cytological and cytochemical studies of centrifuged eggs...
TRANSCRIPT

J. Embryol. exp. Morph. Vol. 27, 1, pp. 1-13, 1972
Printed in Great Britain
Cytological and cytochemical studies of centrifugedeggs of the slug Avion ater rufus L.
By A. H. SATHANANTHAN1
From the Department of Zoology, University of Ceylon, Colombo
SUMMARYThe stratification of various cell organelles and of important chemical substances have
been studied in the eggs of the slug, after moderate centrifugation.As in other molluscs the egg contents stratify typically into three well-defined zones - lipid,
L hyaline and yolky zones - but a distinct equatorial band of inclusions consisting mainly ofF phagosomes and associated lysosomes was detected in the most centrifugal region of the
hyaline zone.The sub-stratification of various cell inclusions in their respective zones was determined
in some detail.The role of the cell membrane and egg cortex in the redistribution of these inclusions and
the nature of the ergastoplasm are discussed in the light of electron-microscope studies ofeggs of this slug and of the sea urchin.
\1 INTRODUCTION
i . The centrifuge has been an extremely valuable tool in cytological and cyto-fc chemical research, especially on eggs. Unlike differential centrifugation and
fractionation procedures used to isolate cellular components in artificial media,by this method the cell inclusions could be concentrated within their cellmembrane and studied in their natural environment.
Such studies have been made of eggs of various other molluscs, includingLimnaea (see Raven, 1958). Recently, centrifuged eggs of the sea urchin havebeen examined with an electron microscope (Mercer & Wolpert, 1962). Themain aim of this paper is to determine in some detail the stratification of variouscell organelles and of certain important chemical substances in a molluscan egg.
MATERIALS AND METHODS
Entire egg capsules with ova, 2-cell and 4-cell stages, were subjected to acentrifugal force of 950 g (at 3750 rev/min) for 6 min using a Cambridge electriccentrifuge. The embryos were then quickly decapsulated in tap-water, rinsed inisotonic saline (0-28 % NaCl) and examined by vital methods or fixed, sectionedand studied by cytological and cytochemical methods.
1 Author's address: 213, N. Roselake Avenue, Los Angeles, California 90026, U.S.A.I E M B 27
A. H. SATHANANTHAN
Table 1. Summary of important staining and cytochemical reactionsof centrifuged embryos
Stain or test
Sudan black B
Acid fuchsin
Mann-Kopsch
PAS
Best's carmine
Hale's reagent
Aldehydefuchsin/Alcianblue
Vitamin C
RNA
FeulgenIron
Calcium
Melanin
Cytochromeoxidase
Succinicdehydrogenase
Reference
McManus'smethod (Pearse,1961)
Metzner & Krause(1928)
McClung Jones(1950)
McManus'smethod (Pearse,1961)
Pearse (1961)
Pearse (1961)
Spicer & Meyer(1960)
Bourne's & Bac-chus's methods(Pearse, 1961)
Kurnick's method(Pearse, 1961)
Pearse (1961)Pearl's method(Pearse, 1961)
Sodium rhodizonate(McGee-Russel,1958)
Lillie's method(Pearse, 1961)
Burstone's method(Pearse, 1961)
Nachlas's method(Pearse, 1961)
Zone
LZHZ
YZ
HZ
YZ
YZ
HZ
YZ
HZ
YZ
HZYZ
HZYZ
YZ
HZ
YZHZYZ
YZ
HZYZ
HZHZYZHZYZ
Stratification Reaction
Lipid globules + + + +Lipochondria + + + +Mitochondria, lysosomes + + +Phagosomes, chromo- + +somes, nucleoli
Golgi + + +Yolk (E, I) + + +, +Mitochondria + + + +Chromosomes, nucleoli + + + +Golgi + + + +Few mitochondria + + + +Golgi + + + +
Lysosomes, phagosomes + + + +Free glycogen + + +Golgi + + + +Yolk (E, I) + + + , + +Free glycogen + + +Phagosomes + +Golgi + + + +Yolk (E, I) + + + , + +Lysosomes, phagosomes + + +Golgi + + + +Yolk (E, I) + + + , + +Lysosomes, phagosomes + + + PGolgi + + + +PYolk (E, I) + + + P , + BFree granules + + + +
Ribosomal RNA, + + +nucleoli
Yolk (E, I) + + , +Chromosomes + + +Free granules, Golgi + + +
Golgi + + + +Yolk (E, I) + + + + , + + +Nucleoli + +Golgi + + + +Yolk (E, I) + + + , + +Nucleoli + +Mitochondria + + + +Golgi + + + +Mitochondria + + +Golgi + + +
<

Centrifuged slug eggs
Stain or test
Acid phos-phatase
Benzidineperoxidase
Thiamine pyro-phosphatase,alkalinephosphatase
Toluidine blue,methylene blue
Neutral red,Nile blue
+ + + + =reaction; +
Table 1
Reference
Azo dye method(Pearse, 1961)
Van Duijn'smethod (Pearse,1961)
Novikoff & Gold-fischer(1961)
Fredricsson'smethod (Pearse,1961)
Baker (1958)
Baker (1958)
Zone
HZ
YZHZ
YZ
HZ
HZYZ
HZ
YZ
Very intense reaction; + += weak reaction; P = purple;
HZ = hyaline zone; LZ = lipid zone; YZ
(cont.)
