rights / license: research collection in copyright - non ...23140/... · pathwayfor...
TRANSCRIPT

Research Collection
Doctoral Thesis
Genetische und physiologische Untersuchungen an femD-Mutanten in Methicillin-resistenten Staphylococcus aureus
Author(s): Glanzmann, Philipp Johann
Publication Date: 1999
Permanent Link: https://doi.org/10.3929/ethz-a-003825062
Rights / License: In Copyright - Non-Commercial Use Permitted
This page was generated automatically upon download from the ETH Zurich Research Collection. For moreinformation please consult the Terms of use.
ETH Library

Diss.ETHNr. 13252
Genetische und physiologische
Untersuchungen an/emD-Mutanten
in Methicillin-resistenten
Staphylococcus aureus
ABHANDLUNG
zur Erlangung des Titels
DOKTOR DER NATURWISSENSCHAFTEN
der
EIDGENÖSSISCHEN TECHNISCHEN HOCHSCHULE
ZÜRICH
vorgelegt von
PHILIPP JOHANN GLANZMANN
Dipl. Natw. ETH
geboren am 6. Mai 1969
von Luzern
Angenommen auf Antrag von
Prof. Dr. M. Teuber, Referent
Prof. Dr. B. Berger-Bäehi, Korreferentin
1999

Inhaltsverzeichnis
Abkürzungen iv
Zusammenfassung vi
Abstract viii
1. Einleitung 1
1.1. Staphylococcus aureus l
1.2. Ursprung und Entwicklung der Antibiotikaresistenz 2
1.3. Resistenzmechanismen 3
1.4. Entwicklung der Antibiotikaresistenz bei Staphylokokken 4
1.5. Aufbau der Zellwand und ihre Biosynthese 5
1.6. Die Methicillin-Resistenz: Penicillin-Bindeproteine und Wirkung der ß-Laktame 8
1.7. Die mec Determinante 10
1.8. Die,fem Faktoren (/actors essential for wethicillin resistance) 11
8.1. Das/emAß-System 12
8.2. Das/emC~System 13
8.3. DasfemD-System 14
8.4. DasfemE-System 14
8.5. Das/emF-System 14
1.9. Zusätzliche Methicillin-Resistenz beeinflussende Faktoren 15
1.10. Globale Regulatoren 15
1.11. Zielsetzung der Arbeit 18
2. Material & Methoden 19
2.1. Bakterien und Plasmide 19
2.2. Medien 19
2.3. Chemikalien und molekularbiologische Produkte 25
2.3.1. Synthetische Oligonukleotide 25
2.3.2. Restriktionsendonukleasen und DNA/RNA modifizierende Enzyme 25
2.3.3. Molekulare Grössen und Gewichtsmarker 25
2.4. Allgemeine molekularbiologische Methoden 26
2.5. DNA Isolierungsmethoden 26
2.5.1. Isolierung chromosomaler DNA aus S. aureus 26
2.5.2. Isolierung von Plasmid DNA 27
2.5.3. Isolierung von X Phagen DNA 27
2.5.4. Isolierung von DNA Fragmenten aus Agarose Gel 27
2.6. RNA Isolierungsmethoden 2 8
2.7. Northern-und Southern-Analyse 28

Inhaltsverzeichnis il
2.7.1. Herstellung von DNA Sonden 28
2.7.2. Northern Blot 29
2.7.3. Southern Blot 29
2.7.4. Northern Hybridisierung 30
2.7.5. Southern Hybridisierung 30
2.8. Klonieren von glmM 31
2.9. Sequenzierungen 31
2.9.1. Nicht radioaktive Sequenzierung 31
2.9.2. Radioaktive Sequenzierung 31
2.10. PCR (Polymerase Chain Reaction) 32
2.11. RT-PCR (Reverse Transcriptase Polymerase Chain Reaction) 32
2.12. Primer Extension 32
2.13. Herstellung eines transduzierenden Phagenlysats 34
2.14. Transduktion von S. aureus 34
2.15. Kompetente Zellen 35
2.15.1. Herstellung DMSO-kompetenter E. coli Zellen 35
2.15.2. Herstellung elektrokompetenter DH10B E. coli Zellen 35
2.15.3. Herstellung elektrokompetenter S. aureus RN4220 Zellen 35
2.16. Transformation 36
2.16.1. Transformation von DMSO-kompetenten E. coli Zellen 36
2.16.2. Transformation elektrokompetenter E. coli Zellen 36
2.16.3. Transformation elektrokompetenter S. aureus RN4220 36
2.17. Nachweis der Penicillin-Bindeproteine (PBP) 37
2.18. Western-Blot Analyse 37
2.19. Resistenzbestimmungen 3 8
2.20. Populationsanalysen 38
2.21. Autolyse 38
2.22. Wachstumskurven 39
3. Resultate 40
3.1. Das £/mM-System 40
3.1.1. Phänotypische Charakterisierung der Insertion Q12F glmM::Tn551 40
3.1.2. Das glmM Operon 48
3.1.3. Transkription der glmM Region 60
3.1.4. Komplementation der glmM Mutation 67
3.1.5. Wachstumsverhalten 72
3.1.6. Proteinanalysen 72

Inhaltsverzeichnis m
3.2. DasfmtB-System 78
3.2.1. Phänotypische Charakterisierung der Tn55i Insertion in fintB 80
3.2.2. Komplementation derfintB Mutation 8 3
3.2.3. Wachstumsverhalten 84
3.2.4. Transkription derfmtB Region 84
3.2.5. Proteinanalysen 89
3.3. Das femX-System (fmhB-System) 89
3.3.1. Identifizierung des fem Faktors femX (finhB) 89
3.3.2. Northern Blot 98
4. Diskussion 100
4.1. Die Phosphoglukosamin-Mutase GlmM 100
4.1.1. Sequenz des Phosphoglukosamin-Mutase Operons 101
4.1.2. Suppressor Mutante PG100 102
4.1.3. Transduktion von glmM::Tx\551 J 05
4.1.4. Komplementation der glmM Mutation 105
4.1.5. Proteine 107
4.2. Das/m^-System 107
4.2.1. Transduktion und Wirkung 107
4.2.2. FmtB Protein 109
4.2.3. Offene Fragen über den Effekt derfintB Inaktivation 109
4.3. Das/emX-System (/mfti?-System) 112
5. Literaturverzeichnis 113
6. Anhang 130
Lebenslauf 130
Publikationen 13 ]
Danksagung 132

