investigation into the control of melittin secondary
TRANSCRIPT

Purdue UniversityPurdue e-Pubs
Open Access Theses Theses and Dissertations
Summer 2014
Investigation into the control of melittin secondarystructure and antimicrobial activityZachary B. MolinetsPurdue University
Follow this and additional works at: https://docs.lib.purdue.edu/open_access_theses
Part of the Analytical Chemistry Commons, Biochemistry Commons, Medicine and HealthSciences Commons, and the Pharmacology Commons
This document has been made available through Purdue e-Pubs, a service of the Purdue University Libraries. Please contact [email protected] foradditional information.
Recommended CitationMolinets, Zachary B., "Investigation into the control of melittin secondary structure and antimicrobial activity" (2014). Open AccessTheses. 656.https://docs.lib.purdue.edu/open_access_theses/656

Graduate School ETD Form 9 (Revised 12/07)
PURDUE UNIVERSITY GRADUATE SCHOOL
Thesis/Dissertation Acceptance
This is to certify that the thesis/dissertation prepared
By
Entitled
For the degree of
Is approved by the final examining committee:
Chair
To the best of my knowledge and as understood by the student in the Research Integrity and Copyright Disclaimer (Graduate School Form 20), this thesis/dissertation adheres to the provisions of Purdue University’s “Policy on Integrity in Research” and the use of copyrighted material.
Approved by Major Professor(s): ____________________________________
____________________________________
Approved by: Head of the Graduate Program Date
Zachary B. Molinets
INVESTIGATION INTO THE CONTROL OF MELITTIN SECONDARYSTRUCTURE AND ANTIMICROBIAL ACTIVITY
Master of Science
Jean A. Chmielewski
Arun K. Ghosh
Christine A. Hrycyna
Jean A. Chmielewski
R. E. Wild 7/17/2014


INVESTIGATION INTO THE CONTROL OF MELITTIN SECONDARY
STRUCTURE AND ANTIMICROBIAL ACTIVITY
A Thesis
Submitted to the Faculty
of
Purdue University
by
Zachary B. Molinets
In Partial Fulfillment of the
Requirements of the Degree
of
Master of Science
August 2014
Purdue University
West Lafayette, Indiana

ii
For my wife, Natalie, who has helped me grow and open my mind,
since the first time we said hello.

iii
TABLE OF CONTENTS
Page
LIST OF TABLES ...............................................................................................................v
LIST OF FIGURES ........................................................................................................... vi
LIST OF SCHEMES........................................................................................................ viii
ABSTRACT ....................................................................................................................... ix
CHAPTER ONE ANTIMICROBIAL PEPTIDES ..............................................................1
Introduction ....................................................................................................................1 Antimicrobial Peptides...................................................................................................2
Sources of Antimicrobial Peptides...........................................................................4 Antimicrobial Peptides with α-helical Secondary Structure ..........................................6
Mechanisms of Action .............................................................................................6 Metal Stabilized/Destabilized Peptides..........................................................................9 Siderophores ................................................................................................................11 References ....................................................................................................................13
CHAPTER TWO MELITTIN ANALOGUES ..................................................................16
Melittin .........................................................................................................................16 Activity of Melittin ................................................................................................18 Structural Requirements for Activity .....................................................................19
Truncated Melittin .......................................................................................................20 1st Generation Melittin Derivatives ..............................................................................21
Synthesis of Metal Binding Moieties ...............................................................23 Synthesis of Melittin Derivatives.....................................................................24 Circular Dichroism...........................................................................................25 Antimicrobial Activity .....................................................................................27
2nd Generation Melittin Derivatives .............................................................................27 Synthesis of 2nd Generation Melittin Derivatives ...........................................28 UV/Vis Spectroscopy.......................................................................................29 Circular Dichroism...........................................................................................30 Antimicrobial Activity .....................................................................................33

iv
Page
Reversibility of the Metal-Ligand Interaction .............................................................34 Melittin Derivatives with Other Metals .......................................................................35 Conclusions ..................................................................................................................37 Materials and Methods .................................................................................................37 Experimental ................................................................................................................38 References ....................................................................................................................43

v
LIST OF TABLES
Table Page
1.1 Amino acid sequences of cecropin-A, magainins-II, protegrin-1, melittin and defensins. Subscript numbering indicates disulfide bridge ............................................4
1.2 Examples of antimicrobial peptides isolated from invertebrates. ..................................5
2.1 Purification of melittin derivatives ..............................................................................25
2.2 Purification of truncated melittin derivatives ..............................................................29
2.3 MIC of melittin derivatives ..........................................................................................34

vi
LIST OF FIGURES
Figure Page
1.1 The barrel-stave representation of AMP activity ...........................................................7
1.2 Toroidal Pore representation of AMP activity ...............................................................8
1.3 The “carpet model” representation of AMP activity .....................................................9
1.4 Generic profile of an α-helix highlighting positions of interaction. ............................10
1.5 Structures of enterobactin(top), pyochelin(bottom left), and rhodotorulic acid(bottom right) ........................................................................................................12
2.1 The structure of melittin...............................................................................................16
2.2 Helical wheel diagram of melittin ................................................................................17
2.3 3D representation of melittin showing the “bent rod” .................................................18
2.4 The structure of truncated melittin(12-25). ..................................................................21
2.5 Placement of metal binding moieties to melittin, where X represents bipyridine or terpyridine ..............................................................................................21
2.6 Metal assisted deactivation of peptide with siderophore reactivation .........................22
2.7 CD spectra of melittin derivatives, 50 μM, in 100 mM phospate buffer, pH 7. ..........26
2.8 CD spectra of melittin derivatives, 50 μM, in 100 mM phospate buffer, pH 7.4, with 1 equivalent Fe(II) ........................................................................................26
2.9 E. coli antibacterial assay of melittin derivatives. 6 hour incubation ..........................27
2.10 Second generation melittin derivatives. From top to bottom: MLTx, MLTx1, MLTx2, MLTx3. Where X is bipyridine or terpyridine ..............................................28
2.11 UV/Vis spectroscopy of MLTx1-tpy:Fe(II), 100 μM, in water .................................30

vii
Figure Page
2.12 UV/Vis spectroscopy of MLTx3-tpy:Fe(II), 100 μM, in water .................................30
2.13 CD of melittin derivatives, 50 μM, in 10 mM phosphate buffer, pH 7.4 ..................32
2.14 CD of melittin derivatives, 50 μM, in 10% HFIP/water ............................................32
2.15 CD of melittin derivatives, 50 μM, with metal ion in 10 mM phosphate buffer, pH 7.4. MLTx3d-tpy was denatured prior to metal addition ...........................33
2.16 MLTx3-tpy with 1 equiv Fe(II) in solution. 500 µM solution of the peptide-Fe(II) complex with 2 equivalents of chelators ...........................................................35
2.17 UV/Vis spectrum of MLTx3-tpy:Co(II), 100μM, (blue) and terpyridine control (red) .................................................................................................................36
2.18 CD of melittin derivatives, 50 μM, in 10 mM phosphate buffer, pH 7.4 ..................36

viii
LIST OF SCHEMES
Scheme Page
2.1 Synthesis of metal binding ligands ..............................................................................23
2.2 Synthesis of melittin derivatives, where X is terpyridine or bipyridine ......................24

ix
ABSTRACT
Molinets, Zachary B. M. S., Purdue University, August 2014. Investigation Into the Control of Melittin Secondary Structure and Antimicrobial Activity. Major Professor: Jean A. Chmielewski.
Antimicrobial resistance has been an exponentially growing problem since the
discovery of antibiotics. Antibiotics have been misused for many years and this misuse
has grown into a real problem for the medical community. While there are countless
safeguards to prevent infection by a resistant strain of bacteria, there are still many
plagued by it and must be treated with sometimes dangerous antibiotics. Melittin, along
with many other peptides, contain potent antimicrobial properties, but are also toxic
toward enthrocytes. The control of the secondary structure of peptides provides the key to
adjusting their activity.

1
CHAPTER ONE ANTIMICROBIAL POLYPEPTIDES
Introduction
Antimicrobial resistance has been an exponentially growing problem since the
discovery of antibiotics. The misuse of antibiotics is a major driving force for antibiotic
resistance. The first source of misuse was during the golden age of antibiotics, when there
was very little resistance. Antibiotics would be prescribed for the slightest symptoms,
with this wide spread use bacteria found ways to adapt and resist the drugs.1 Another
source of misuse is in medical patients, when a prescription is not taken as directed.
Patients across the globe, especially in less developed countries, have said that they save
or do not finish the prescription.2 Overuse in agriculture is another major source of
resistance. In 2010 the FDA released a report estimating that the use of antibiotics in food
animals was over 11 million kilograms were used in 2009, and 2011 the number has rose
to 13.5 million kilograms.3 After bacteria encounter a class of antibiotics over several
generations, the antibiotic becomes less effective and resistance is born.
Resistance is formed by mutations in the bacterial DNA that evolve mechanisms
to counter antibiotics. β-lactams are a class of antibiotics which block a pathway in the
production of the cell wall, thereby killing the bacteria. β-lactams such as penicillin or
amoxicillin are the workhorse of antibiotics and are the most prescribed antibiotics
available. As such, the resistance to β-lactams has risen dramatically. The mechanism of

2
bacterial resistance has been pinpointed to β-lactamase enzymes. These enzymes cleave
the lactam ring making the β-lactam compounds inert. There are several groups of β-
lactamase enzymes, each are capable of a range of mechanisms. For example, some β-
lactamase enzymes are capable of being inhibited by clavulanic acid, a β-lactam mimetic.
However, there are also some that show resistance to clavulanic acid. These data point to
the necessity for novel strategies to combat resistance.
There is not only a biological cost to the increase in bacterial resistance but a
societal cost as well. Increasing resistance brings with it increased mortality and health
care expenses. For salmonella, a fairly common source for food poisoning, the mortality
rate against resistant strains is 3.4%, while only 0.2% for non-resistant strains.4 The
increased patient cost to treat resist strains of bacteria in the United States is greater than
8 billion US dollars.5
Antimicrobial Peptides
Antimicrobial resistance from misuse of antibiotics has created a global health
concern which draws the need for novel types of antimicrobial therapeutics.
Antimicrobial peptides are short, 15-40 amino acids, peptides that are a part if the innate
immune response in organisms.6 Many isolated AMPs have been shown to have activity
against a wide array of organisms including Gram-positive and Gram-negative bacteria,
fungi, and viruses.7 The diverse mechanisms of activity or AMPs include formation of
pores in the membrane, inhibiting proteins, and triggering autolysis.6 Antimicrobial