Stratification
LysosomesPhagosomesGolgiLysosomesPhagosomes
GolgiYolk (E, I)Nucleoli
Lysosomes, phagosomesGolgiYolk (E, I)LysosomesPhagosomesGolgiYolk (E, I)
+ = strong reaction; + +B = blue; E = externum; I
\ = yolky zone.
Reaction
+ + + ++ +
+ + + ++ + + +
+ + +
+ + + ++ + + +, + +
+ +
+ + +P+ + + +B
+ +, +B+ + +
+ ++ + +
+ +, += moderate= internum;
Vital studies included examination of embryos by phase-contrast microscopyand after vital staining and incubating them alive for certain enzymes - ' vitalenzymology' (Sathananthan, 1968). For phase-contrast work the embryos weresquashed in isotonic saline and examined under oil immersion with a Wild M 20phase-microscope. Vital staining was carried out according to Baker (1958).Embryos were stained in Grubler's neutral red, Nile blue, toluidine blue andmethylene blue for 5-40 min using a 0-01 % solution of the dye in isotonicsaline, rinsed, squashed and examined with an apochromatic oil-immersionobjective. Most of the enzyme tests were carried out successfully by incubatingalive whole stratified embryos in various media and examining them in muchthe same way as in the vital staining procedure. Cytochrome oxidase, succinicdehydrogenase, acid phosphatase, benzidine peroxidase and thiamine pyro-phosphatase were the enzymes studied in this manner (Table 1).
Centrifuged embryos were also fixed in a wide variety of fixatives, includingAltmann, Champy, Mann, Helly, Zenker, Ciaccio, Perenyi, Bouin, Carnoy andacetone/ethanol, impregnated with celloidin and rapidly embedded in ceresin/wax (56 °C) according to Peterfi's method (Pantin, 1960). Serial sections (4 /*mthick) of each embryo were cut with a Cambridge rocking microtome and thedistribution of mitochondria, lysosomes, Golgi, yolk and important cyto-chemical substances such as lipids, nucleic acids, polysaccharides, muco-substances, vitamin C, iron, calcium, pigment, thiamine pyrophosphatase and
A. H. SATHANANTHAN
LZ
HZ
EB
YZ
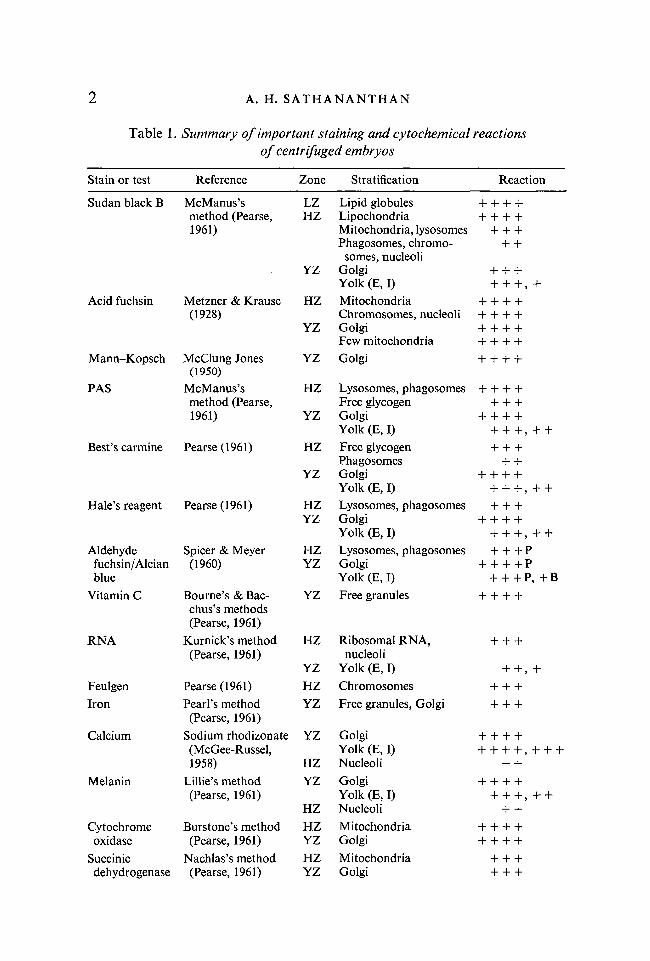
EBERGHZLLGLZM
ABBREVIATIONSequatorial bandendoplasmic reticulumGolgihyaline zonelysosomelipid globulelipid zonemitochondrion
ON FIGURESmNnPPBRYYZ
microsomenucleusnucleolusphagosomepolar bodyribosomeyolkyolky zone
Figs. 1-5. Illustrations showing distributions of various cell inclusions in centrifugedova.Fig. 1. General distribution of cell organelles in lipid, hyaline and yolky zones.Fig. 2. Cytochrome oxidase- and succinic dehydrogenase-positive granules (mito-chondria and Golgi).Fig. 3. Acid phosphatase-positive granules (lysosomes, phagosomes and Golgi).Fig. 4. Benzidine peroxidase activity (pronuclei are copulating).Fig. 5. Glycogen (fine stippling) and vitamin C (coarse granules).