Abkürzungen IV
Abkürzungen
A Adenin
Amp Ampicillin
dATP Deoxyadenosintriphosphat
bp Basenpaare
bzw. beziehungsweise
C Cytosin
°c Grad Celsius
Cm Chloramphenicol
cm Zentimeter
cps counts per second
dCTP Deoxycytosmtriphosphat
DEPC Diethyl Pyrocarbonat
DNA Desoxyribonukleinsäure
Em Erythromycin
EtOH Ethanol
G Guanin
GlcNAc-1-P N-Acetylglukosamin-1 -Phosphat
h Stunde
kb Kilobasen
kcal Kilokalorien
LB Luria-Bertani
kDa kilo-Dalton
M Molar
Mcr Methicillin-resistent
Mcs Methicillin-empfindlich
min Minuten
mJ Millijoule
raM Millimolar
NAG N-Acetylglukosamin

Abkürzungen
NAM
NaOAc
nm
OD
Ox
ORFs
PCR
RNA
RT
RT-PCR
T
Tet
UDP
UV
UN
w/v
N-Acetylmuraminsäure
Natriumacetat
Nanometer
optische Dichte
Oxacillin
Open Reading Frames
Polymerasen-Kettenreaktion
Ribonukleinsäure
Raum Temperatur
Reverse Transkriptase Polymerase Kettenreaktion
Thymin
Tetracyclin
U
Ultraviolettes Licht
über Nacht
Gewicht pro Volumen

Zusammenfassung vi
Zusammenfassung
Die Methicillin-Resistenz in Staphylococcus aureus wird durch ein
zusätzliches Penicillin-bindendes Protein, das PBP2' (PBP2a), vermittelt.
Dieses PBP2' übernimmt die Transpeptidase-Funktion der anderen vier PBP,
wenn diese durch hohe ß-Laktam Konzentrationen inaktiviert werden. Dieses
zusätzlich PBP2' wird nur in Methicillin-resistenten S. aureus Stämmen
(MRSA) gefunden und wird durch das Strukturgen mecA kodiert, das sich auf
der mec Determinante befindet. Eine charakteristische Eigenschaft der mec-
bedingten Resistenz ist deren heterogene Expression, d.h. die meisten Zellen
einer MRSA Population exprimieren eine niedrige Basisresistenz, während
einige wenige Zellen hochresistent sind.
Neben PBP2' gibt es eine grosse Zahl chromosomaler Faktoren, die die
Methicillin-Resistenz beeinflussen. Diese Faktoren wurden mittels Transposon-
Insertionsinaktivierung mit Tn55I identifiziert und werden fem Faktoren
(factors essential for methicillin resistance) genannt. Im Gegensatz zur mec
Determinante kommen die fem Faktoren auch in Methicillin-empfindlichen S.
aureus Stämmen (MSSA) vor. Diese Faktoren sind direkt oder indirekt in den
Peptidoglykan-Metabolismus involviert und ihre Inaktivierung reduziert die
Methicillin-Resistenz. Einer dieser Faktoren ist das glmM, welches für eine
Phosphoglukosamin Mutase GlmM kodiert und ursprünglich als femD
bezeichnet wurde. Seine Inaktivierung führt zu einer generell erhöhten
Empfindlichkeit gegenüber ß-Laktamen, unabhängig davon ob es sich um einen
MRSA oder einen MSSA handelt. Weiter resultierte die glmM Inaktivierung in
einer Teicoplanin-Hyperempfindlichkeit. Trotz dieser erhöhten
Empfindlichkeit gegen ß-Laktame und Teicoplanin wurde die PBP2'
Produktion nicht beeinflusst. glmM ist Teil eines dreizistronischen Operons
orfJ-orß~glmM. Distal von orfl, als auch vor glmM konnte ein
Transkriptionsstart identifiziert werden, was zu einem dreizistronischen und
einem monozistronischen Transkript führte.

Zusammenfassung vu
Komplementation der g/mM-Mutation mit dem ganzen g/mM-Operon
stellte sowohl die Methicillin-Resistenz als auch die ursprüngliche Teicoplanin-
Empfindlichkeit wieder her. Wurde hingegen nur mit glmM komplementiert,
so konnte zwar die Methicillin-Resistenz wieder hergestellt werden, die
Teicoplanin-Hyperempfindlichkeit blieb jedoch bestehen.
Aus g/mM-Mutanten konnten hoch Methicillin-resistente Suppressor»
Mutanten durch Wachstum auf Methicillin selektioniert werden. Eine dieser
Suppressor Mutanten wurde genauer charakterisiert. Sie produzierte wie die
MethiciUin-empfindlich gewordenen g/mM-Mutanten keine
Phosphoglukosamin-Mutase mehr und war ebenfalls Teicoplanin
hyperempfindlich. Die Suppressor-Mutation war nicht mit der g/mM-Mutation
transduzierbar, korrelierte mit einer erniedrigten spontanen Autolyse und
zeigte eine erhöhte Produktion eines 49 kDa grossen Proteins. Daraus wurde
geschlossen, dass es in S. aureus einen alternativen Weg für die Glukosamin-1-
Phosphat Synthese geben muss.

Abstract vin
Abstract
Methicillin resistance in Staphylococcus aureus is due to the additional
penicillin-binding protein, PBP2' (PBP2a), which has a lower affinity for
methicillin than the endogenous four PBPs. PBP2' is encoded by the structural
gene mecA, has in vitro transpeptidase activity and is thought to substitute for
the functions of the PBPs when they are inactivated by high concentrations of
ß-lactams. Methicillin resistance levels are strain-specific and may vary from
very low to high values. Characteristic for methicillin-resistant S. aureus is the
heterogeneous expression of the resistance with the production of a
subpopulation of cells highly resistant to methicillin.
Besides mecA, there is a large number of chromosomal genes, initially
called fern factors, which are known to influence methicillin resistance levels.
These genes are involved, directly or indirectly, in peptidoglycan metabolism.
Any mutations that alter peptidoglycan precursor composition and/or -
formation reduce methicillin resistance. The phosphoglucosamine mutase
GlmM, initially identified as femD mutation catalyses the conversion of
glucosamine-6-phosphate to glucosamine-1-phosphate, which is an early
cytoplasmatic step in peptidoglycan biosynthesis.
glmM was shown to be the last gene of a three-cistronic Operon orfl-
orß-glmM, One transcriptional start was identified upstream of orf], and a
second transcriptional start producing a mono-cistronic transcript, upstream of
glmM. Disruption of glmM abolished GlmM production, decreased methicillin
resistance, and resulted in teicoplanin hypersusceptibility, without affecting the
production of the endogenous penicillin-binding proteins and PBP2'.
Complementation of the glmM mutation by the complete glmM Operon
restored both methicillin resistance and normal teicoplanin susceptibility. In
contrast, complementation with glmM only restored the methicillin resistance,
but the teicoplanin susceptibility remained low.
A highly methicillin-resistant suppressor mutant obtained by selection for
growth in presence of methicillin remained GlmM deficient and teicoplanin

Abstract ix
hypersusceptible. The inactivation of glmM can be overcome by a suppressor
mutation leading to high level methicillin resistance. The suppressor mutation
was not linked to the glmM Operon and correlated with decreased autolysis and
increased production of a 49 kDa protein suggesting that there is an alternative
pathway for glucosamine-1-phosphate synthesis in S. aureus.