3
peptides are attractive as a new class of clinical antibiotics because of their broad
spectrum of activity and diverse mechanisms in contrast to small molecule antibiotics.8
Antimicrobial peptides (AMPs) first came into light in the 1980’s when Boman,
Zasloff, and Lehrer concurrently isolated AMPs from separate animal kingdoms.9,10
Notably, in Zasloff’s lab, it was noticed that infections rarely occurred after non-sterile
surgical procedures with the African clawed frog Xenopus laevis.11 It was discovered that
small peptides in the skin had broad spectrum antibacterial properties, and these peptides
became known as magainins.11 Since the discovery of the first AMPs over 800, have been
found throughout the animal and plant kingdom.12
There are several inherent characteristics associated with AMPs. AMPs are
usually short, less than 50 amino acids, and contain a cationic amphipathic structure.13
AMPs are classified by their amino acids and secondary structure: α-helix, β-sheet,
cysteine containing, containing many standard AA, and containing modified AAs.7
Cecropins, one of the first classes identified, is a cationic amphipathic peptide isolated
from Hyalophora cecropia.14 Through NMR studies it was determined that cecropins
form an α-helix in 15% HFIP.15 Peptides which form α-helical secondary structures are
most known for their pore forming mechanism. Some well studied peptides with this
mechanism are cecropins16, melittin17 magainins18, and protegrin-1.19

4
Table 1.1: Amino acid sequences of cecropin-A, magainins-II, protegrin-1, melittin and defensins. Subscript numbering indicates disulfide bridge.
Peptide Sequence
Cecropin-A KWKLFKKIEKVGQNIRDGIIKAGPAVAVVGQATQIAK-NH2
Magainin-II GIGKFLHSAKKFGKAFVGEIMNS
Protegrin-1 RGGRLC1YC2RRRFC2VC1VGR-NH2
Melittin GIGAVLKVLTTGLPALISWIKRKRQQ-NH2
Human α-
defensin ACYCRIPACIAGERRYGTCIYQGRLWAFCC
Sources of Antimicrobial Peptides
As stated earlier, the first AMPs isolated were from amphibians and many more
AMPs have since been isolated from a plethora of species, including insects, plants,
animals, and humans. Additionally, these peptides have a broad range of secondary
structures and activity against different types of invasive organisms.20 This menagerie of
structure, activity, and chemical properties is a hotspot for the development of novel
therapeutics.
AMPs in invertebrates were first isolated from Hyalophora cecropia, commonly
known as the Cecropia moth, the largest moth in North America.21 These peptides
became known as ceropins and have broad spectrum activity against protozoa, bacteria,
and fungi.7 Since the initial discovery of AMPs of the Cecropia moth several unique
AMPs have been identified in many other invertebrates ranging from honey bees to crabs.

5
A sample of some of those peptides can be seen in table 1.2 with their respective
activities.
Table 1.2: Examples of antimicrobial peptides isolated from invertebrates
Source Peptide Activity
Drospohila melanogaster Drosomycin F
Diptericin G-
Aedes Aegypti Defensin-α G-,G+
Defensin-β G-,G+
Apis mellifera Mellitin G-,G+, H
Defensin G-,G+
Royalisin G-,G+
Lycosa carolinensis Lycotoxin G-,G+
Stomoxys calcitrans Smd1 G-,G+
Smd2 G-,G+
Tachypleus tridentatus Tachyplesin G-,G+
Limulus polyphemus Tachyplesin II G-,G+,F
Polyphemnsin I G-,G+,F
Polyphemnsin II G-,G+,F
Androctonus australis Androctonin G-,G+,F
Mytilus edulis Mytilin A G-,G+
Hyalophora cecropia Cecropin G-
G-, Gram Negitive; G+, Gram Positive; F, antifungal; H, Hemolytic
With the initial discovery of AMPs in amphibians and soon invertebrates, it
wasn’t long until AMPs were being discovered in many other species including plants
and other vertebrates. Defensins are a group of AMPs that are found in several different
mammals including humans and rabbits, and they also have been isolated from plants and
insects.22 Defensins are 41 amino acids in length and are characterized by their disulfide

6
bridges. Defensins are classified into sub-categories depending on the arrangement of the
disulfide bridges.
Antimicrobial Peptides with α-helix Secondary Structure
Particular classes of AMPs are highly predisposed to form pores in plasma
membranes. α-helical AMPs are one such class that lyses pathogens and erythrocytes. An
α-helical AMP's first interaction with a target is electrostatic due to the cationic amino
acids (AAs) interacting with the negatively charged heads of the phospholipids in the
plasma membrane or other negatively charged components on the cell wall. There is
much speculation about the mechanism of pore forming peptides, but the consensus is
that three types of mechanisms exist: toroidal pore, carpet model, and barrel stave.23
Mechanism of Action
The barrel stave model, figure 1.1, describes a mechanism where the peptides
integrate into the plasma membrane and form a pore. The amphiphatic characteristics of
the peptide make this possible in that the hydrophobic features of the peptide is faced
outward toward the hydrophobic tails of the membrane and the hydrophilic face forms an
aqueous pore.24 One peptide alone cannot form the aqueous pore, therefore, many
peptides come together to form the pores. Some AMPs, such as melittin, utilize a
structural feature known as the leucine zipper to hold the pore together.25 The stability of
the hydrophobic interactions and the leucine zipper allow for the intercellular material to
leak through the pore from the cell.

m
th
H
hy
p
w
co
Figure
The se
mechanism. T
he peptides i
However, onl
ydrophobic
lasma memb
with the pepti
ontributes al
e 1.1: The ba
econd mode
The toroidal
interact perp
ly the phosph
tails.26 As sh
brane curves
ides. Unlike
long with the
arrel-stave re
l of AMP in
pore mecha
endicularly t
holipid head
hown in figu
s in the toroid
the barrel st
e peptides to
epresentation
nduced killin
anism is very
to the plane
d groups inte
ure 1.2, the c
dal pore and
tave model,
o form the po
n of AMP ac
ng of pathoge
y similar to th
of the plasm
eract with the
core differen
d the polar ph
in this mode
ore.27
ctivity.12
ens is the tor
the barrel sta
ma membran
e peptide, no
nce between
hospholipid
el, the plasm
roidal pore
ave model in
ne to form po
ot the
the models i
heads intera
ma membrane
7
n that
ores.
is the
act
e

fu
th
m
ca
fo
fo
as
sh
Figur
The fi
undamentally
he plane of th
mechanism ex
ationic amin
orming a por
orm holes in
s a detergent
hows how th
re 1.2: The T
inal model fo
y different fr
he membran
xplains the a
no acids simi
re, they aggr
n the membra
t, destabilizi
he peptides a
Toroidal Pore
or AMP acti
from the prev
ne instead of
activity of ov
ilarly interac
regate on the
ane which al
ing the plasm
aggregate on
e representat
ivity is the ca
vious model
f forming per
vispirin, an A
ct with the an
e surface. At
llows more p
ma membran
n the extracel
tion of AMP
arpet model
s in that the
rpendicular i
AMP found
nionic plasm
t a critical co
peptides to a
ne and formin
llular side of
P activity.12
. This mecha
peptides ori
interactions.
in myeloid c
ma membran
oncentration
aggregate. T
ng micelles.
f the plasma
anism is
ient parallel
. This
cells.28 The
e, but instea
, the peptide
The peptides
Figure 1.3,
a membrane
8
to
ad of
es
act
and,

co
th
F
ir
fi
im
co
onsequently
he rest.29 Wi
igure 1.3: Th
It is w
ron or zinc a
ingers”, or o
mportant to t
omplexes ar
, work into t
thout the me
he “carpet m
well known th
s cofactors.
ffer activity,
the survival
re usually his
the intracellu
embrane, the
model” repre
Metal Stabi
hat many pe
These cofac
, as with cyt
of multicellu
stidine or the
ular membra
e necessary i
sentation of
ilized/Destab
eptides and p
ctors can pro
ochrome p4
ular organism
e anionic AA
ane and lyse
intracellular
f AMP activi
bilized Pepti
proteins cont
ovide structur
50. These m
ms. The liga
As, aspartic a
portions of m
material is l
ity.12
ides
tain various m
ral rigidity,
metal to ligan
ands in these
acid and glu
membrane f
lost.
metals such
as with “zin
nd complexe
e bioinorgani
utamic acid. T
9
from
as
c
s are
ic
The

sa
an
m
p
d
th
se
h
th
th
F
cr
ame metal b
nd to stabiliz
Stabil
moieties that
ositions prov
isulfide link
he peptide it
econdary str
elical peptid
he strain that
he helix.
igure 1.4: G
Befor
ross-links w
inding techn
ze or destabi
lizing or dest
will account
vide a stabili
kage or coval
creates a “st
ructure. Conv
de has the ab
t is applied t
Generic profil
e the use of
ere common
nique is used
ilize peptide
tabilizing re
t for the prop
izing effect w
lent linkage.
taple” which
versely, crea
ility to desta
to the second
le of an α-he
metal-ligand
nly used, suc
d to research
secondary s
quires specif
per effect. I
when locked
30 By linkin
h holds the tu
ating a linkag
abilize the he
dary structur
elix highligh
d complexes
ch as disulfid
the formatio
structures.
fic placemen
In figure 1.4
d together by
ng two amino
urns of the h
ge between t
elix.31 This d
re by the link
hting position
s to stabilize
de links or am
on of higher
nt of ligands
, the i, i+ 4 a
y an interact
o acids on th
helix in place
the i, i + 2 p
destabilizing
kage form op
ns of interac
α-helical pe
mide bonds.
r order struct
s or other
and i + 7
tion, such as
he same face
e stabilizing
positions on a
g effect is du
pposite sides
ction.
eptides, cova
. For instanc
10
tures
e of
the
a
ue to
s of
alent
ce,