Centrifuged slug eggs 5
alkaline phosphatase were determined using appropriate methods (Table 1).Some embryos were fixed in formaldehyde/saline (10% neutral formalin in0-28 % NaCl), embedded in 15 % gelatin (Pearse, 1961) and frozen sections (6 /tmthick) were cut in a cryostat. These were used to study lipids and acid phos-phatase activity. All colour reactions were visualized with a Zeiss apochromaticobjective and photomicrographs were taken with a Zeiss photomicroscopeusing Ilford Pan-F film.
OBSERVATIONS
Fairly good stratification of various cell inclusions and very sharp demarca-tions between three well-defined zones (fat cap, hyaline and yolky zones) wereobtained immediately after centrifugation (Figs. 1, 6). Broadly speaking therelative amounts of lipid andprotein present in an inclusion seems to determineits stratification. Centrifuged eggs are oval but on standing they round up in afew minutes and redispersion of certain inclusions begins almost immediately,especially between the lipid and hyaline zones. The demarcation between thehyaline and yolky zones, however, persists for about 1 h or more. Prolongedcentrifugation tends to constrict the embryo at the equator and sever it into twosegments. If centrifuged eggs are left to develop in saline or within the capsulecleavage continues apparently normally though slightly retarded. The cleavagefurrows, when established, prevent further redistribution of visible inclusions.
As in other molluscs (Raven, 1958) the egg contents stratify typically into threedistinct major zones (Fig. 1) according to their specific gravity: (a) centripetaloil cap zone, which is lightest and consists of lipid globules; (b) intermediateclear hyaline zone with most of the ergastoplasm, mitochondria, lysosomes,phagosomes, nuclear inclusions and asters; (c) centrifugal yolky zone, which isheaviest and consists mostly of yolk spherules and Golgi bodies. The mostremarkable feature in this embryo is that an equatorial band of inclusions con-sisting mainly of albumen vesicles (phagosomes) and lysosomes was detected inthe most centrifugal region of the hyaline zone.
The axis of stratification is usually more or less parallel or is oblique to theanimal-vegetal axis of the embryo, and the polar bodies are usually attached toits lighter half. Exceptionally they may be found attached to the yolky half.
Substratification of cell organelles
The distribution of various cell organelles in their respective zones will nowbs dealt with. Most of the important cell constituents stratify in the hyalinezone (Fig. 1).