Einleitung 1
1. Einleitung
1.1. Staphylococcus aureus
Die Gattung der Staphylokokken wird in 30 Spezies und Subspezies
unterteilt. Staphylokokken sind grampositiv, unbeweglich, katalasepositiv,
fakultativ anaerob und können in einem bis zu 10% Kochsalz enthaltenden
Medium bei Temperaturen von 18 bis 42 °C wachsen. Sie sind kugelförmig mit
einem Durchmesser von 1 (im und ordnen sich in Haufen bzw. Trauben
(griechisch: staphylé) an, was auf die in verschiedenen Ebenen stattfindende
Zellteilung zurückzuführen ist. Die Pigmentierung ist variabel, sie reicht von
weiss bis goldgelb. Bei diesen Pigmenten handelt es sich um Karotinoide,
welche die Staphylokokken vor Licht und UV-Strahlen schützen (118).
Die grösste pathogène und klinisch bakteriologische Bedeutung kommt
der koagulasepositiven Spezies Staphylococcus aureus zu. Für den gesunden
Menschen stellt S. aureus, der bei ungefähr 30% der Bevölkerung
regelmässiger Bestandteil der Normalflora von Haut und Schleimhäuten ist,
keine Gefahr dar. Bei hospitalisierten und immunsupprimierten Patienten kann
S. aureus jedoch schwere Infektionen hervorrufen. Er gehört zusammen mit
Escherichia coli zu den häufigsten Erregern bakterieller Infekte beim
Menschen. Seine Pathogenität beruht auf einer Vielzahl von Virulenzfaktoren.
Dazu gehören Faktoren wie zum Beispiel die Fibrinogen-bindenden Proteine A
und B, welche es dem Bakterium ermöglichen an extrazelluläre Matrix zu
binden, oder die V8 Protease, die IgG schneidet und inaktiviert oder das
Protein A, welches die Opsonisierung durch Phagozyten behindert, oder die
verschiedenen Hämolysine, die die Invasion von Zellen erleichtern (91).
Die Krankheitsbilder reichen von kutanen Infektionen (Impetigo,
Furunkel), postoperativen und posttraumatischen Wundinfektionen bis hin zu
lebensgefährlichen, systemischen Krankheiten wie Sepsis, Endokarditis,
Pneumonie, toxisches Schocksyndrom oder Lebensmittelvergiftung (16).

Einleitung 2
1.2. Ursprung und Entwicklung der Antibiotikaresistenz
Substanzen, die für eine Behandlung von Infektionen zum Einsatz
kommen, fasst man unter dem Begriff Antiinfektiva zusammen. Neben den
synthetischen Chemotherapeutika gehören Antibiotika zu den wichtigsten und
am meisten verwendeten Antiinfektiva. Bei den Antibiotika handelt es sich um
Naturstoffe, die von Bakterien oder Pilzen produziert werden, die diese zur
Verteidigung ihres natürlichen Lebensraumes gegen andere Mikroorganismen,
die im gleichen Biotop leben, brauchen. Semisynthetische Antibiotika mit
einem erweiterten oder verändertem Wirkungsspektrum werden durch
chemische Modifikationen der Antibiotika erhalten.
Zahlreiche Antibiotika-Resistenzgene waren schon vor dem Einsatz von
Antibiotika in der Medizin als Teile des Antibiotika-Syntheseweges in
Antibiotikaproduzenten vorhanden oder sind in Bakterien entstanden, die mit
Antibiotikaproduzenten eng zusammenleben. Das bedeutet, dass viele
Resistenzdeterminanten bereits vor Millionen von Jahren in Mikroorganismen
entstanden sind.
Neben den in den Antibiotikaproduzenten vorkommenden natürlichen
Resistenzdeterminanten können auch Gene von empfindlichen
Mikroorganismen durch Mutation und Rekombination zu Resistenz führen.
Die horizontale Ausbreitung von Resistenzdeterminanten zwischen
Mikroorganismen erfolgt durch interzellulären Gentransfer mittels
Transformation, Transduktion oder Konjugation (52).
Unter Antibiotikadruck weisen resistente Varianten gegenüber
empfindlichen einen Vorteil auf, werden selektioniert und nehmen schliesslich
deren Platz ein. Der Einsatz von Antibiotika führt also einerseits zu einer
Selektion und Verbreitung von Resistenzgenen, die bereits existierten, und
andererseits zur Evolution neuer Resistenzen (16).