11
the Shultz laboratory synthesized model peptides and incorporated a disulfide bridge at
the i, i + 7 positions.32 Taylor incorporated amide bonds into model peptides utilizing the
natural functional groups of lysine and aspartic acid or glutamic acid.33 The formation of
the amides stabilized the helical structure when compared to the acyclic analogue. While
these methods could provide triggers, as with the disulfide bonds, or permanent
modifications with amide bonds, more reversible methods could provide great value.
Metal – ligand interactions allow a lot of variability and exploration with various
metals. For example, Paul Hopkins’ laboratory used various divalent metal ions to
stabilize the helicity of model peptides.34 The study found that Co(II), Ni(II), Cu(II),
Zn(II), and Cd(II) all had a positive effect with most of the peptides. In another study by
Futaki et al, the group was able to stabilize or destabilize model peptides using
iminodiacetic acid.34 The group made another step to selectively destabilize the Jun-Fos
herterodimer to form a Jun-Jun homodimer.31 The destabilizing of one peptide in the
heterodimer changes the natural recognition of the dimer.
Siderophores
Iron is a critical metal in many organisms as it is needed to do a range of activities
in different enzymes. In mammals iron is needed to transport oxygen in the blood and is a
cofactor of superoxide dismutase which neutralizes harmful radicals in the body. Iron is
also required by bacteria. For example, in E. Coli it is a cofactor in 3-deoxy-D-
arabinoheptulosonate 7-phosphate (DAHP) synthase which is used to make aromatic
compounds and is in the biosynthetic pathway to produce the aromatic amino acids.35

12
Bacteria obtain iron by releasing a class of compounds known as siderophores. These
compounds have some of the highest known binding constants to iron(III). For example,
enterobactin, is the siderophore released by E.Coli for iron scavenging. Enterobactin
binds to iron with the catechol moieties and is brought into the cell by transport proteins
which have receptors on the cell surface.36 Rhodotorulic acid is another siderophore
isolated form Rhodotorula pilimanae.37 Unlike enterobactin which is a catechol based
siderophore, rhodotorulic acid is a hydroxymate. Additionally, rhodotorulic acid contains
only four binding sites, a complex of Fe2(RA)3 must be formed. Pyochelin is another
siderophore isolated from Pseudomonas aeruginosa.38 Pyochelin is unique in its structure
compared with other siderophore which are usually rich in catechols or hydroxymates.
Figure 1.5: Structures of enterobactin(top), pyochelin(bottom left), and rhodotorulic acid(bottom right).

13
References
1) Levy, S. The Antibiotic Paradox: How the misuse of antibiotics destroys their curative powers. 2nd ed. Perseus, Boston, 2001
2) Pechere, J. Patients’ Interviews and misuse of antibiotics. Clin Infect Dis., 2001, 33, S170-S173
3) Holmberg S, Solomon S, Blake P. Health and economic impacts of antimicrobial resistance. Review of Infectious Diseases 1987, 9, 1065-1078
4) John J, Fishman N. Pragmatic role of the infectious diseases physician in controlling antimicrobial costs in the hospital. Clinical Infectious Diseases 1997, 24, 471-85
5) Medzhitov, R Janeway, C, Innate immunity: The virtues of a nonclonal system of recognition cell. 1997, 91, 295–298
6) Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature, 2002, 415, 389-395
7) Reddy, K.V.R., Yedery, R.D., Aranha, C., Antimicrobial peptides: premises and promises. Int. J. Antimicro. Agents, 2004, 24, 536-554
8) Brandenburg , L, Merres, J , Lea-Jessica, V, Deike, P, Thomas. Antimicrobial peptides: multifunctional drugs for different applications. Polymers, 2012, 4, 539-560
9) Steiner, H., Hultmark, D., Engstrom, A., Bennich, H. & Boman, H. G. Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature, 1981, 292, 246–248
10) Ganz, T., Selsted, M. E. & Lehrer, R. I. Defensins. Eur. J. Haematol, 1990, 44, 1–8
11) Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus skin: isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc. Natl Acad. Sci. USA, 1987, 84, 5449–5453
12) Brogden, K. Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria. Nature, 2005, 3, 238-250
13) Zaiou, M. Multifunctional antimicrobial peptides: thearpeutic targets in several human diseases. J. Mol. Med. 2007, 85, 317-329
14) Moore, A, Beazley, W, Bibby, M, Devine, D. Antimicrobial activity of cecropin. J. Anti. Chem., 1996, 37, 1077-1089

14
15) Holak. T. A., Engstrom, A., Kraulis. P. J ,. Lindeberg. G., Bennich. H., Jones, T. A., Gronenborn. A. M. & Clore. G. M. The solution conformation of the anti bacterial peptide cecropin A: a nuclear magnetic resonance and dynamical simulated annealing study. Biochemistry, 1988, 27, 7620-7629
16) Shai, Y. Molecular recognition between membrane-spanning polypeptides. Trends Biochem. Sci., 1995, 20, 460–464
17) Yang, L., Harroun, T. A., Weiss, T. M., Ding, L. & Huang, H.W. Barrel-stave model or toroidal model? A case study on melittin pores. Biophys. J., 2001 81, 1475–1485
18) Yang, L., Harroun, T. A., Heller, W. T., Weiss, T. M. & Huang, H. W. Neutron off-plane scattering of aligned membranes. Method of measurement. Biophys. J., 1998, 75, 641–645
19) Bechinger, B. The structure, dynamics and orientation of antimicrobial peptides in membranes by multidimensional solid-state NMR spectroscopy. Biochim. Biophys. Acta, 1999, 1462, 157–183
20) Reddy, K.V.R., Yedery, R.D., Antimicrobial peptides as microbicidal contraceptives: prophecies for prophylactics – a mini review. Euro. J. Cont. Rep. Health Care, 2005, 10, 32-42
21) Steiner, H., Hultmark, D., Engstrom, A., Bennich, H., Boman, H. G., Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature, 1981, 292, 246-248
22) García-Olmedo, F., Molina, A., Alamillo, J. M. and Rodríguez-Palenzuéla, P. Plant defense peptides. Biopolymers, 1998, 47, 479–491
23) Yeaman , M, Yount, N., Mechanisms of antimicrobial peptide action and resistance. Pharm. Rev. 2003, 51, 27-55
24) Matsuzaki K, Sugislita K, Fshibe N, et al. Relationship of membrane curvature to the formation of pores by magainin 2. Biochemistry 1998, 37, 11856–11863
25) Asthana, N., Yadav, S., Ghosh, J., Dissection of antibacterial and toxic activity of melittin. J. Bio. Chem., 279, 55042-55050
26) Yang, L., Harroun, T., Weiss, T., Ding, L., Huang, H. Barrel-stave model or toroidal model? A case study on melittin pores. Biophysical Journal, 2001, 81, 1475–1485
27) Matsuzaki, K., Murase, O., Fujii, N, Miyajima, K., An antimicrobial peptide, magainin 2, induced rapid flip-flop of phospholipids coupled with pore formation and peptide translocation. Biochemistry, 1996, 35, 11361-11368

15
28) Yamaguchi, S., Huster, D., Waring, A., Lehrer, R., Kearney, W., Tack, B., Hong, M., orientation and dynamics of an antimicrobial peptide in the lipid bilayer by solid-state nmr spectroscopy. Biophysical Journal, 2001, 81, 2203–2214
29) Shai, Y., Mechanism of the binding, insertion and destabilization of phospholipid bilayer membranes by K-helical antimicrobial and cell non-selective membrane-lytic peptides. Biochimica et Biophysica Acta, 1999, 1462, 55-70
30) Henchey, L., Jochim, A., Arora, P., Contemporary strategies for the stabilization of peptides in the a-helical conformation Current Opinion in Chemical Biology 2008, 12, 692–697
31) Futaki, S., Kiwada, T., Sugiura, Y., control of peptide structure and recognition by Fe(III)-induced helix destabilization. J. Am. Chem. Soc. 2004, 126, 15762-15769
32) Jackson, D. Y.; King, D. S.; Chmielewski, J.; Singh, S.; Schultz, P. G. General approach to the synthesis of short α-helical peptides. J. Am. Chem. Soc. 1991, 113, 9391-9392
33) Osapay, G.; Taylor, J. W. Synthesis and conformational properties of a highly α-helical uncosapeptide constrained by three side-chain to side chain lactam bridges. J. Am. Chem. Soc. 1992, 114, 6966-6973
34) Ruan, F.; Chen, Y.; Hopkins, P. B. Metal ion enhanced helicity in synthetic peptides containing unnatural , metal ligand residues. J. Am. Chem. Soc. 1990, 112, 9403- 9404
35) Messenger, A., Barclay, R., Bacteria, iron, and pathogenicity. Biochemical Education., 1983, 11, 54-63
36) Neilands, J., Siderophores: structure and function of microbial iron transport compounds. J. Bio. Chem., 1995, 270, 26723-26726
37) Atkin, C., Neilands, J., Rhodotorulic acid, a diketopiperazine dihydroxamic acid with growth-factor activity. I. Isolation and characterization. Biochemistry, 1986, 7, 3734–3739
38) Ankenbauer, R., Toyokuni, T., Staley, A., Rinehart, K., Cox, C., Synthesis and biological activity of pyochelin, a siderophore of Pseudomonas aeruginosa. J Bacteriol. 1988, 170, 5344–5351