Ergastoplasm is the chief component of this zone. It consists of groundcytoplasm (hyaloplasm), ribosomes and elements of the endoplasmic reticulum(ER). These observations are supported by cytochemical evidence and alsoultrastructural studies of normal eggs (Sathananthan, 1966). The presence of
A. H. S A T H A N A N T H A N
50 //m
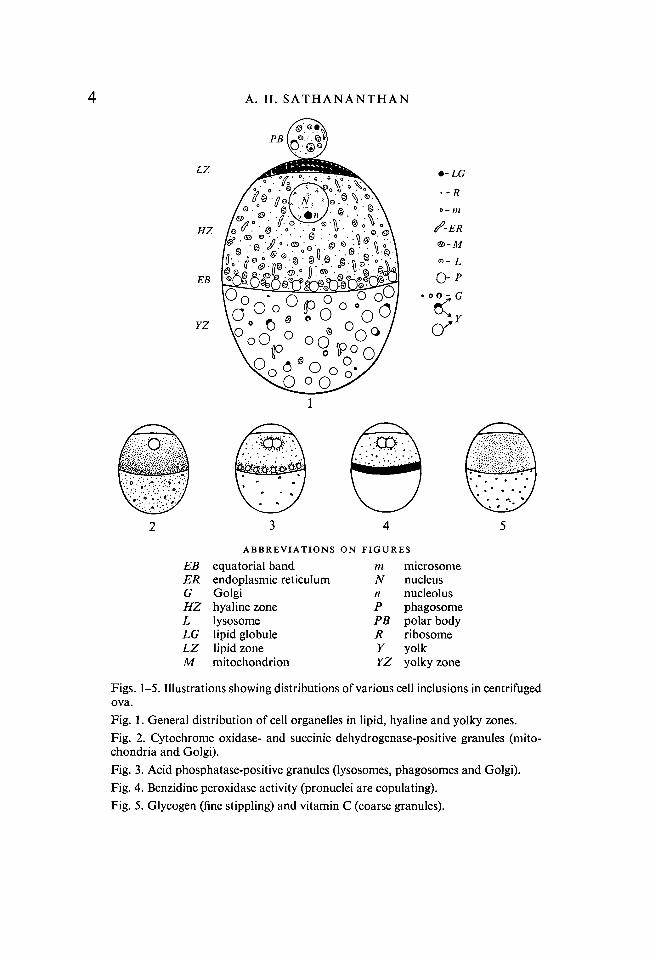
F I G U R E S 6-8
Photomicrographs of centrifuged embryosFig. 6. v.s. mature ovum (Helly/acid fuchsin).Fig. 7. v.s. ovum at second maturation (Helly/Sudan black B). Note equatorial bandand flecks of lipid in yolky zone.Fig. 8. v.s. ovum at second maturation (Helly/PAS). Note equatorial band.

Centrifuged slug eggs 7
ribosomes is deduced by the stratification RNA in this zone (Fig. 12). Electronmicrographs of unstratified eggs show that there is an abundance of free ribo-somes in ova and that the ER is sparsely developed and consists of vesicularelements (microsomes) and a few elongate cisternae, sometimes associated withmitochondria and yolk spherules. Further, strands of ER were also seen afterstaining with pyronin and Sudan black and some of these may remain in theyolky zone on account of their association with yolk spherules.
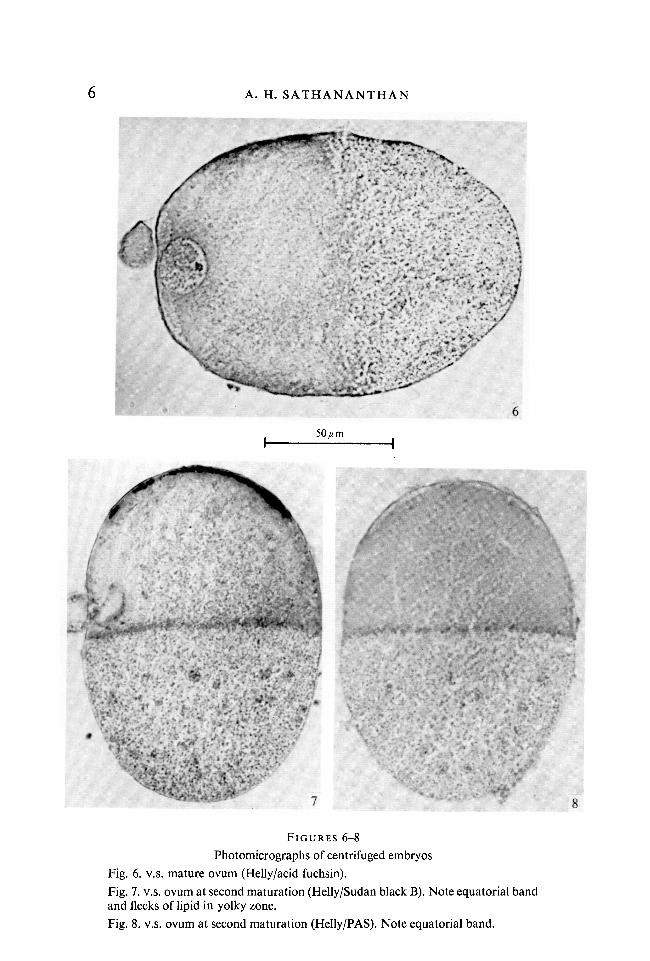
Mitochondria are easily stainable with acid fuchsin and show cytochromeoxidase and succinic dehydrogenase activities. They stratify chiefly in the hyalinezone, forming a very broad band above the equator (Fig. 10). They are moreabundant towards the equator and decrease gradually in number towards theoil cap zone. There is invariably an almost clear region just below the fat capwith few visible inclusions, which presumably abounds with ribosomes andperhaps microsomes. A few mitochondria are always seen in the yolky zoneevenly distributed between the yolk spherules or form rows or chains aroundthe latter.
