no te - jst
TRANSCRIPT

Endocrinol. Japon. 1970, 17 (1), 89~92
NO TE
The Mechanism of Insulin Action on Hexokinase Isozymes
KOKO MURAKAMI* AND SADAHIKO ISHIBASHI*
Department of Physiological Chemistry, Fuculty ofPharmaceutical Sciences, University of Tokyo, Tokyo
Synopsis
The mechanism of insulin action on hexokinase isozymes was examined by theelectrophoresis on cellulose acetate membrane. It is demonstrated that insulin, some(receptor ?) protein and type II may form a complex sensitive to sulfhydryl compounds,which appears as type III in some tissues, or as the abortive one with disappearance oftype II in others. On this mechanism insulin was deduced to act to induce the protein forthe complex as well as to form this complex as a constituent.
Many investigations have been reportedabout the effect of insulin on hexokinase (E.C. 2. 7. 1. 1.) with controversial results
(Katzen 1967, McLean et al., 1966). Theauthors examined the mechanism of the actionof insulin on hexokinase for the level of itsconsisting isozymes, mainly using proliferat-ing granuloma in rats.
Materials and Methods
Granuloma was formed by the injection of 4mlof 2% carrageenin solution to the dorsal region ofrats (Donryu strains, 6 weeks old). It was so prolifera-tive tissue as to reach 5-6 g. in wet-weight after 5-6days. Granuloma as well as other tissues, was homo-genized and the enzyme extract was obtained after thecentrifugation at 105,000 g. for 30 min. Hexokinaseisozymes were separated with the electrophoresis oncellulose acetate membrane (Cellogel, Italy) by thetechnique of Sato et al. (1969) and detected accordingto the method of Katzen (1967). In this seriesisozymes were developed in the substrate concentra-tion of 10-2M. The patterns obtained were consistentwith those by the electrophoresis on starch gel. In theexperiment with mercaptoethanol and EDTA, enzyme
extracts were prepared and electrophoresis performed
in the presence of 5•~10-3M mercaptoethanol and
2.7•~10-3M EDTA.
Results
Hexokinase in granuloma consisted of type
I, II and III, which showed a specific pattern
that changed with a consistent tendency ac-
cording to the tissue formation. That is, in
the early proliferative phase type III was
predominent and type II was absent (Fig. 1).
This type III was consistent with that of spleen
and liver. Afterwards around the 5th day of
the carrageenin injection type III was gradual-
ly replaced by type II (Fig. 2). From these
patterns, some switch mechanism was sup-
posed between type III and type II. On the
other hand, type I had a constant activity
throughout the patterns.
The mechanism of the insulin action on
hexokinase isozymes was examined from the
change of these patterns in granuloma. As
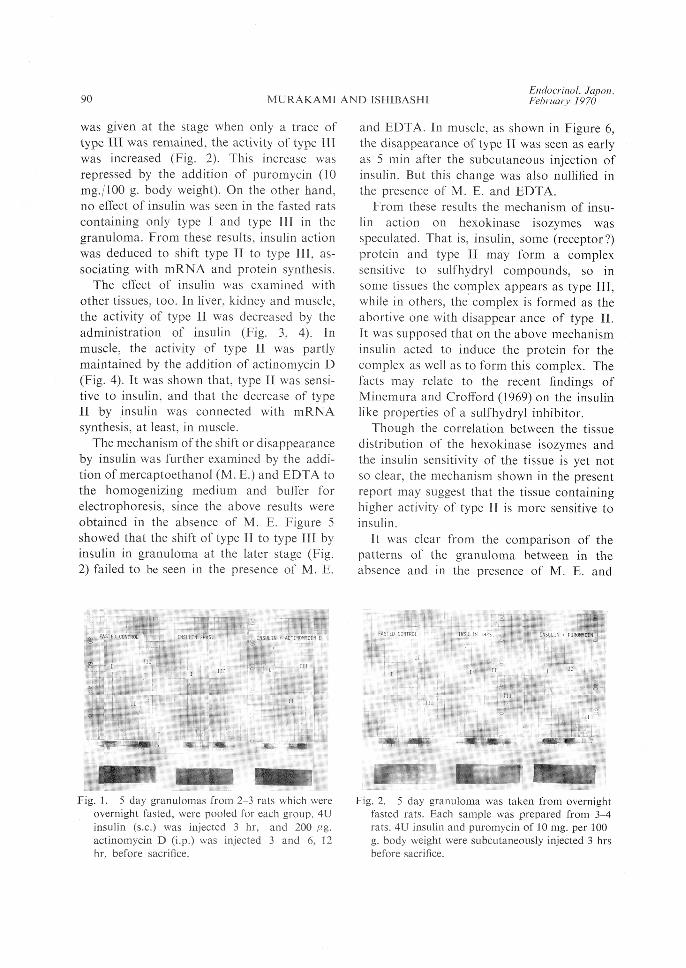
shown in Figure 1, when fasted rats containing
a trace of type II in the granuloma was sub-
cutaneously injected with 4U insulin, the
activity of type II was disappeared. By the
administration of 200 ƒÊg. actinomycin D, this
change by insulin was canceled. When insulin
Received for publication February 20, 1970.* Present address: Department of Physiological
Chemistry, Hiroshima University School of
Medicine, Hiroshima.
90 MURAKAMI AND ISHIBASHIEndocrinol.Japon.
February 1970
was given at the stage when only a trace oftype III was remained, the activity of type IIIwas increased (Fig. 2). This increase wasrepressed by the addition of puromycin (10mg./100 g. body weight). On the other hand,no effect of insulin was seen in the fasted ratscontaining only type I and type III in the
granuloma. From these results, insulin actionwas deduced to shift type II to type III, as-sociating with mRNA and protein synthesis.
The effect of insulin was examined withother tissues, too. In liver, kidney and muscle,the activity of type II was decreased by theadministration of insulin (Fig. 3, 4). Inmuscle, the activity of type II was partlymaintained by the addition of actinomycin D
(Fig. 4). It was shown that, type II was sensi-tive to insulin, and that the decrease of typeII by insulin was connected with mRNAsynthesis, at least, in muscle.
The mechanism of the shift or disappearanceby insulin was further examined by the addi-tion of mercaptoethanol (M. E.) and EDTA tothe homogenizing medium and buffer forelectrophoresis, since the above results wereobtained in the absence of M. E. Figure 5showed that the shift of type II to type III byinsulin in granuloma at the later stage (Fig.2) failed to be seen in the presence of M. E.
Fig. I. 5 day granulomas from 2-3 rats which wereovernight fasted, were pooled for each group. 4Uinsulin (s.c.) was injected 3 hr, and 200 pig.actinomycin D (i.p.) was injected 3 and 6, 12hr, before sacrifice.
and EDTA. In muscle, as shown in Figure 6,the disappearance of type II was seen as earlyas 5 min after the subcutaneous injection ofinsulin. But this change was also nullified inthe presence of M. E. and EDTA.
From these results the mechanism of insu-lin action on hexokinase isozymes was
speculated. That is, insulin, some (receptor ?)
protein and type II may form a complexsensitive to sulfhydryl compounds, so insome tissues the complex appears as type III,while in others, the complex is formed as theabortive one with disappear ance of type II.It was supposed that on the above mechanisminsulin acted to induce the protein for thecomplex as well as to form this complex. Thefacts may relate to the recent findings ofMinemura and Crofford (1969) on the insulinlike properties of a sulfhydryl inhibitor.
Though the correlation between the tissuedistribution of the hexokinase isozymes andthe insulin sensitivity of the tissue is yet notso clear, the mechanism shown in the presentreport may suggest that the tissue containinghigher activity of type II is more sensitive toinsulin.
It was clear from the comparison of the
patterns of the granuloma between in theabsence and in the presence of M. E. and
Fig. 2. 5 day granuloma was taken from overnightfasted rats. Each sample was prepared from 3-4rats. 4U insulin and puromycin of 10 mg. per 100
g. body weight were subcutaneously injected 3hrsbefore sacrifice.