Einleitung 3
1.3. Resistenzmechanismen
Um sich der Wirkung der Antibiotika zu entziehen haben Bakterien
faszinierende Strategien erworben. Damit ein Antibiotikum überhaupt seine
Wirkung entfalten kann, muss es an den Wirkungsort gelangen. Um ins
Zellinnere zu gelangen kann das Antibiotikum die vom Bakterium zur
Verfügung gestellten Transportsysteme benutzen. Resistenz kann sowohl durch
eine veränderte Permeabilität der Zellmembran erworben werden, die das
Antibiotikum nur bedingt oder überhaupt nicht ins Zellinnere gelangen lässt,
als auch durch eine Erhöhung des Exports des Antibiotikums. Dabei werden
Antibiotika mittels einer in der Zytoplasmamembran liegender Effluxpumpe
aktiv von innen nach aussen gepumpt, wie zum Beispiel durch NorA, eine
Effluxpumpe für Norfloxacin und andere Chinolone in S. aureus (80).
Andere Resistenzmechanismen basieren auf inaktivierenden Enzymen,
die das Antibiotikum entweder abbauen oder modifizeren. Zu den abbauenden
Enzymen gehören unter anderem die ß-Laktamasen, welche den ß-Laktamring
der Penicilline hydrolysieren. Zu den modifizierenden Enzymen gehören
sowohl Chloramphenicol-Acetyltransferasen, die die freien Hydroxylgruppen
von Chloramphenicol acetylieren, als auch Aminoglykosid-modifizierende
Enzyme, welche Aminoglykoside entweder durch Phosphorylierung, oder
NukleotidyHerung freier Hydroxylgruppen oder durch Acetylierung freier
Aminogruppen wirkungslos machen (52).
Eine weitere Strategie, derer sich die Bakterien bedienen ist die
Veränderung des Zielmoleküls. Durch Mutation in einzelnen Genen werden
Proteine mit einer erniedrigten Affinität zu Antibiotika gebildet, wie zum
Beispiel Mutationen in der Topoisomerase und Gyrase, die zu Chinolon-
Resistenz oder Mutationen in der DNA-Polymerase, die zu Rifampicin-
Resistenz führen. Oder die Bakterien erwerben ein neues Gen, das für ein
Protein mit erniedrigter Affinität kodiert, wie zum Beispiel das mecA-Gen, das
für ein Penicillin-Bindeprotein (PBP) mit geringerer Affinität zu ß-Laktamen
als die zelleigenen PBPs kodiert. Zur gleichen Strategie kann auch die

Einleitung 4
Vancomycin-Resistenz (2) gezählt werden, die für Enzyme kodiert, die sowohl
den Abbau des empfindlichen Zielmoleküls, wie auch die Synthese eines neuen
resistenten Moleküls katalysieren.
1.4. Entwicklung der Antibiotikaresistenz bei Staphylokokken
Die antibakterielle Wirkung des Penicillins wurde vom englischen
Mikrobiologen Alexander Fleming im Jahre 1928 entdeckt (31). In den frühen
40er Jahren wurde Penicillin in den klinischen Gebrauch eingeführt. Damit
verbesserten sich zwar die Überlebenschancen eines mit Staphylokokken
infizierten Patienten, aber kurz nach Einführen des Therapeutikums konnten
schon die ersten Penicillin-resistenten klinischen S. aureus Isolate identifiziert
werden (4, 83). Ihre Resistenz beruhte auf der Produktion einer
plasmidkodierten ß-Laktamase, die sich horizontal mittels Bakteriophagen
rasch verbreitete. Heute sind weltweit 50-80% der klinischen S. aureus Isolate
gegen Penicillin resistent (69).
Parallel zur Weiterentwicklung der Antibiotika nahmen auch die
Resistenzen kontinuierlich zu. Kaum war Methicillin, das erste
semisynthetische ß-Laktamase-feste Penicillin, im Jahre 1959 in den klinischen
Gebrauch eingeführt worden, konnten kurze Zeit später auch schon die ersten
Methicillin-resistenten S. aureus Stämme (MRSA) identifiziert werden (5, 47).
Ursprünglich war ein einzelner Klon für die ersten weltweiten Epidemien von
MRSA verantwortlich (64). Inzwischen scheint sich die Methicillin-Resistenz in
verschiedenen weiteren klonalen Linien von S. aureus integriert zu haben (1,
77). Mittlerweile wurden in S. aureus an die 40 weitere Resistenzmechanismen
gegen praktisch alle klinisch relevanten Antibiotika identifiziert (102).
Die MRSA Stämme haben ein DNA-Element von 30-50 kb erworben,
das in eine spezifische Stelle ins Chromosom integriert und die Methicillin-
Resistenz-Determinante mec trägt. Diese vermittelt eine inhärente Resistenz
gegen alle ß-Laktame (11), einschliesslich gegen Cephalosporine und

Einleitung 5
Carbapeneme. Zudem begünstigt das DNA-Element die Integration von
weiteren, nicht verwandten Resistenzplasmiden und Transposons in das DNA-
Element, was zu Multiresistenz führt.
Multiresistente S. aureus Stämme haben in den letzten Jahren weltweit
stark zugenommen (19) und Vancomycin ist häufig das einzige, therapeutisch
noch wirksame Antibiotikum. Kürzlich wurden aber bereits die ersten
klinischen Isolate mit reduzierter Empfindlichkeit gegenüber Vancomycin
gefunden (42). Experimentell konnten ebenfalls durch schrittweise Erhöhung
der Vancomycin-Konzentration resistente Mutanten ausgehend von
empfindlichen Stämmen selektioniert werden (100). Es konnte sogar gezeigt
werden, dass unter Laborbedingungen Vancomycin-Resistenz (vanA) von
Enterokokken auf Staphylokokken übertragbar ist (82). Es scheint also nur
eine Frage der Zeit, bis auch Vancomycin bei einer S. aureus Infektion
wirkungslos sein wird. Daher ist es sehr wichtig, nach neuen Wegen und
Mitteln zur Bekämpfung von Infektionskrankheiten zu suchen, wenn verhindert
werden soll, dass die moderne Medizin in die präantibiotische Zeit zurückfällt.
1.5. Aufbau der Zellwand und ihre Biosynthese
Die Zellwand von S. aureus ist ein halb-rigides, dynamisches
Makromolekül, das sich aus Peptidoglykan, auch Murein genannt, und kovalent
gebundenen Teichonsäuren und Proteinen zusammensetzt. Die Mureinschicht
kann bis zu 40 Peptidoglykanstränge dick sein und 50% des Zellwandgewichtes
betragen (35). Das Peptidoglykan besteht aus unverzweigten Glykansträngen,
die über kurze Peptide miteinander vernetzt sind (96). Aufgebaut sind diese
Glykanstränge aus alternierend ß-1,4 glykosidisch verbundenen Aminozuckern
dem N-Acetylglukosamin (NAG) und der N-Acetylmuraminsäure (NAM).
Glykanstränge erreichen im Durchschnitt eine Länge von 10
Disaccharideinheiten (99). Die Peptide, die die Glykanstränge untereinander
vernetzen, bestehen aus dem von der NAM ausgehenden Stammpeptid L~Ala-