16
CHAPTER TWO MELITTIN ANALOGUES
Melittin
Melittin is a hemolytic AMP isolated from the European honey bee Apis
mellifera. Melittin is the primary toxic component in bee venom and accounts for
approximately 50% of the dry mass. Melittin, figure 2.1, is a short linear peptide
containing 26 amino acids (GIGAVLKVLTTGLPALISWIKRKRQQ-NH2). The residues
at the N-terminus are mostly hydrophobic while the residues at the C-terminus are mainly
hydrophilic due to the basic amino acids. The properties of melittin have promoted its use
as a model peptide for hemolytic peptides.
Figure 2.1: The structure of melittin.
Melittin, as with other cytolytic peptides, is mainly hydrophobic, however the
peptide has a net charge of +6 at physiological pH. Most of this charge, +4, is located on
the highly basic C-terminus containing Lys-Arg-Lys-Arg. Figure 2.2 shows that most of
the positive charge resides on one side of the peptide helix which provides the

am
h
b
h
in
F
ce
ag
an
p
mphiphilic c
ighly import
etween the p
ead groups o
nteractions b
igure 2.2: H
In aqu
ertain condit
ggregate into
nd high salt
eptide and a
character alo
tant for the m
peptide and t
of the memb
become weak
Helical wheel
ueous solutio
tions the pep
o tetramers.
content.2 Th
allows for the
ong with othe
mechanism o
the membran
brane; withou
ker.
l diagram of
on, melittin e
ptide can be
These condi
hese conditio
e formation
er hydrophil
of action.1 A
ne are betwe
ut the cation
f melittin.
exists as a m
induced to f
itions includ
ons negate th
of aggregate
lic groups. T
As stated befo
een the catio
nic charges in
monomeric ra
form a helica
de high pH, h
he charge rep
es. In the hel
The charged C
ore, the initi
nic residues
n melittin, th
andom coil.2
al secondary
high melittin
pulsion betw
lical conform
C-terminus i
al interaction
and anionic
hese and the
2 However, u
y structure an
n concentrati
ween melittin
mation, melit
17
is
ns
c
under
nd
ions,
n
ttin

co
sh
F
ac
qu
is
is
co
h
as
m
ontains two
hape, figure
igure 2.3: 3D
Melittin’
ctivity is me
uickly to ent
s approximat
s attributed t
ontains 1.8x
owever evid
In additio
s well. Melit
melittin’s min
α-helical sub
2.3.
D representa
s primary ta
elittin’s distin
throcytes. Th
tely 1-2 µM
o the large a
x107 binding
dence suppor
on to hemoly
ttin is active
nimum inhib
bunits separa
ation of meli
Ac
arget is the en
nguishing ch
he melittin c
depending o
amount of bi
sites.4 The b
rts the torida
ytic activity,
against gram
bitory concen
ated by a pro
ittin showing
ctivity of M
nthrocyte me
haracteristic.
concentration
on experime
nding sites f
binding mec
al pore mode
, melittin pos
m-positive a
ntration (MI
oline-14 resi
g the “bent r
elittin
embrane and
.3 At lower c
n needed to
ental conditio
for melittin.
hanism has b
el described
ssesses a str
and gram-neg
IC) against S
idue to give
rod”.
d, as such, th
concentration
cause 50% h
ons.3 The low
For instance
been though
in chapter 1
rong antibact
gative bacter
S. Aureus is 3
a “bent-rod”
he hemolytic
ns melittin b
hemolysis (H
w concentrat
e, an enthroc
h much deba
.5,6
terial compo
ria. For exam
3.6 µM and
18
”
c
binds
HC50)
tion
cyte
ate,
onent
mple,

19
against E. Coli, 3.9 μM.7 The exploitation of this low MIC and the reduction of hemolytic
activity has been the focus of much research.
Structural Requirements for Activity Several extensive studies of melittin have determined that the hemolytic and
antibacterial activities are independent of one another. In the early structural activity
relationship studies in to the requirements of melittin it was determined the N-terminal 1-
20 residues play mainly a structural role in melittin’s absolute activity.8 Additionally, it is
determined that the C-terminal 21-26 residues are a necessary component for hemolytic
activity.8 However, more recent studies have proven that this is not the case.
In a comprehensive study by Blondelle and Houghten, the necessary residues
were discovered for melittin’s hemolytic and antibacterial properties. The individual
omission of Leu-6, Leu-9, Leu-13, or Leu-16 give an over 100-fold decrease in hemolytic
activity.9 This is also evidence to support the need for the leucine zipper design for
aggregation in the plasma membrane. The omission of Ile-17 and Trp-19 also leads to
considerable decrease in hemolytic activity. However, the omission of the previously
described amino acids did not have the same drastic effects on the antibacterial activity.
Only a 2-fold or 3-fold decrease in antibacterial activity was observed in contrast to the
100-fold seen in the hemolytic studies.
Blondelle and Houghten also explored an individual leucine substitution of each
amino acid in the melittin peptide. It is interesting to note that at 19 amino acids that were
substituted had considerably higher hemolysis percentage than the unadulterated

20
melittin.10 Two amino acids of note, Lys-7 and Trp-19 had detrimental effects on the
hemolytic activity when substituted, even at the highest concentrations tested.10
Unfortunately, the antibacterial activity was not probed with these analogues.
A core requirement for hemolytic activity is the amphiphilic helical secondary
structure, yet the helical structure is not a requirement for antibacterial activity. In a study
by Oren and Shai, four amino acid residues, Val-5, Val-8, Iso-17 and Lys-21, were
replaced with D-isomers of the respective amino acids. The melittin analogue showed a
low propensity to form a helical secondary structure. Because the peptide had low
propensity for helix formation, the hemolytic activity was also low, yet there was still a
significant level of antibacterial activity.11
Truncated Melittin
As described previously, it is documented that the N-terminus plays a minimal
role with the mechanism for antibacterial activity. As such, researchers have explored
using truncated melittin fragments to determine their viability as therapeutics. Extensive
studies have focused on the fragment around residues 12-25, figure 2.4, containing the C-
terminal basic amino acids.12-14 The peptide has a lower propensity to form a helical
secondary structure which makes it an ideal lead compound to study. As a result, the
hemolytic activity is reduced while the antibacterial activity is retained.

21
Figure 2.4: The structure of truncated melittin(12-25).
1st Genertation Melittin Derivatives
The current project explored the coupling of melittin, metal destabilization, and
siderophores as targeted antimicrobial peptides.. Previous studies have shown that Thr-
10 and Lys-21 are not critical for antibacterial activity, so, in the present project, these
sites were selected for incorporation of ligands for metal binding. Thr-10 was changed to
Lys-10 to be able to couple the ligand to the peptide at residues 10 and 21. Two ligands
were selected to determine the possible differences between bidentate ligand and
tridentate ligands.
Figure 2.5: Placement of metal binding moieties to melittin, where X represents bipyridine or terpyridine.
The design of this work was to use metal chelation to deactivate the peptide’s
hemolytic activity and retaining antibacterial activity with a metal-ligand switch. The
targeted activity will activate once a siderophore from a bacteria will remove the metal
ion and reactivate the peptide. Since siderophores are released from bacteria must remain

in
p
p
re
F
n close proxi
eptide to atta
eptide will b
eleased.
igure 2.6: M
imity to be a
ack the bacte
be deactivate
Metal assisted
absorbed bac
eria instead
ed with a me
d deactivatio
ck into a bac
of healthy ce
etal ion, and
on of peptide
cterium.15 Th
ells. Figure 2
then reactiv
e with sidero
he proximity
2.6 represen
vated when th
ophore reacti
y will allow t
nts how the
he metal ion
ivation.
22
the
n is

23
Synthesis of Metal Binding Moieties
Scheme 2.1: Synthesis of metal binding ligands.
To preserve the positive charge required for melittin activity, two neutral metal
binding ligands were chosen as chelators, 4-methyl-2,2'-bipyridine-4'-carboxylic acid
and 2,2’:6’, 2”-terpyridine-4’-carboxylic acid, scheme 2.1. The carboxylic acid moieties
will be used to couple the ligand to a lysine residue with the modified melittin. The
compounds were synthesized according to literature procedures. To synthesize the
bipyridyl compound 3, commercially available compound 1 was selectively oxidized
with selenium dioxide. Compound 2 was further oxidized with silver oxide to afford the
carboxylic acid in compound 3.16 The terpyridyl compound 6 was synthesized by treating
commercially available citrazinic acid (4) with phosphorous oxybromide to produce
compound 5.17 The dibromide compound 5 was then used in a Stille coupling reaction
using a palladium catalyst and 2-(tributylstannyl)pyridine to provide the desired final
product, compound 6.18 The methyl ester is hydrolyzed during the work up procedure.

24
Synthesis of Melittin Derivatives
Scheme 2.2: Synthesis of melittin derivatives, where X is terpyridine or bipyridine.
The melittin derivatives MLT-bpy and MLT-tpy, figure 2.5, were synthesized by
solid phase peptide synthesis following the Fmoc based protocol. 4-methyltrityl (Mtt)
protected lysine was used to selectively allow binding of the ligand to the desired lysine
residue. Other lysine residues were protected with the allyloxycarbonyl (alloc) protecting
group. The solid support for the synthesis was the ChemMatrix rink amide resin with a
0.52 mmol/g loading. The coupling reactions were monitored using the ninhydrin test
(Kaiser test) for primary amines and the chloranil test for secondary amines. Once the
peptide was completed, the Mtt protecting groups were removed with 1.8%
trifluoroacetic acid in methylene chloride. The reaction mixture was drained and a yellow
solution indicated the removal of the protecting group. More deprotection cocktail was
added and repeated until little to no color appeared in the reaction mixture. The ligand
compounds were coupled to the free amine of the deprotected lysine using standard