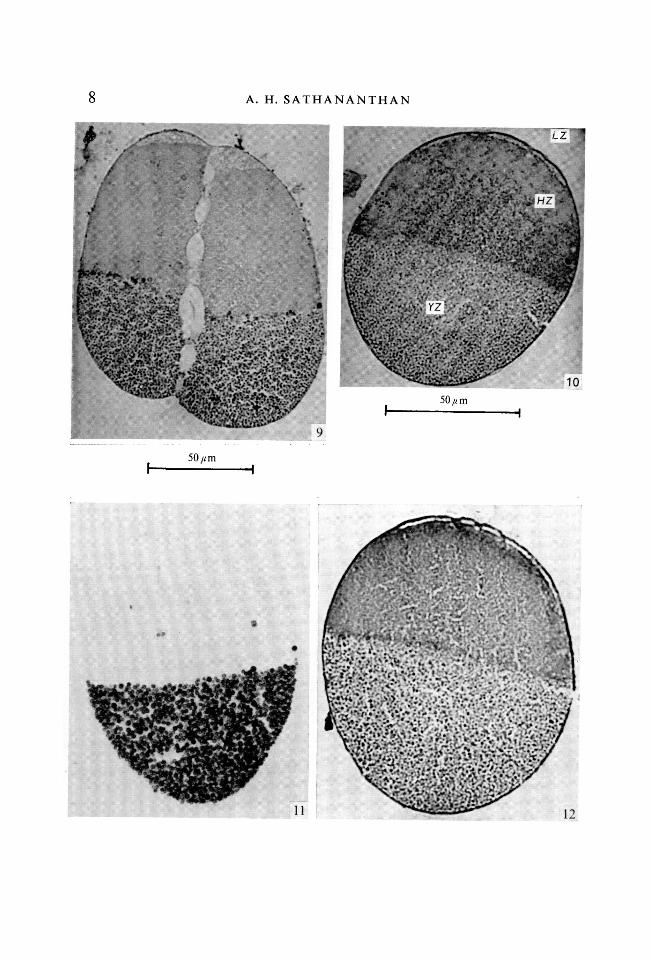
Lysosomes stratify among the mitochondria in the hyaline zone and could bebest demonstrated by the acid-phosphatase test and the periodic acid/Schiff's(PAS) reaction. They also show vital metachromasy with toluidine blue and areneutral red-positive and contain peroxidase. They are, however, more abundanttowards the equator and together with larger bodies called phagosomes, withwhich they are associated as 'satellites', they form a distinct supra-equatorialband (Figs. 1, 7-9). The phagosomes are, in fact, tiny albumen vesicles whichstratify above the equator in the most centrifugal region of the hyaline zone.Albumen is ingested at the egg surface from the very onset of development(Sathananthan, 1968). Perhaps this is the first time that such a layer has beendetected in a molluscan egg.
Nuclear inclusions, etc. Nuclei with nucleoli, maturation and mitotic figuresand associated chromosomes stratify in the hyaline zone. The nuclei are some-times displaced centripetally towards the oil cap (Fig. 6). The second maturationfigure and chromosomes are often seen on a side associated with the first polarbody (Fig. 7) and are not displaced from their normal site at the animal pole.The mitotic figures lie freely in the hyaloplasm and usually occupy a morecentral position in the hyaline zone (Fig. 9).
The heaviest inclusions, chiefly consisting of yolk and Golgi bodies, stratifyin the yolky zone (Figs. 1, 9, 11).
Golgi bodies exist in a variety of forms and range from tiny spheres (dictyoles),duplex vesicles, crescents, bean-shaped bodies to rodlets, as revealed in Mann-Kopsch preparations, where they are heavily impregnated with osmium (Fig. 1).They also show thiamine pyrophosphatase activity and are easily stainable withthe vital dyes used. While the dictyoles and vesicles are independent objects theother dictyosomes are associated with yolk spherules and are inseparable oncentrifugation. The dictyoles and vesicles which have yolky interna usually
i
A. H. SATHANANTHAN
50/tm
10
J

Centrifuged slug eggs 9
stratify in the most centripetal region of this zone just below the equator and afew may occasionally invade the clear hyaline zone.