Einleitung 6
iD-Gln-L-Lys-D-Ala-D-Ala und der für S. aureus charakteristischen
Pentaglycinkette, die das Peptidoglykan zwischen der e-Aminogruppe des L-
Lys einer Mureineinheit und dem D-Ala der anderen unter Abspaltung des
letzten D-Ala Restes verbindet. Der Quervernetzungsgrad im Peptidteil des
Mureins erreicht bei S. aureus bis zu 90% (62).
Weiter kann das Kohlestoffatom C-6 der NAM mit Teichonsäuren
substituiert sein, die 20-50% der Zellwandmasse ausmachen können, oder mit
O-Acetylgruppen verbunden sein (96).
Die Biosynthese und der Einbau der Bausteine des Peptidoglykans in das
Zellwandgerüst lassen sich in drei lokale Abschnitte unterteilen und zwar in die
Biosyntheseschritte die im Zytoplasma, an der Zytoplasmamembran und in der
Zellwand stattfinden. Ausgehend von Glukosamin-6-Phosphat wird in
aufeinanderfolgenden enzymatischen Schritten UDP-N-Acetyl-Muraminsäure
(UDP-NAM) gebildet, an die schrittweise die fünf Aminosäuren des
Stammpeptids, L-Ala-iD-Gln-L-Lys-D-Ala-D-Ala, synthetisiert werden (Fig.
1). Durch den Austausch von UDP gegen den C55-Lipidcarrier
Undekaprenylphosphat wird der Komplex, das Lipid 1, an die
Zytoplasmamembran gebunden. Dort wird zusammen mit UDP-NAG ein
Disaccharidpentapeptid, das Lipid II, gebildet. In den nächsten Schritten wird
die Pentaglycinseitenkette an das L-Lys des Stammpeptids synthetisiert. Im
dritten Biosyntheseabschnitt wird die membranständige Vorstufe, unter
Freisetzung des Membranankers Undekaprenyldiphosphat, durch die
Zytoplasmamembran nach aussen in den Bereich der Zellwand geschleust. Hier
erfolgt die Verknüpfung der Disaccharide mit dem Peptidoglykangerüst durch
Transglykosylierung und einer anschliessenden Quervernetzung des
Peptidanteils mittels Transpeptidasen. Dabei muss das endständige D-Ala
abgespalten werden, damit die freie Carboxylgruppe des anderen D-Ala mit
der Aminogruppe des endständigen Glycins der Pentaglycinkette verknüpft
werden kann (93).

Einleitung 7
Fruktose-6-P
NagB lÀGImSH<
Glukosamin-6-P
1 GlmM
Glukosamin-l-P
acetyl-CoA . GimU
.glul.
:etyl
I4Ac-
'-Mi
N-Acetylglukosamin-1 -P
UTP 1 GimU
UDP-N-Acetylglukosamin
MurZ
UÜP-GIcNAc-Enolpyruvat
MurB
UDP-MurNAc
MurC, D. E. F
Pentapeptid
UDP-MurNAc
C„-P •=> è MraY
Pentapeptid
C„-PP-MurNAc
UDP-GlcNAc => IMurG
Lipidl
IPentapeptid
C„-PP-MurNAe Lipidil
GlcNAc
I(Gh ),=t> w FmhBi FemAj FemB
Pentapeptid~~(Gly)s
C,,-PP-MurNAc
GlcNAc
C S-PP ^""X Transpeptidase. Fransglykosylase
Peptidoglykan
Figur 1: Zellwandhiosynthese in S. aureus, modifiziert naeh Höltje (44). NagB: Deaminase; GlmS:
Amidotraiisleiase; GlmM: Phosphoglukosamin-Mutase; GimU. bifunktionelles Enzym, Acetyltransferase
und UDP-Pyrophosphorylase; MmZ: Enolpyru\\l Transferase; MurB: Reduktase; MurCDEF:
Synthetasen, die die einzelnen AS übertragen ; MraY: Transferase; MurG: Transferase; FmhB, FemAB:
Bildung der Pentaglycinseitenkette.