25
HBTU coupling. The remaining Alloc protecting groups were deprotected using a
palladium catalyzed decoupling procedure. The peptides were cleaved from the resin with
non-acid liable protecting groups removed with a cleavage cocktail composed of 95%
TFA, 2.5% TIPS, 2.5% water. The peptides were then purified by reverse phase HPLC
and characterized Matrix-assisted laser desorption ionization (MALDI) mass
spectroscopy, table 2.1.
Table 2.1: Purification of melittin derivatives.
Compound HPLC
%acetonitrile/waterRetention
Time Actual Mass
Observed Mass
MLT-bpy 35-70 25 min 3308.04 3308.99
MLT-tpy 45-80 27 min 3434.09 3435.56
Circular Dichroism
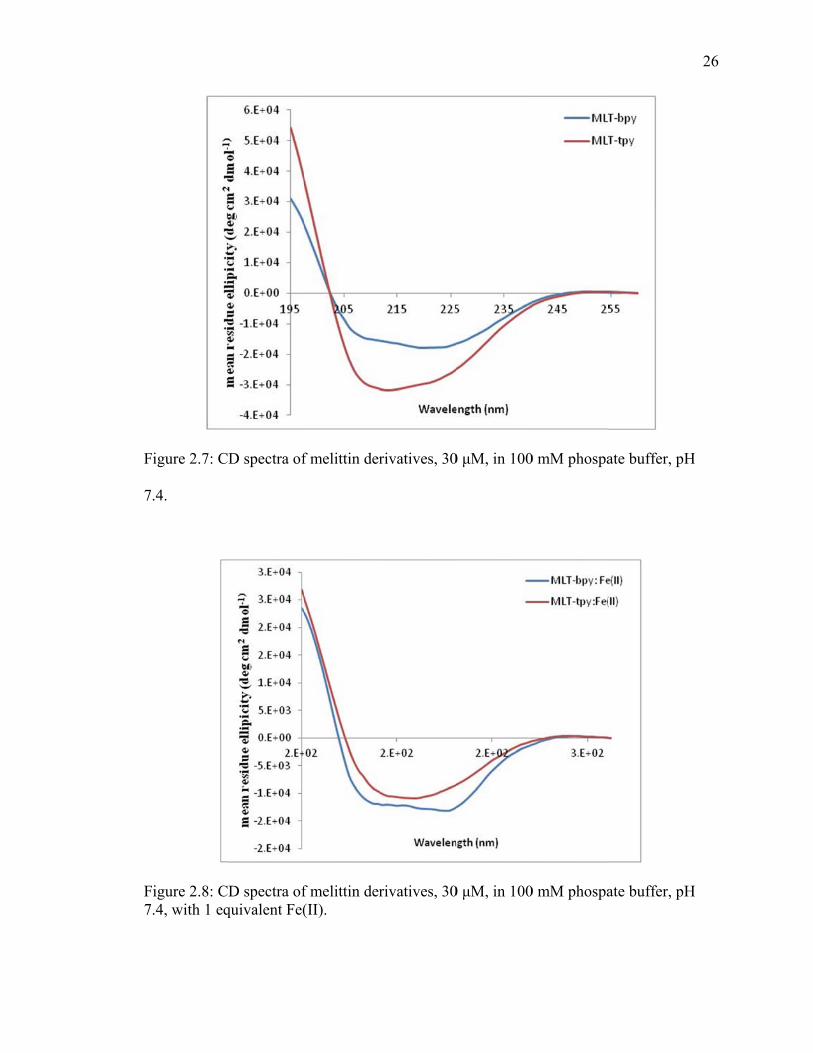
Circular dichroism experiments were conducted to determine helicity of the
designed peptides, figure 2.7. In 100mM phospate buffer at pH 7.4 showed that the
peptide was found to form an α-helix. Interestingly, when iron (II) as added to the
peptides in solution, little change was observed in the helicity of MLT-bpy, but the
helical characteristics were somewhat lost with MLT-tpy.
F
7
F7
igure 2.7: C
.4.
igure 2.8: C.4, with 1 eq
D spectra of
D spectra ofquivalent Fe(
f melittin der
f melittin der(II).
rivatives, 30
rivatives, 30
0 μM, in 100
0 μM, in 100
0 mM phospa
0 mM phospa
ate buffer, pH
ate buffer, pH
26
H
H
n
4
U
un
ac
F
se
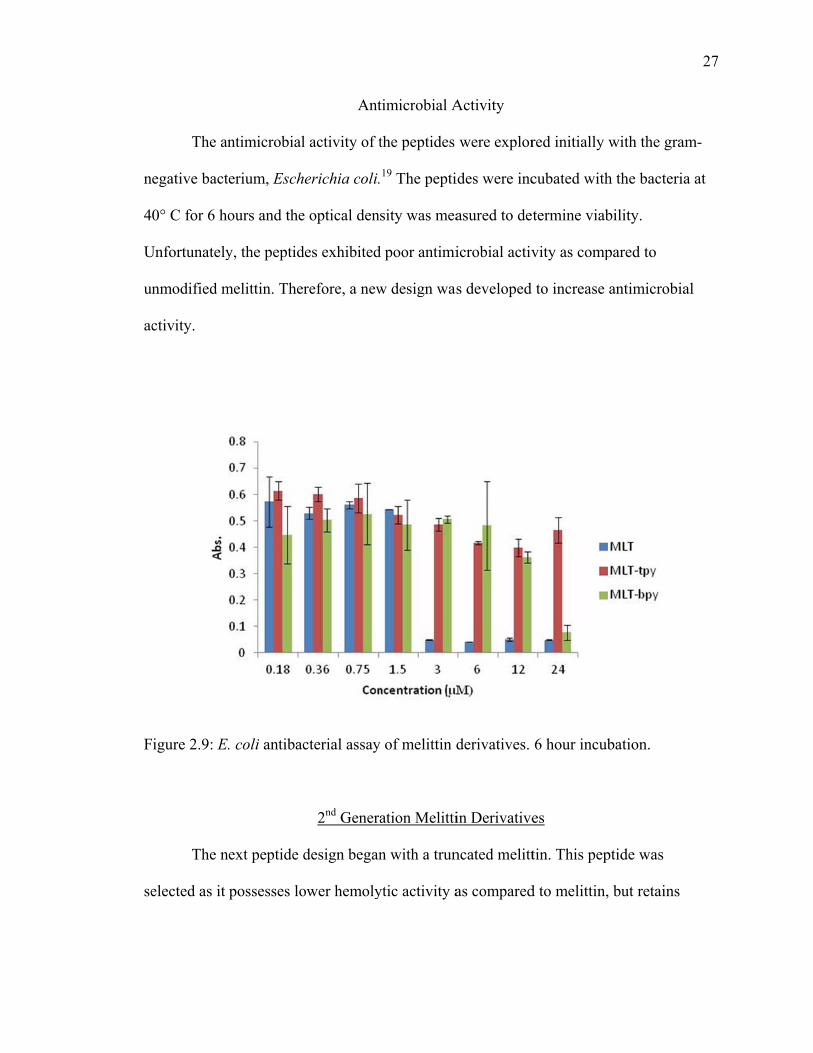
The an
egative bact
0° C for 6 ho
Unfortunately
nmodified m
ctivity.
igure 2.9: E.
The n
elected as it
ntimicrobial
erium, Esch
ours and the
y, the peptid
melittin. Ther
. coli antibac
ext peptide d
possesses lo
Ant
l activity of t
erichia coli.
e optical dens
des exhibited
refore, a new
cterial assay
2nd Genera
design began
ower hemoly
timicrobial A
the peptides
19 The pepti
sity was mea
d poor antimi
w design was
y of melittin d
ation Melitti
n with a trun
ytic activity a
Activity
were explor
ides were inc
asured to de
icrobial activ
s developed
derivatives.
in Derivative
ncated melitt
as compared
red initially w
cubated with
termine viab
vity as comp
to increase
6 hour incub
es
tin. This pep
d to melittin,
with the gra
h the bacteria
bility.
pared to
antimicrobia
bation.
ptide was
but retains
27
am-
a at
al

28
similar antibacterial activity. The metal ligands were also moved to the terminus of the
peptide in an effort to have less effect on activity. Four truncated peptides, figure 2.10;
containing melittin residues 12-25 with metal binding ligands were synthesized to
determine their antibacterial activity with and without metal ions. Two of the peptides
contained a single ligand on each of the C- and N- termini of the peptides as shown
below. The third peptide contained ligands at both termini. We also prepared a truncated
12-25 residue melittin fragment as a control peptide. The same synthetic strategy was
used to attach the ligand to the lysine residues. A glycine spacer was between the
truncated peptide and the ligand-modified lysine residues at the termini.
Figure 2.10: Second generation melittin derivatives. From top to bottom: MLTx, MLTx1, MLTx2, MLTx3. Where X is terpyridine.
Synthesis of 2nd Generation Melittin Derivatives
The new peptides were synthesized in the same manner as described previously.
Terpyridine and bipyridine were used as ligands, however, the coupling of of the
bipridine MLTx1 proved to be difficult and was abandoned, nevertheless, MLTx2-bpy

29
was completed. Given that bipyridine is a bidentate ligand and would most likely bridge
multiple peptides leading to higher ordered structures, only terpyridine was used in the
synthesis of MLTx3. The peptides were then purified by reverse phase HPLC and
characterized Matrix-assisted laser desorption ionization (MALDI) mass spectroscopy,
table 2.2.
Table 2.2: Purification of truncated melittin derivatives.
Compound HPLC
%acetonitrile/waterRetention
Time Actual Mass
Observed Mass
MLTx 10-50 38 min 1887.28 1886.45 MLTx1-tpy 10-40 48 min 2109.52 2110.01 MLTx2-tpy 20-50 30 min 2110.98 2109.96 MLTx3-tpy 20-60 26 min 2554.01 2553.54
UV/Vis spectroscopy
UV/Vis spectroscopy was used to monitor that the ligands within the peptide are
chelating the metal. There is a distinctive charge transfer that occurs between terpyridine
and iron(II), with the λmax for the terpyridine-iron(II) complex occurring at around
570nm. The spectra below show that the charge transfer is occurring, demonstrating that
the peptides do bind to the metal ions.
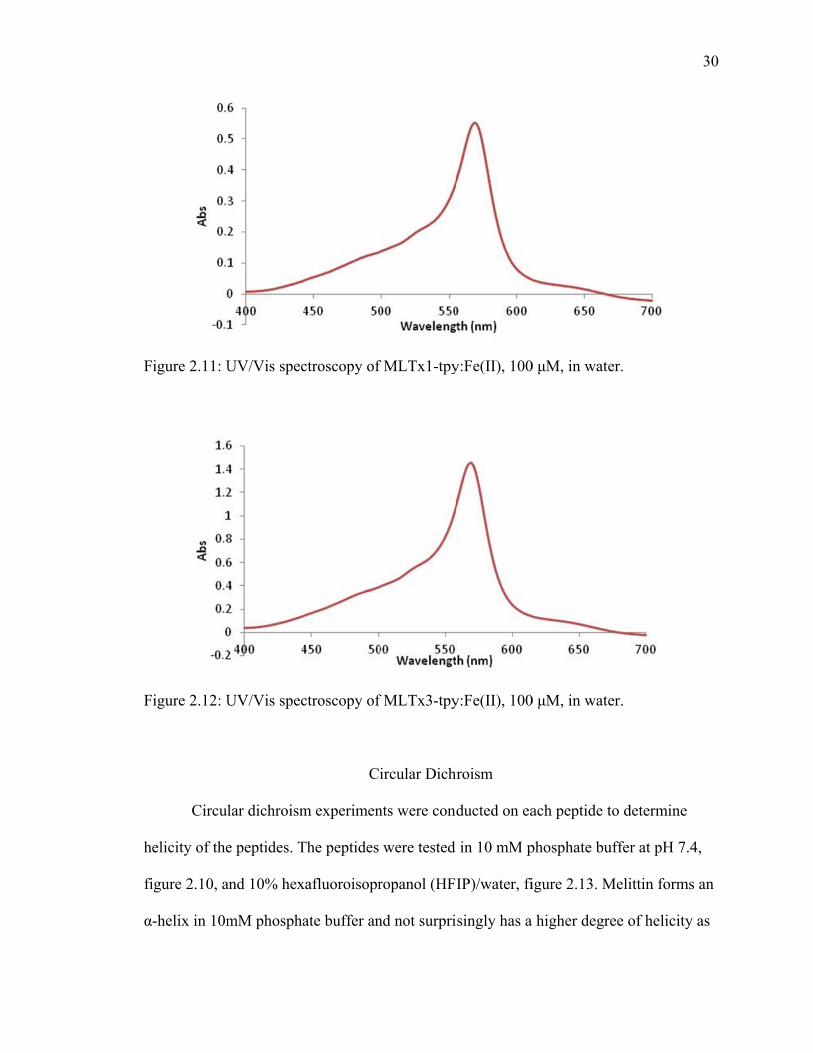
F
F
h
fi
α
igure 2.11: U
igure 2.12: U
Circul
elicity of the
igure 2.10, a
-helix in 10m
UV/Vis spec
UV/Vis spec
lar dichroism
e peptides. T
and 10% hex
mM phospha
ctroscopy of
ctroscopy of
Ci
m experimen
The peptides
xafluoroisopr
ate buffer an
f MLTx1-tpy
f MLTx3-tpy
ircular Dichr
nts were cond
were tested
ropanol (HFI
nd not surpri
y:Fe(II), 100
y:Fe(II), 100
hroism
ducted on ea
in 10 mM p
IP)/water, fi
singly has a
0 μM, in wate
0 μM, in wate
ach peptide t
phosphate bu
igure 2.13. M
higher degr
er.
er.
to determine
uffer at pH 7
Melittin form
ree of helicit
30
e
7.4,
ms an
ty as

