde MV (1986) Neocortical layers I and II of
the hedgehog (Erinaceus europaeus). I. Intrinsic organization.
Anat Embryol 173:413–430
Van Brederode JFM, Foehring RC, Spain WJ (2000) Morphological
and electrophysiological properties of atypically oriented layer 2
pyramidal cells of the juvenile rat neocortex. Neuroscience
101:851–861
Van der Loos H (1965) The ‘‘improperly’’ oriented pyramidal cell in
the cerebral cortex and its possible bearing on problems of
neuronal growth and cell orientation. Bull Johns Hopkins Hosp
117:228–250
von Economo CF, Koskinas GN (1925) Die cytoarchitektonik der
Hirnrinde des erwachsenen Menschen. Springer, Berlin
Wang SS-H, Shultz JR, Burish MJ, Harrison KH, Hof PR, Towns LC,
Wagers MW, Wyat KD (2008) Functional trade-offs in white
matter axonal scaling. J Neurosci 28:4047–4056
Wildman DE, Uddin M, Opazo JC, Liu G, Lefort V, Guindon S,
Gascuel O, Grossman LI, Romero R, Goodman M (2007)
Genomics, biogeography, and the diversification of placental
mammals. Proc Natl Acad Sci USA 104:14395–14400
Wittenberg GM, Wang SS-H (2007) Evolution and scaling of
dendrites. In: Stuart G, Spruston N, Hausser M (eds) Dendrites.
Oxford University Press, New York, pp 43–67
Wyss JM, Van Groen T, Sripanidkulchai K (1990) Dendritic bundling
in layer I of granular retrosplenial cortex: intracellular labeling
and selectivity of innervation. J Comp Neurol 295:33–42
Yamamoto T, Samejima A, Oka H (1987) Morphological features of
layer V pyramidal neurons in the cat parietal cortex: an
intracellular HRP study. J Comp Neurol 265:380–390
Zeitsev AV, Povysheva NV, Gonzalez-Burgos G, Rotaru D, Fish KN,
Krimer LS, Lewis DA (2009) Interneuron diversity in layers 2–3
of monkey prefrontal cortex. Cereb Cortex 19:1597–1615
Zhang K, Sejnowski TJ (2000) A universal scaling law between gray
matter and white matter of cerebral cortex. Proc Natl Acad Sci
USA 97:5621–5626
298 Brain Struct Funct (2011) 215:273–298
123