Einleitung 8
1.6. Die Methicillin-Resistenz:
Penicillin-Bindeproteine und Wirkung der ß-Laktame
ß-Laktame und ihre Derivate bilden eine grosse Gruppe von sicheren
und effizienten Antibiotika. Sie sind gegen die letzten Schritte der
Zellwandbiosynthese gerichtet. Zu den ß-Laktamen gehört auch das
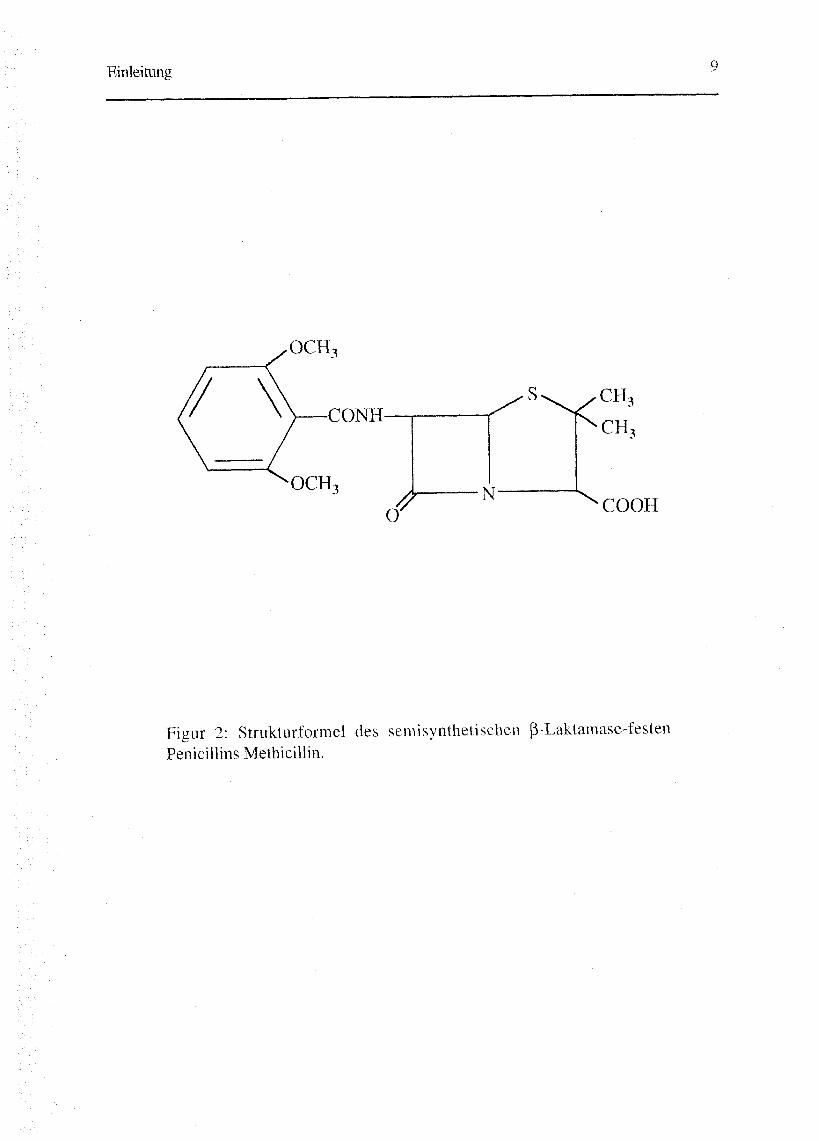
semisynthetische Methicillin (Fig. 2).
Der primäre Wirkungsort der ß-Laktame sind die sogenannten
Penicillin-Bindeproteine (PBP), die die letzten Schritte der Mureinbiosynthese,
die Transpeptidase Reaktion, katalysieren. PBP werden in allen Bakterien
nachgewiesen, die einen Mureinsacculus synthetisieren.
Alle Methicillin-empfindlichen S. aureus Stämme (MSSA) verfügen über
drei hochmolekulare PBP (PBP1-3) und ein niedermolekulares PBP (PBP4).
Im Gegensatz zu den bifunktionellen PBP von E. coli, die sowohl
Transpeptidase- als auch Transglykosylase-Aktivität besitzen, katalysieren die
PBP von S. aureus nur Transpeptidasereaktionen, während die
Transglykosylase-Aktivität auf einem separaten, nicht Penicillin-sensitiven
Enzym vorhanden ist (86). Die genaue physiologische Funktion der
Staphylokokken-PBP ist bis heute nicht geklärt, jedoch zeigt sich eine
unterschiedliche Bedeutung für die Zelle. Durch selektive Inaktivierung
einzelner PBP konnte nachgewiesen werden, dass PBP1 die Schlüsselrolle
zufällt. Hemmung von PBP1 führt zu einem Verlust des „Splitting Systems",
das an der Trennung der Tochterzellen beteiligt ist (8). Zudem ist PBP1
essentiell für das Zellwachstum (112). PBP2 wirkt als Transpeptidase und
katalysiert die Quervernetzimg von neu synthetisiertem Peptidoglykan mit der
bereits existierenden Zellwand, während PBP3 in die Septum-Bildung
involviert zu sein scheint (92). PBP4 schliesslich katalysiert eine sekundäre
Transpeptidase-Reaktion, die in die Quervernetzung des Peptidoglykans
involviert ist.
Die Wirkung der ß-Laktame ist stark konzentrationsabhängig und daher
ziemlich komplex. Niedere Konzentrationen hemmen das Zellwachstum,
Einleitung9
OCH3
CONH
COOH
Figur 2: Strukturformel des semisynthetischen ß-Laktamase-festenPenicillins Methicillin.

Einleitung 10
während höhere Konzentrationen, die der Minimalen Wachstums-Hemm-
Konzentration (MHK) entsprechen, zu einer Abnahme der Quervernetzung der
Zellwand und zu einer Hemmung der Bildung eines neuen Septums führen.
Zelltod wird bei solchen Konzentrationen durch Murosomen-induzierte
Perforation der Zellwand verursacht (63). Diese lokalisierte Perforation der
Zellwand wird am Ort des neuen Septums durch eine N-Acetylmuramyl-L-
Alanin-Amidase und eine endo-ß-N-Acetylglukosaminidase bewerkstelligt, die
durch das atl-Gen kodiert werden (107). Bei sehr hohen ß-Laktam-
Konzentrationen erleiden die Bakterien einen Tod, der nicht von schneller
Lyse begleitet wird (113).
1.7. Die mec Determinante
Methicillin war das erste semisynthetische ß-Laktamase-feste Penicillin,
das im lahre 1959 in den klinischen Gebrauch eingeführt wurde. Kurze Zeit
später konnten jedoch bereits die ersten Methicillin-resistenten S. aureus
Stämme (MRSA) identifiziert werden (5, 47).
Die intrinsische Methicillin-Resistenz bei MRSA-Stämmen ist durch die
Anwesenheit einer chromosomal lokalisierten mec Determinante bedingt. Sie
liegt auf einem zusätzlichen, mindestens 30 kb grossen Stück DNA (7), das in
Methicillin-empfindlichen S. aureus (MSSA) Stämmen nicht vorhanden ist
(104). Diese Determinante enthält neben den regulatorischen Elementen med
und mecRI das Strukturgen mecA, das für ein zusätzliches, hochmolekulares
PBP, genannt PBP2' oder PBP2a, kodiert (40). Dieses zusätzliche PBP wird in
allen MRSA-Stämmen gefunden und besitzt eine stark reduzierte Affinität zu
ß-Laktamen (18). In Anwesenheit hoher ß-Laktam-Konzentrationen
übernimmt PBP2' offenbar die Aufgabe der anderen PBP (62) und
funktioniert möglicherweise als Transpeptidase (34). Die Transkription von
mecA steht sowohl unter der Kontrolle seines eigenen Regulationssystems
mecRI-mecI als auch unter der des ß-Laktamase-Regulationssystems blaRI-blal

Einleitung 11
(94). med kodiert für den Repressor (97), während mecRI für den
transmembranen ß-Laktam Sensor-Transduktor kodiert, der aus einer ß~
Laktam erkennenden Domäne ausserhalb der bakteriellen Membran und einem
ins Zytoplasma hereinragenden Zink-Peptidase Motiv besteht (50). Vermutlich
aufgrund der Selektion durch ß-Laktame besitzen viele klinische MRSA Isolate
Punktmutationen oder Deletionen im med-Gen oder im mecA Operator, so
dass der Repressor nicht mehr an seine Zielsequenz binden kann. Als Folge
wird PBP2' konstitutiv synthetisiert (53, 108). Die Inaktivierung des
Repressors führt zwar in gewissen Stämmen aufgrund der Zunahme sowohl
der mecA Transkription als auch der PBP2' Produktion zu einem Anstieg der
Methicillin-Resistenz, doch scheint ein vollständiges mecI-mecRI und eine
intakte Operatorsequenz in anderen, genetisch unterschiedlichen Stämmen nur
einen geringen Effekt auf die mecA Transkription und die PBP2' Produktion
zu haben (81). Offenbar gibt es stammspezifische Unterschiede und es scheinen
einzelne Schritte im Regulationsnetzwerk der PBP2' Induktion noch nicht
bekannt zu sein.
Eine charakteristische Eigenschaft der raec-bedingten Resistenz der
MRSA-Stämme ist deren heterogene Expression (29). Die meisten Zellen einer
MRSA Population exprimieren eine niedrige Basisresistenz gegen Methicillin,
während einige wenige Zellen (eine in 104 bis 107) hochresistent sind. Die Höhe
der Resistenz und die Grösse der hochresistenten Subpopulation ist eine
stammspezifische, konstante Eigenschaft (51). Neben dem genetischen
Hintergrund eines jeden Stammes beeinflussen auch äussere Faktoren wie
Temperatur, pH, Osmolarität, Medium, Anaerobiose und Licht die Höhe der
Resistenz (73).
1.8. Ditfem Faktoren (/actors essential for methicillin resistance)
Aufgrund verschiedener Beobachtungen, dass die in der Zelle
vorhandene Menge PBP2' nicht direkt mit der Höhe der Methicillin-Resistenz