31
it has a longer sequence. The truncated melittin fragments did not possess appreciable
helicity in the 10 mM phosphate buffer, but, unexpectedly MLTx3-tpy did possess
moderate helicity in 10 mM phosphate buffer. Furthermore, all peptides showed
appreciable helicity in 10% HFIP, figure 2.14. Hexafluoroisopropanol is commonly used
to induce helicity of peptides which form α- helical secondary structures.
When metal ions were added to the peptides, figure 2.14, it was observed that the
helix is destabilized in solution. Interestingly, the denatured peptide appears to retain
more helical characteristics. The peptide was denatured in 10 mM urea solution, then iron
(II) was added to solution. The urea was removed from solution by dialysis filtration. The
denaturing of the peptide was necessary because of the possibility that the peptide may be
partially formed into a helix. This might contribute to a higher than normal antibacterial
activity.
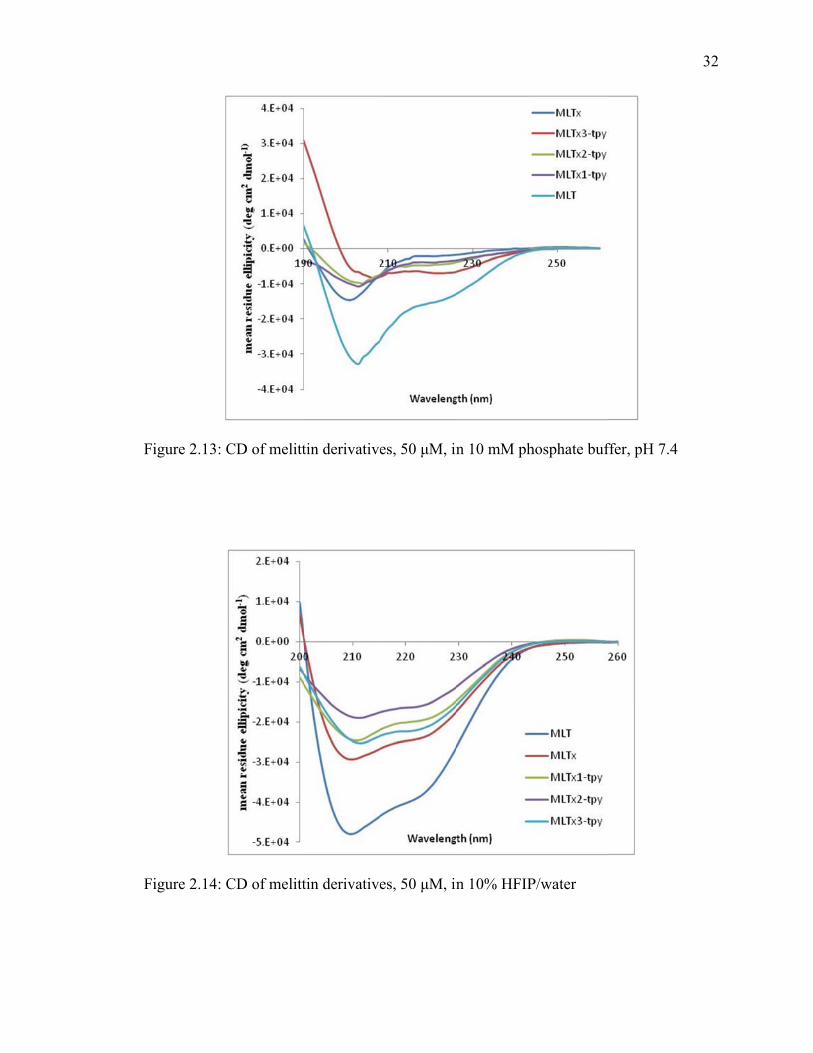
F
F
igure 2.13: C
igure 2.14: C
CD of melitt
CD of melitt
tin derivative
tin derivative
es, 50 μM, in
es, 50 μM, in
n 10 mM ph
n 10% HFIP
hosphate buf
P/water
ffer, pH 7.4
32
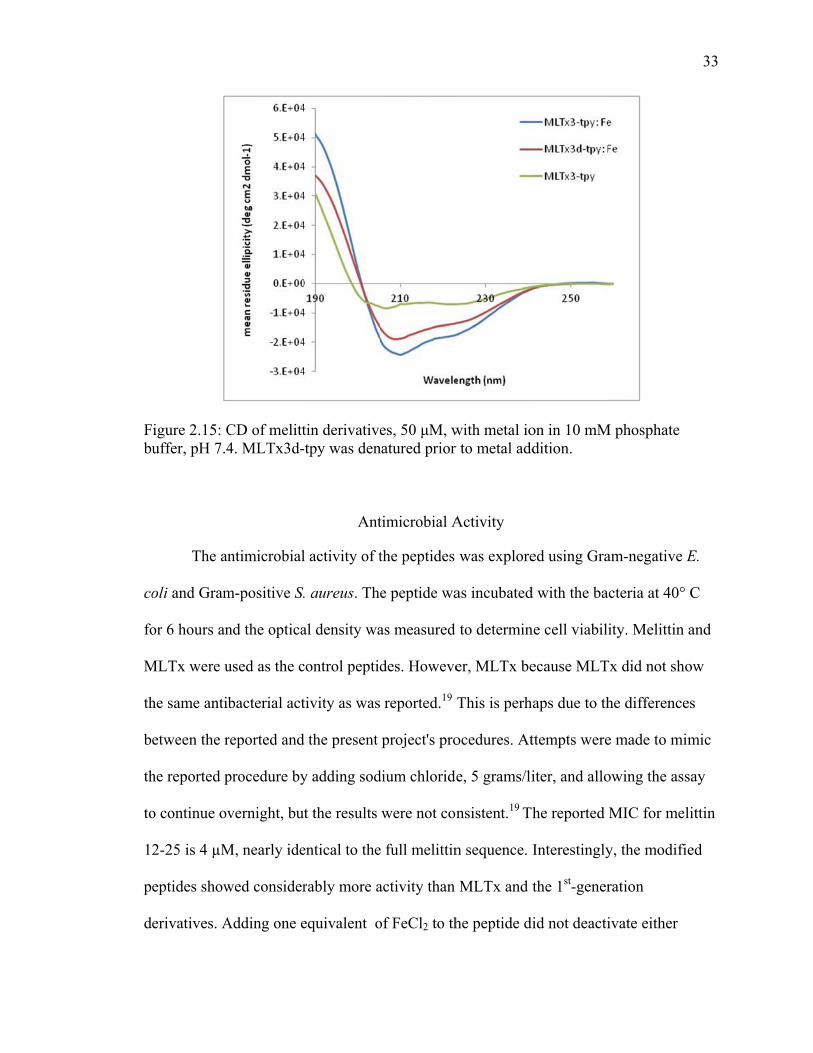
Fbu
co
fo
M
th
b
th
to
1
p
d
igure 2.15: Cuffer, pH 7.4
The an
oli and Gram
or 6 hours an
MLTx were u
he same antib
etween the r
he reported p
o continue ov
2-25 is 4 µM
eptides show
erivatives. A
CD of melitt4. MLTx3d-
ntimicrobial
m-positive S.
nd the optica
used as the c
bacterial act
reported and
procedure by
vernight, but
M, nearly ide
wed consider
Adding one e
tin derivativetpy was den
Ant
l activity of t
. aureus. The
al density wa
control peptid
tivity as was
d the present
y adding sod
t the results
entical to the
rably more a
equivalent o
es, 50 μM, wnatured prior
timicrobial A
the peptides
e peptide wa
as measured
des. Howeve
reported.19
project's pro
dium chloride
were not con
e full melittin
activity than
of FeCl2 to th
with metal ior to metal add
Activity
was explore
as incubated
to determin
er, MLTx be
This is perh
ocedures. At
e, 5 grams/li
nsistent.19 Th
n sequence. I
MLTx and t
he peptide di
on in 10 mMdition.
ed using Gra
d with the ba
ne cell viabili
ecause MLT
aps due to th
ttempts were
iter, and allo
he reported
Interestingly
the 1st-gener
id not deacti
M phosphate
am-negative
cteria at 40°
ity. Melittin
Tx did not sh
he difference
e made to mi
owing the as
MIC for me
y, the modifi
ration
ivate either
33
E.
° C
and
how
es
imic
say
littin
ied