Yolk spherules could be best demonstrated by tests for thiamine pyrophos-phatase and alkaline phosphatase and by the PAS reaction. They are also stainedby vital dyes but less intensely than Golgi bodies. Each spherule is duplex instructure, with a membranous externum and a yolky internum. They vary insize and are sometimes arranged in their zone along a gradient, the smaller onesbeing more centripetal while the larger mature ones are displaced centrifugally.Yolk is chemically very complex, being glyco-lipo-protein in nature, withproteins predominating.
Stratification of chemical substances
These substances are either found freely or are bound to some cell organelle,in which case their stratification will conform to that of the latter. The cyto-chemical methods used to identify these substances are given in Table 1.
Lipids. Free lipids stratify mainly in the oil cap zone (Figs. 1,7). The oil capstains very intensely with Fettrot and Sudan black B and consists of numerousminute globules and also some diffuse fat. Certain lipid inclusions remain in theyolky zone (Fig. 7) and appear as blobs or flecks in association with a few mito-chondria. All these lipid inclusions are composed of both neutral fats andphospholipids. There are also discrete membrane-bound lipid bodies (lipo-chondria) which stratify close to the oil cap zone or are associated with astersor are found in polar bodies. They are probably overgrown lysosomes impreg-nated with lipid (Sathananthan, 1968). They have a strong affinity for Sudanblack and are stained by vital dyes such as neutral red and Nile blue and henceseem to be predominantly phospholipid. Sudan black also stains the membrane-bound lipids of various cell organelles (Fig. 7). Thus mitochondria, lysosomes,Golgi, yolk membranes, plasma and vitelline membranes, and even strands ofthe ER appear in these preparations. These protein-bound or masked lipids areessentially phospholipid in nature. The phagosomes that stratify at the equator,chromosomes and nucleoli too have appreciable amounts of lipid.
Oxidative enzymes. When centrifuged eggs are incubated for cytochrome
FIGURES 9-12
Photomicrographs of centrifuged embryosFig. 9. v.s. 2-cell stage (Helly/PAS). Note phagosomes and lysosomes at the equator.Fig. 10. v.s. ovum at second maturation showing mitochondria in hyaline zone(Helly/acid fuchsin). Note redispersion of lipid has begun.Fig. 11. v.s. ovum fixed in acetone/ethanol and incubated for thiamine pyrophos-phatase.Fig. 12. v.s. ovum showing distribution of RNA (Helly/pyronin Y). Note diffusestaining in hyaline zone and pyroninophily of yolk granules.

10 A. H. SATHANANTHAN
oxidase and succinic dehydrogenase, the sites of enzyme activity closely corre-spond to the distribution of mitochondria (Fig. 2). The hyaline zone gives anintense positive reaction, the region above the equator being most intense. Oncloser examination, the mitochondria are the chief sites of enzyme activity.Those in the yolky zone and the Golgi bodies there are also intensely positive.The results obtained on incubation for succinic dehydrogenase, however, areless striking than those obtained for cytochrome oxidase (see Sathananthan,1970).
Acid-phosphatase activity. Lysosomes, phagosomes and Golgi bodies arestrongly acid phosphatase-positive (Fig. 3). When incubated by the azo dyemethod the hyaline zone is intensely stained and there appears the supra-equatorial band which consists of phagosomes and associated lysosomes. TheGolgi bodies are found distributed among the yolk spherules, which are acid-phosphatase-negative. Acid-phosphatase-rich granules, chiefly lysosomes anda few lipochondria, are also seen around nuclei and asters in the hyaline zoneand in polar bodies.
Benzidine peroxidase activity. A remarkable reaction is seen when centrifugedeggs are tested for this enzyme (Fig. 4). A sharply defined, intense blue, supra-equatorial band appears in about 20 min of incubation, similar to that observedin Limnaea (Raven, 1958). The picture resembles very much the reaction seen inembryos incubated for acid phosphatase. When examined closely the blue bandis seen to consist of fine irregular granules (lysosomes) and larger rounded bodies(phagosomes). A few of these inclusions are also found in the hyaloplasm,especially around nuclei and asters. A diffuse cytoplasmic reaction in the bandregion is also evident and needle-like spicules are sometimes seen among theinclusions, which is probably an artifact. The identification of benzidine per-oxidase as an intrinsic component of lysosomes and phagosomes, perhaps forthe first time, is rather significant (see Sathananthan, 1968).