INSUIN ACTION ON HEXOKINASE ISOZYMES 91
Fig. 3. Each group was poo ed from 5 rats which were overnight fasted.4U insu in wassubcutaneous y injected 2.5 hr before sacrifice.
Fig. 4. Each sample was prepared from 3-4 over-
night fasted rats. 4U insulin was subcutaneously
injected 3 hr before sacrifice. 200ƒÊg. actinomycin
D was intraperitoneally administered 12 and 6hr
before sacrifice.
EDTA, that this change by insulin was notbetween the two interconvertible forms oftype II as reported by Katzen (1967), but wasdeduced to be the shift between type II andIII.
The disappearance by insu in of type II inmusc e is not in conf ict with the conceptthat insulin is necessary for the maintenanceof type II (Katzen 1967), but may be inter-
preted that the interaction between the com-plex formation by insu in with type II and itsdissociation by M. E. might be contro ing the
physiological state of hexokinase isozymes.
Fig. 5. Granuloma was taken 8 days after car-rageenin injection from two rats. 4U insulin wassubcutaneously injected 3 hr before sacrifice.
The level of insulin might be a factor, since
in the present report insulin was given in
overnight fasted rats, not in diabetic rats as
used in many reports.
Acknow edgment
Many thanks must be expressed to Drs. M.
Yamada and S. Tsurufuji for their kindest advices.

92 MURAKAMI AND ISHIBASHIEndocrinol. Japon.February 1970
Fig. 6. Overnight fasted rats were subcutaneouslyinjected with 4U insulin 5 mins. before sacrifice.The group with M.E. was prepared from 2 rats,that without M.E. from 5 rats.
References
Katzen H. M. Adv. Enz. Reg. Vol. 5, p.335
(1967).McLean P., J. Brown, K. Greenslade and K.
Brew (1966). Biochem. Biophys. Res. Corn-mun., 23, 117.
Minemura T. and O. B. Crofford (1969). J.Biol. Chem., 244, 5181.
Sato S, T. Matsushima and T. Sugimura
(1969). Cancer Res., 29, 1437.