Einleitung 12
korreliert (20, 40, 76), wurden zusätzliche Faktoren für die Expression der
Methicillin-Resistenz verantwortlich gemacht (13). Diese Faktoren, die mittels
Transposon-Insertionsinaktivierung mit Tn55i identifiziert wurden, liegen auf
dem Chromosom, sind aber nicht mit der mec Determinante assoziiert (14, 24,
111). Sie werden fem (factors essential for methicillin resistance) (12) oder
aux (auxiliäre) Faktoren (109) genannt und kommen sowohl in MRSA- als
auch in MSSA-Stämmen vor. Die meisten dieser fem Faktoren beeinflussen
direkt oder indirekt den Zellwandmetabolismus (15).
1.8.1. Das femAB-System
Von allen bekannten fem Faktoren hat das femAB Operon den grössten
Einfluss auf die Methicillin-Resistenz. Seine Inaktivierung führt zu einer
Hyperempfindlichkeit gegenüber allen ß-Laktamen. Das femAB Operon liegt
auf dem Smoi Fragment A und entstand vermutlich aufgrund einer
Sequenzduplikation. Es kodiert für zwei in ihrer Aminosäurensequenz sehr
ähnliche Proteine von ungefähr 50 kDa, die 39% Identität und 70%
Ähnlichkeit aufweisen (12). Die Proteine FemA und FemB sind an der Bildung
der charakteristischen Pentaglycinseitenkette des Mureins beteiligt, wobei
FemA für das Anhängen der Glycine Gly2 - Gly3 (70) und FemB für die
Glycine Gly4 - Gly5 (41) verantwortlich ist. Trotz ihrer Sequenzähnlichkeiten
können sich FemA und FemB nicht gegenseitig komplementieren (27).
femA und femB Mutanten produzieren nur noch Mono- respektive
Triglycinseitenketten, wobei die femA Mutation zu einer Resistenz gegenüber
der Glycyl-Glycin Endopeptidase Lysostaphin führt. Dieser reduzierte
Glycingehalt hat einerseits eine erniedrigte Quervernetzung des Mureins und
andererseits verminderte Autolysinaktivität zur Folge (70). Weiter zeigen diese
Stämme eine gestörte Septumanordnung, eine verlangsamte Zelltrennung und
ein pseudomultizelluläres Erscheinungsbild (41). Der wichtigste Effekt der
Inaktivierung ist jedoch die Reduktion der Resistenz gegen Methicillin und
allen anderen ß-Laktamen bei MRSA-Stämmen, obwohl diese Stämme

Einleitung 13
weiterhin PBP2' exprimieren (10). Zudem zeigen femAB Mutanten auch
grössere Empfindlichkeit gegen ein ganzes Set anderer Antibiotika (67).
Von den Resultaten der kompletten Inaktivierung des femAB Operons,
die nur noch zu Monoglycinseitenketten führt, lässt sich neben FemA und
FemB ein zusätzlicher Faktor postulieren (105). Dieser dritte Faktor, vorläufig
FemX genannt, soll für den Einbau des Gly, verantwortlich sein (57). Da aber
eine femAB Mutation das Wachstum einer Zelle bereits sehr stark reduziert,
kann davon ausgegangen werden, dass eine Inaktivierung des postulierten
Faktors FemX wahrscheinlich letal ist.
Der charakteristische Aufbau der Pentaglycinseitenkette mit Hilfe der
Fem Faktoren spricht für eine räumlich kontrollierte Anordnung der
Reihenfolge dieser Faktoren FemX-FemA-FemB, ähnlich einem streng
geordneten Multienzymkomplex. Interessanterweise sind in S. aureus vier
verschiedene Glycyl-t-RNA gefunden worden. Eine dient der
Proteinbiosynthese, die drei weiteren der Mureinbiosynthese (37), die
zahlenmässig den drei Fem Faktoren X, A und B entsprechen.
1.8.2. Das femC-System
femC befindet sich ebenfalls auf dem chromosomalen Smal Fragment A,
ist jedoch nicht mit dem femAB Operon assoziiert (14), sondern ist mit thrB
kotransduzierbar (38). Durch eine Tn55i Insertion bedingte Inaktivierung von
glnR im Glutaminsynthetase (glnRA) Operon wird in femC Mutanten die
Transkription vom distalen glnA Gen, das für die Glutaminsynthetase kodiert,
herabgesetzt, was zu einem Mangel an Glutamin führt (38). Im weiteren zeigt
sich in der Mutante eine deutliche Reduktion des Amidierungsgrades der iso-D-
Glutaminsäure (iD-Glu) im Stammpeptid und eine geringere Quervernetzung
des Mureins (84, 93). Die Inaktivierung hat aber nur einen schwachen Effekt
auf die Ausprägung der Methicillin-Resistenz. Dabei ist nur die Grundresistenz
dieser Mutanten reduziert, die Resistenzhöhe der hochresistenten Subpopulation
bleibt annähernd erhalten (14). Die Methicillin-Resistenz (38) und die