34
MLTx1 or MLTx2 containing the terpyridine ligand. However, some decrease in
antibiotic activity did occur with MLTx3-tpy. A denatured peptide, MLTx3d-tpy, was
also tested. The peptide was first denatured in 10mM urea, then the metal ion was added
to solution. The solution was than dialyzed to remove the urea from the solution. By
denaturing the peptide, the activity of the peptide-metal complex (MLTx3d: Fe(II)) was
lowered 2-fold as compated to the non-denatured sample (MLTx3: Fe(II)).
Table 2.3: MIC of melittin derivatives.
Peptide E.Coli S. Aureus
Melittin 4 μM ± 0.002 1 μM ± 0.006
MLTx >64 μM ± 0.02 -
MLTx1-tpy 16 μM ± 0.001 8 µM ± 0.019
MLTx2-tpy 16 μM ± 0.02 8 µM ± 0.02
MLTx3-tpy 8 μM ± 0.012 8 µM ± 0.009
MLTx1-tpy: Fe(II) 16 μM ± 0.08 8 μM ± 0.02
MLT2x-tpy: Fe(II) 16 μM ± 0.02 8 μM ± 0.02
MLTx3-tpy: Fe(II) 16 μM ± 0.006 8 μM ± 0.01
MLTx3d-tpy: Fe(II) 32 μM ±0.020 16 μM ± 0.02
Reversibility of the Metal-Ligand Interaction
Terpyridine-Fe(II) interactions are known to be reversible in polymers and
peptides.21,22 However, attempts to remove the metal ion from the melittin derivatives has
proven to be very difficult. Several solvents and chelators were used to attempt the
removal of the metal ions, figure 2.16. Water, methanol, and dimethylformamide (DMF)

w
an
T
w
FF
on
M
U
o
C
sh
were the solv
nd N-(hydro
The solutions
was only brok
igure 2.16: Me(II) comple
Other
nly cobalt af
MLTx3-tpy w
UV/Vis spect
ccurred. Th
Co(II) to the p
howed little
vents used, w
oxyethyl)-eth
s were heated
ken with the
MLTx3-tpy ex with 2 equ
possible for
fforded a me
was analyzed
tra of MLTx
he CD spectr
peptide dest
difference in
while deferox
hylenediamin
d to 50° C an
e peptides in
with 1 equivuivalents of
Melittin De
r metal ions
etal-ligand ch
d with 1 equi
x3-tpy: Co(II
ra of MLTx3
abilized the
n the spectru
xamine, ethy
netriacetic a
nd kept for 1
DMF as can
v Fe(II) in sochelators.
rivatives wit
were explor
harge transfe
ivalent of m
I), in shows t
3-tpy: Co(II)
helix, and d
um.
ylenediamine
acid (HEDTA
18 hours. Th
n been seen b
olution. 500
th Other Me
red: cobalt, c
fer that was o
metal ion as p
the metal lig
, figure 2.18
denaturing th
etetraacetic a
A) were the c
he metal-liga
by the comp
µM solution
etals
copper, and n
observable, f
previously de
gand charge
8, shows that
he peptide wi
acid (EDTA
chelators use
and interactio
plete color lo
n of the pept
nickel. How
figure 2.17.
escribed.The
transfer that
t adding the
ith a 10 mM
35
A),
ed.
on
oss.
tide-
ever,
e
t
M urea
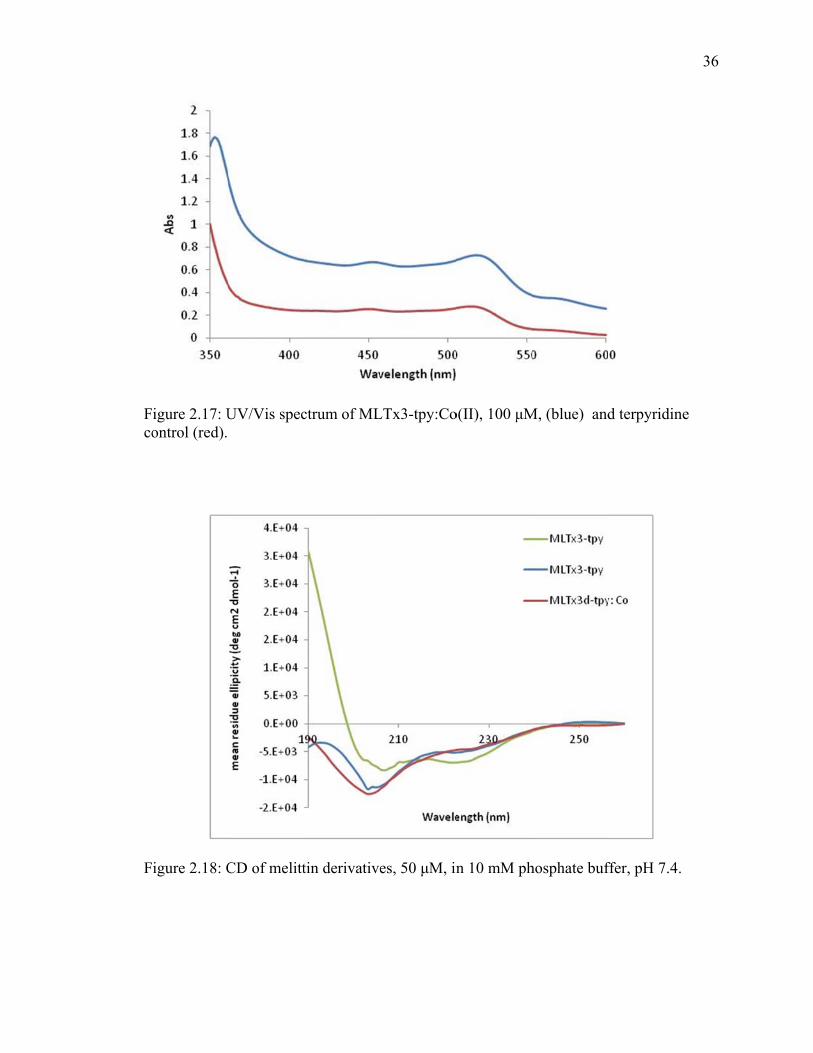
Fco
F
igure 2.17: Uontrol (red).
igure 2.18: C
UV/Vis spec
CD of melitt
ctrum of ML
tin derivative
LTx3-tpy:Co
es, 50 μM, in
o(II), 100 μM
n 10 mM ph
M, (blue) and
hosphate buf
d terpyridine
ffer, pH 7.4.
36
e

37
Conclusions
Melittin derivatives are susceptible to small changes which have major impact to
their activity. The 1st-generation peptides ultimately failed to maintain any antibacterial
activity. The addition of the aromatic ligands at residues 10 and 21 was enough to reduce
the antibacterial activity 30-fold. However, the truncated peptides show promise as novel
antibacterials as they have retained most of their activity. Unfortunately, the difficulty of
reversing the metal-ligand interaction is a problem that needs to be solved to move the
project further.
Materials and Methods
All peptides were synthesized on Rink amide resin (200-400 mesh) purchased
from novabiochem. Critizanic acid and 4,4'-dimethyl-2,2' bipyridine were both
purchased from VWR International. Other reagents used were purchased from Sigma-
Aldrich or VWR International. The peptides were purified and analyzed with Waters
Delta Prep 4000 HPLC. The ligand intermediates and final compounds were
characterized by NMR spectroscopy with the Varian Inova-300 Mhz nuclear magnetic
resonance (NMR) spectrometer. All peptides were characterized by mass spectrocopy
using an Applied Biosystems Voyager-DE Matrix-assisted laser desorption ionization –
time of flight mass spectromerter. Circular dichroism experiments were performed on a
Jasco J-810 circular dichroism spectrophotometer. UV/Vis spectra were obtained on a
Cary 100 Bio UV-visible spectrophotometer.

38
Experimental
4-methyl-2,2'-bipyridine-4'-carboxaldehyde(2) To a dry 500 ml round bottom flask
was added 4,4'-dimethyl-2,2' bipyridine (1) (5.00 g, 27 mmol) followed by 200 ml
dioxane. The solution was sparged with nitrogen for 15 minutes with stirring. Selenium
dioxide (3.31 g, 30 mmol) was then added and the solution was sparged while stirring for
15 minutes. The solution was heated to a gentle reflux for 24 hours. After cooling the
reaction mixture was vacuum filtered and the solvent removed. The remaining solid was
dissolved in 100 ml ethyl acetate and refluxed for an additional 1 hour and vacuum
filtered while hot. The filtrate was washed with 0.1 M sodium carbonate followed by 0.3
M sodium metabisulfate. The pH was then adjusted to 10 with solid sodium carbonate
and then extracted with methylene chloride. (yield: 1.67 g, 31%) 1H-NMR (300 MHz,
CD2Cl2) δ 10.19 (s, 1H), 8.84(d, 1H), 8.80(s, 1H), 8.52(d, 1H), 8.29(s, 1H), 7.71(d, 1H),
7.22(d, 1H), 2.47(s, 3H)
4-methyl-2,2'-bipyridine-4'-carboxylic acid(3) To a dry 250 ml round bottom flask
was added 95% ethanol/water solution followed by 4-methyl-2,2'-bipyridine-4'-
carboxaldehyde (2) (1.67 g, 8.40 mmol). An aqueous 20 ml solution of silver nitrate (
1.56 g, 9.20 mmol) was then added. A 1 M solution of sodium hydroxide (1.67 g, 42
mmol) was added via an addition funnel for 20 minutes and the reaction mixture was
stirred for 24 hours. The ethanol was removed by reduced pressure. The reaction mixture
was then vacuum filtered and the solids washed with 2 M sodium hydroxide followed by
water. The filtrate was extracted with methylene chloride. The pH of the aqueous solution
was adjusted to 3. The lower pH allowed a white precipitate to form and was collected.