Thiamine pyrophosphatase activity is almost exclusively limited to the yolkyzone (Fig. 11). The Golgi bodies and yolk membranes (externa of yolk spherules)are intensely positive while the yolk interna also show some activity. Golgi areprobably involved in vitellogenesis and the yolk membranes are very likelyderived from Golgi membranes (Sathananthan, 1966). Nucleoli in the hyalinezone are also positive.
Alkaline-phosphatase activity. Results are exactly similar to those obtained forthiamine pyrophosphatase activity.
Nucleic acids. Free RNA stratifies in the hyaline zone (Fig. 12) and presumablycorresponds to the distribution of ribosomes in the hyaloplasm. A few strandsof ER that remain in the yolky zone are RNA-positive and these seem to be asso-ciated with yolk spherules. Bound RNA is mainly found in the yolk membranes.The mitochondria in the hyaline zone are also feebly positive while the nuclearRNA is also stainable. The presence of RNA was confirmed with ribonuclease.DNA is found in the nuclei that stratify in the hyaline zone but is rather difficult

Centrifuged slug eggs 11
to demonstrate in ova. The chromosomes, however, are readily DNA-positiveand could be easily stained with acid fuchsin in mitochondrial preparations(Fig. 10).
Polysaccharides and mucosubstances. Glycogen appears finely granular andstratifies in the hyaline zone (Fig. 5). It could be stained with Best's carmine orPAS reagent and removed with saliva. Traces of galactogen could also bedemonstrated in the phagosomes by the same methods, by predigesting withpectinase.
The PAS reaction also reveals larger polymorphic granules (lysosomes) in thehyaline zone and a fairly distinct equatorial band consisting of phagosomes andassociated lysosomes (Figs. 8, 9) corresponding to that seen after incubation foracid phosphatase and benzidine peroxidase. Some of these inclusions are alsoseen, as usual, around nuclei and asters and in polar bodies. In the yolky zone,the yolk membranes and Golgi are also intensely PAS-positive, while the flecksof lipids give a weak reaction. Various mucosubstances and glycolipids areknown to answer the PAS test.
The distribution of acid mucopolysaccharides (AMP) in the hyaline zonealso conforms to that of the phagosomes and lysosomes but the results are notas striking as those obtained with the PAS reaction due to a weaker reactionand a diffuse staining of the hyaloplasm. Aldehyde fuchsin, Alcian blue andHale's reagent were used to demonstrate AMP. More striking results wereobtained using vital metachromasy. Lysosomes and phagosomes stain purplewith toluidine blue, methylene blue and sometimes with Nile blue. The hyalinezone as a whole, however, stains diffuse purple while a more intense meta-chromatic band is seen above the equator corresponding to that seen earlier.The diffuse staining could be due to some free AMP and also RNA that stratifieshere, which is feebly metachromatic. AMP in bound form is also present in theGolgi bodies, yolk membranes (externa), nucleoli, plasma and vitelline mem-branes. All results indicate that most inclusions have a higher content ofsulphated AMP, the exceptions being the yolky interna and vitelline membranewhich have more non-sulphated AMP.
Vitamin C exists freely as sharply defined coarse granules (1-2 /an. in diameter),strongly resembling Golgi bodies, and stratifies in the yolky zone (Fig. 5). Thesubstance, however, is not bound to Golgi or any other inclusion.
Iron. Ferric iron is found distributed diffusely as fine irregular particles (0-3-1 /tm in diameter), which stratify in the yolky zone. It seems to be also boundto Golgi bodies and to a lesser extent to nucleoli.
Calcium is bound to Golgi bodies and yolk spherules that stratify in theyolky zone. The yolk membranes are more intensely positive than their interna.Nucleoli are also calcium-positive. Golgi seem to be involved in the metabolismof calcium.
Pigment. Golgi bodies and yolk spherules have a yellowish-brown pigmentwhich has been identified as melanin. Its distribution is similar to that of calcium.

12 A. H. SATHANANTHAN
The fat cap is also pigmented brown, but this pigment has not been confirmedas melanin.
DISCUSSION
The results show that by moderate centrifugation one could obtain a fairlysharp stratification of cell inclusions. This undoubtedly has been most valuablein subcellular studies of this egg. Although stratification is sharp, it is by nomeans complete. A few mitochondria and elements of the ergastoplasm arealways seen in the yolky zone. Likewise some lipid inclusions are retained inthe yolky half or may invade the hyaloplasm. Exceptionally a few Golgi bodiesor yolk spherules may be found in the hyaline zone. This confirms the view thatcomplete stratification cannot be achieved by the moderate centrifugationcommonly used in embryological studies.