39
(yield: 0.746g, 41%) 1H -NMR (300 MHz, (CD3)2SO) δ 8.86(d, 1H), 8.81(s, 1H), 8.57(d,
1H), 8.26(s, 1H), 7.86(d, 1H), 7.33(d, 1H), 2.48(s, 3H)
2,6-Dibromoisonicotonic acid methyl ester (5) Phosphorous oxybromide (10 g, 35
mmol) and citrazinic acid (4) (1.7 g, 11 mmol) were ground together, put in a dry 100 ml
round bottom flask and purged with nitrogen. The reaction mixture was heated to 180
ºC under an nitrogen atmosphere for 90 min. The mixture was then cooled to room
temperature and quenched with anhydrous MeOH (10 mL). After dilution with saturated
aqueous K2CO3 (20 ml), the solution was extracted with dichloromethane (3x20 ml). The
organic layer was washed with brine and dried over sodium sulfate. The solution was
decanted into a 1 inch silica plug and washed with DCM (3x15 ml). The filtrate solution
was evaporated under lower pressure. No further purification was necessary. (yield: 2.22
g, 68.5%) 1H -NMR (300 MHz, CD2Cl) δ 8.00(s, 2H), 3.97(s, 3H)
2,2':6', 2"-terpyridine-4'-carboxylic acid (6) 2,6-Dibromoisonicotonic acid methyl
ester (1.6 g, 5.4 mmol), tributylstannyl pyridine (3.51 ml, 10.8 mmol), and
tetrakis(triphenylphosphine)palladium (0.63 g, 0.54 mmol) were dissolved in 50 ml dry
toluene in a dry 150 ml round bottom flask. The reaction mixture was heated to 110° C
under nitrogen for 16 hours. The reaction was monitored by TLC. Once the reaction was
complete, the mixture was cooled to room temperature and quenched with saturated
ammonium chloride (30 ml) and stirred for additional 30 minutes. The reaction mixture
was filtered over celite and the filtered solids washed with DCM. The aqueous phase was
extracted with DCM (3x15 ml). The organic phases were dried with sodium sulfate and
the solvent removed. Concentrated hydrochloric acid (15 ml) was added to the residue to
hydrolyze the methyl ester. The mixture was neutralized with 2 M sodium hydroxide and

40
extracted with DCM (3x10 ml). The organic layer was separated and the solvent
evaporated. A 2/1 water/methanol mixture (25 ml) was added to the residue. 2 M sodium
hydroxide solution (15 ml) was added and the mixture was stirred for 1 hour, this was to
complete the hydrolysis of the methyl ester. The solution was extracted with DCM. The
aqueous layer was then acidified to pH 3 and then the methanol removed under reduced
pressure. The pH was adjusted to 6 which allowed a white precipitate to form. (yield:
0.983 g, 65.6%) 1H -NMR (300 MHz, CD3OD) δ 8.86(s, 2H), 8.71(m, 4H), 8.02(td, 2H),
7.49(qd, 2H)
General Synthesis of Peptides. In a 20 ml peptide synthesis flask was added 200 mg
Rink-Amide resin (0.052 mmol/g). The peptide was shaken with 10ml DCM to swell the
resin. Fmoc-protected amino acids (6 equiv), HBTU (6 equiv), DIEA (12 equiv) were
dissolved in 10 ml DMF and added to the resin. The synthesis flask was spun for 4 hours.
The resin was washed with DMF, methanol, and DCM (3x10 ml). Piperidine (25% in
DMF, 10 ml) was added to the resin and spun for 20 minutes and then drained. The
peptide was washed with DMF, methanol, and DCM (3x10 ml). The procedure was
continued until the peptide was complete.
Kaiser Protocol Approximately 20 resin beads were removed from the synthesis flask
and placed in a small culture tube. Three drops of ninhydrin solution (1.0 g of ninhydrin
in 20 mL of n-butanol), three drops of phenol solution (40 g of phenol in 20 mL of n-
butanol), and three drops of KCN solution (dissolve 16.5 mg of KCN in 25 mL of
distilled water. Dilute 1.0 mL of KCNaq solution with 49 mL of pyridine) were added to
the culture tube. The mixture was added to a beaker of boiling water. If the beads turned

41
blue there is a free primary amine. If there is no color change or if the beads turn slightly
brown there are not any free primary amines present.
Chloranil Protocol Approximately 20 resin beads were removed from the synthesis flask
and placed in a small culture tube. Three drops of chloranil solution (2% chloranil in
DMF) and three drops of acetylaldehyde solution (2% acetylaldehyde in DMF) were
added to the culture tube. The mixture was allowed to sit at room temperature. A blue-
green beads indicates a free secondary amine. A clear resin indicated were not any free
secondary amines present.
4-Methyltrityl (Mtt) deprotection The resin was washed with DCM then 15 ml of 1.8%
TFA in chloroform and then added to the resin. The resin was allowed to stir for 4
minutes. The yellow solution was drained and the resin washed with DCM. The
procedure was continued until little yellow color evolved from the solution.
Coupling of metal-chelating ligand The resin was washed with DMF. The ligand (6
equiv), HBTU (6 equiv), DIEA (12 equiv) were dissolved in 10 ml DMF and added to the
resin. The resin was spun for 4 hours then washed with DMF, methanol, and DCM (3x10
ml). The resin was tested to ensure complete coupling. If a positive Kaiser test occurred,
a second coupling procedure was performed.
Aloc deprotection The resin was washed with chloroform and an aloc cleavage cocktail
was added to the resin in 15 ml chloroform/acetic acid (0.5 mL per gram of resin), N-
methylmorpholine (2 mL per gram of resin), and Pd(PPh3)4 (0.3 equivalents based on
resin substitution). The resin was spun for 45 minutes and analyzed by MALDI. If the
reaction was not complete, the procedure was repeated if necessary.

42
CD Protocol A peptide sample was taken from the stock solution and diluted in 10 mM
phosphate buffer solution pH 7.4 or 10% HFIP in water to make a 200 µL solution. The
sample solution was scanned from 260 nm to 190 nm using a 1 nm bandwidth.
UV/Vis Protocol A peptide sample was taken from the stock solution and diluted with DI
water to a volume of 500 µL. 1 µL of a stock FeCl2 solution was added to the peptide
solution and a color change was observed. The solution was then scanned.
Denaturation Protocol A sample of stock peptide solution was diluted in a 10 mM urea
solution. After 15 min 1 equivalent of a stock FeCl2 solution was added to the solution
and a color change observed. The solution was then added to a dialysis tubing. The
tubing was placed in a 1 L beaker containing DI water. The solvent was changed over 30
minute intervals twice. The solvent was changed again and left overnight.
Antibacterial Assay Procedure Bacteria were grown on a TSB agar culture plate
overnight at 40° C. The bacteria were inoculated in 50 ml TSB media until the OD600 was
between 0.4 and 0.6. 5 ml of the culture media was centrifuged and washed with MHB
media (3x 5 ml). The culture media was diluted to an OD600 of 0.001. 90 µl of the
bacterial solution was pipetted on a 96 well plate along with 10 µl peptide solution. The
96 well plate was incubated at 40° C with shaking for 6 hours. The OD600 was taken and
the lowest concentration where there is minimal growth was decided to be the MIC.

43
References
1) Raghuraman, H., Chattopadhyay, A., Melittin: a membrane-active peptide with diverse functions. Biosci Rep., 2007, 27, 189–223
2) Tatham, A., Hider, R., Draket, A., The effect of counterions on melittin aggregation. Biochem. J., 1983, 211, 683-686
3) Tosteson, M., Holmes, S., Razin, M., Tosteson, D., Melittin lysis of red cells. J Membr Biol, 1985, 87, 35–44
4) Lee, T., Mozsolits, H., Aguilar, M., Measurement of the affinity of melittin for zwitterionic and anionic membranes using immobilized lipid biosensors. J. Pept. Res., 2001, 58, 464-476
5) Yang, L., Harroun, T., Weiss, T., Ding, L., Huang, H. Barrel-stave model or toroidal model? A case study on melittin pores. Biophysical Journal, 2001, 81, 1475–1485
6) Irudayam, A., Berkowitz, M., Influence of the arrangement and secondary structure of melittin peptides on the formation and stability of toroidal pores. Biochimica et Biophysica Acta, 2011, 2258–2266
7) Asthana N, Yadav SP, Ghosh JK (2004) Dissection of antibacterial and toxic activity of melittin: a leucine zipper motif plays a crucial role in determining its hemolytic activity but not antibacterial activity. J Biol Chem 279:55042–55050
8) Schroder, F., Lubke, K., Lehmann, M., Beetz, I., Haemolytic activity and action on the surface tension of aqueous solutions of synthetic melittins and their derivatives. Experientia, 1971, 27, 764-765
9) Blondelle, S., Houghten, R., Hemolytic and antimicrobial activities of the twenty-four individual omission analogues of melittin. Biochemistry, 1991, 30, 4671-4678
10) Blondelle, S., & Houghten, R., Probing the relationships between the structure and hemolytic activity of melittin with a complete set of leucine substitution analogues. Pept. Res. 1991, 4, 12-18
11) Oren, Z., Shai, Y., Selective lysis of bacteria but not mammalian cells by diastereomers of melittin: structure-function study. Biochemistry, 1997, 36, 1826-1835
12) Yan, H., Li, S., Sun, X., Mi, H., He, B., Individual substitution analogs of Mel(12–26), melittin’s C-terminal 15-residue peptide: their antimicrobial and hemolytic actions. FEBS Lett. 2003, 554, 100-104

44
13) Subbalakshmi, C., Nagaraj, R., Sitaram, N., Biological activities of C-terminal 15-residue synthetic fragment of melittin: design of an analog with improved antibacterial activity FEBS Lett. 1999, 448, 62 -66
14) Yan, H., Li, S., Sun, X., Mi, H., Chen, S., Fan, Y., Deletion of two C-terminal Gln residues of 12-26-residue fragment of melittin improves its antimicrobial activity. Peptides, 2005, 3,369-375
15) Messenger, A., Barclay, R., Bacteria, iron, and pathogenicity. Biochemical Education., 1983, 11, 54-63
16) Peek, B., Ross, G., Edwards, S., Meyer, G., Meyer, T., Erickson, B., Synthesis of redox derivatives of lysine and related peptides containing phenothiazine or tris(2,2'-bipyridine)ruthenium(II) Int. J. Pep. Protiens Res., 1991, 38. 114-123
17) Leblond, J., Gao, H., Petitjean, A., Leroux, J., pH-Responsive molecular tweezers. J. Am. Chem. Soc., 2010, 132, 8544–8545
18) Fallahpour, R., Carboxylate derivatives of oligopyridines. Synthesis, 8, 1138-1142
19) Kuriakose, J., Hernandez-Gordillo, V., Nepal, M., Brezden, A., Pozzi, V.,
Seleem, M., Chmielewski, J., Targeting intracellular pathogenic bacteria with unnatural proline-rich peptides: coupling antibacterial activity with macrophage penetration. Angew. Chem., 2013, 125, 9846 –9849
20) Yan, H., Li, S., Sun, X., Mi, H., Chen, S., Fan, Y., Deletion of two C-terminal Gln residues of 12-26-residue fragment of melittin improves its antimicrobial activity. Peptides, 2005, 3, 369-375
21) Schmatloch, S., Gonza´lez, M., Schubert, U., Metallo supramolecular diethylene glycol:water-soluble reversible polymers. Macromol. Rapid Commun. 2002, 23, 957–961
22) Futaki, S., Kiwada, T., Sugiura, Y., Control of peptide structure and recognition by Fe(III)-induced helix destabilization. J. Am. Chem. Soc. 2004, 126, 15762-15769