The centrifugal force causes an inevitable stretching of the cell membrane butit is capable of regaining its original form when the force ceases to act. Thisindicates that the membrane is fairly elastic in nature. The tension developedin it probably causes the egg to round up soon after centrifugation. In Aplysiathe elastic recovery of the spherical shape by the elongated egg after centrifuga-tion is thought to play an important part in the redistribution of its cytoplasmiccontents (see Raven, 1958). This seems to be the case in Arion.
It has been also postulated that in certain molluscs the egg cortex may play apart in the redistribution of inclusions (ooplasmic re-segregation) after centrifu-gation, and that it is the seat of polarity of the egg (Raven, 1958). It is believedthat this cortex is a gel that cannot be displaced by moderate centrifugation.This is also the commonly accepted view in the case of the sea urchin. There isno structural evidence of a cortical gel layer in the egg of this slug. Electronmicrographs of normal eggs (Sathananthan, 1966) show a well-defined 'unit'cell membrane and a zone usually devoid of larger granules (about 1 jam thick)below it, corresponding to a clear agranular cortical region seen with the lightmicroscope. This zone has an abundance of ribosomes and also vesicularelements of the ER and those of micropinocytotic origin. Occasionally largerinclusions like mitochondria and lysosomes invade it and are found very closeto the cell membrane, indicating the absence of a cortical gel layer. Further,ultrastructural studies of both normal and centrifuged eggs of the sea urchin(Mercer & Wolpert, 1962) have failed to detect a gel-like cortex, beneath thecell membrane. These authors have, however, shown that the small particulatecytoplasmic fraction (probably ergastoplasm) appears to be relatively unaffectedby centrifugation and forms a continuous 'ground substance' in which arefound the larger mobile inclusions, such as mitochondria and yolk granules.This, indeed, is the impression gained by the present study. The incompletestratification of the ER and undoubtedly of the hyaloplasm which suspends thegranules even in the yolky zone is evidence that the ergastoplasm is relativelyunaffected. A similar electron-microscope investigation of centrifuged eggs,however, needs to be carried out for further confirmation of these views.

Centrifuged slug eggs 13
I am most grateful to Professor Alastair Graham and Dr Vera Fretter for providing thefacilities for my work at the University of Reading, U.K., and for their supervision andencouragement.
REFERENCES
BAKER, J. R. (1958). Principles of Biological Microtechnique. London: Methuen.MCCLUNG JONES, R. (1950). McClung's Handbook of Microscopical Technique. New York:
Hoeber.MCGEE-RUSSEL, S. M. (1958). Histochemical methods for calcium. /. Histochem. Cytochem.
6, 22-42.MERCER, E. H. & WOLPERT, L. (1962). An electron microscope study of the cortex of the sea
urchin (Psammechinus mi liar is) egg. Expl Cell Res. 27, 1-13.METZNER, R. & KRAUSE, R. (1928). In Abderhalden's Handbuch der biologischen Arbeits-
methoden, Abt. v, Teil 2, Halfte i, s. 325.NOVIKOFF, A. B. & GOLDFISCHER, S. (1961). Nucleosidephosphatase activity in the Golgi
apparatus and its usefulness for cytological studies. Proc. natn. Acad. Sci. U.S.A. 47, 802.PANTIN, C. F. A. (1960). Notes on Microscopical Technique for Zoologists. Cambridge Univer-
sity Press.PEARSE, A. G. E. (1961). Histochemistry: Theoretical and Applied. London: Churchill.RAVEN, CHR. P. (1958). Morphogenesis: The Analysis of Molluscan Development. New York:
Pergamon Press.SATHANANTHAN, A. H. (1966). Studies on the Early Embryology of the Slug Arion ater rufus
L. Ph.D. thesis, Reading University, U.K.SATHANANTHAN, A. H. (1968). Lysosomes in the early development of the slug Arion.
Proc. Int. Symp. Mollusca, India 2, 687-706.SATHANANTHAN, A. H. (1970). Studies on mitochondria in the early development of the slug
Arion ater rufus L. / . Embryol. exp. Morph. 24, 555-582.SPICER, J. R. & MEYER, D. B. (1960). Histochemical differentiation of acid mucopoly-
saccharides by means of combined aldehyde fuchsin-Alcian blue staining. Am. J. clin.Path. 33, 453^60.
{Manuscript received 10 February 1971)