Einleitung 14
Amidierung von iD-Glu (93) kann durch externe Zugabe von Glutamin ins
Medium wiederhergestellt werden.
1.8.3. Das femD-System
femD wurde mittels Tn55i Insertion identifiziert und liegt auf dem
chromosomalen Smal Fragment I (14). Die Inaktivierung von femD hat einen
ähnlich schwachen Effekt auf die Ausprägung der Methicillin-Resistenz wie die
Inaktivierung von femC, d.h. es wird ebenfalls nur die Grundresistenz
reduziert, während die Resistenzhöhe der hochresistenten Subpopulation
annähernd erhalten bleibt (14, 36). Neben einem verringerten Anteil von
Alanin anstelle von Glycin in der ersten Position der Seitenkette kann keine
weitere, signifikante Zellwandveränderung festgestellt werden (93).
Interessanterweise liegt femD, gleichbedeutend wie glmM (116), genau
auf dem chromosomalen Smal Fragment auf welchem Mutationen, die zu einer
erhöhten Resistenz gegen Teicoplanin führen (98), lokalisiert wurden.
1.8.4. Das femE-System
Bis heute ist sehr wenig über femE bekannt. Es befindet sich wie das
femAB Operon und femC auf dem chromosomalen Smal Fragment A. Die
Methicillin-Resistenz ist nur geringfügig herabgesetzt. Es kann auch keine
signifikante Zellwandveränderung in entsprechenden Mutanten festgestellt
werden (24).
1.8.5. Das femF-System
femF Mutanten haben eine beeinträchtigte Zellwandbiosynthese und zwar
kann in diesen Mutanten kein L-Lysin mehr an das Stammpeptid des Mureins
gehängt werden. Die Folge ist eine Akkumulation von Dipeptid-Peptidoglykan-

Einleitung 15
Vorstufen in der Zellwand (85). Das Gen ist auf dem Smal Fragment B
lokalisiert und wurde noch nicht eingehender untersucht.
1.9. Zusätzliche Methicillin-Resistenz beeinflussende Faktoren
Neben den erwähnten fem Faktoren gibt es noch weitere chromosomale
Faktoren, die die Methicillin-Resistenz beeinflussen.
Das Gen fmt kodiert für ein PBP-ähnliches Protein (54), das
möglicherweise in die Zellwandbiosynthese involviert ist oder als
Signaltransduktor fungiert.
Ein weiters Gen, das mittels Insertion inaktiviert wurde und dabei die
Methicillin-Resistenz reduziert, ist das //m-Gen, das für ein 38 kDa schweres
membrangebundenes Protein kodiert (71).
lytH war das erste Gen, bei dem gezeigt werden konnte, dass zwischen
der lytischen Aktivität und der Methicillin-Resistenz ein Zusammenhang
besteht (32). Inaktivierung von lytH führt zu einer hohen homogenen
Methicillin-Resistenz.
1.10. Globale Regulatoren
In S. aureus gibt es verschiedene chromosomal kodierte globale
Regulatoren, die die Ausprägung der Methicillin-Resistenz beeinflussen können
(Tab. 1). Dazu gehören unter anderem die Operons agr und sar, die beide
einen geringen Einfluss auf die Methicillin-Resistenz haben, wobei allerdings
noch unklar ist auf welche Art und Weise sie die Methicillin-Resistenz
beeinflussen (26). Bei agr handelt es sich um einen globalen Regulator, der die
Regulation der wachstumsabhängigen Exoprotein- und Zellwand-assoziierten
Proteinbiosynthese kontrolliert (88). Der agr-Lokus baut sich aus zwei
divergenten, von zwei verschiedenen Promotoren kontrollierten Operons auf.

Tabelle
1:Be
einf
luss
ungderMethicillinresistenzdurchchromosomaleFaktoren
Gen
Funktion
Referenz
Membran-
oderZellwand-assoziiertechromosomaleGene
fmt
PBP-ähnlichesProtein
Funktionunbekannt
(54)
php2
PBP2,Tr
ansp
eptida
se,Peptidoglykanbiosynthese
(89)
Um
lipophiles,membrangebundenes
Protein
Beei
nflu
ssun
gderau
toly
tisc
henAktivität
(71)
lytH
autolytische
Aktivität
(32)
GlobaleRegulatoren
sar,agr
sigB
Regu
lati
onderwachstumsabhängigenExoprotein-undzellwand-assoziierten
Proteinbiosynthe
se
Expression
vonGenen
inderstationärenWachstumsphaseundunterbestimmten
Stre
ssbe
ding
ungen
(26)
(117)

Tabelle
1:Fo
rtse
tzun
g.
InPe
ptid
oglykanvorläufersynthese
involviertechromosomaleGene
femX
postulierteGlycin
1Se
iten
kett
enbi
ldun
g(5
7)
femA
Glyc
in2-
Glyc
in3Se
iten
kett
enbi
ldun
g(1
2)
femB
Glycin4-Glycin5Se
iten
kett
enbi
ldun
g(4
1)
femC
(glnR)
Glut
amin
synt
hetase
Repressor
AmdierungdesiD-GlutamatdesStammpeptids
(38)
femD(glmM)
Phosphoglukosaminmutase,BildungdesPeptidoglykanvorlä
ufersGlukosamin-
1-Phosphat
(48)
femF
Lysinaddierender
SchrittbeiderStammpeptid-Bildung
(84)

Einleitung 18
Bei sar handelt es sich ebenfalls um einen globalen Regulator, der die
Expression von agr zu beeinflussen scheint (21).
Der Transkriptionsfaktor sigB reagiert auf Stresseinwirkung und die
Stationärphase (60). Es wurde gezeigt, dass eine Tn55i Insertion in sigB in
einem homogen resistenten MRSA die Methicillin-Resistenz herabsetzt (117).
Die Rolle der globalen Regulatoren sowie SigB bezüglich der Methicillin-
Resistenz wird noch genauer untersucht werden müssen.
Da äussere Faktoren wie Temperatur, Osmolarität, Nährmedien und pH
die Methicillin-Resistenz verändern können, muss die Expression der
Methicillin-Resistenz auch von Umweltsensoren abhängen (73).
1.11. Zielsetzung dieser Arbeit
Die Inaktivierung von glmM (-femD) mittels Tn551 erhöht die
Empfindlichkeit von allen S. aureus Stämmen gegenüber ß-Laktamen ob es
sich nun um einen mec Determinante tragenden MRSA, eine in vitro durch
Mutation und Selektion für Wachstum auf Methicillin erhaltene Methicillin-
resistente Mutante, oder einen MSSA handelt (14, 36). Herabgesetzt wird
gleichzeitig auch die Resistenz gegenüber den Glycopeptiden Vancomycin und
Teicoplanin, wobei vor allem die Empfindlichkeit gegenüber Teicoplanin
zunimmt (36).
Ausgehend von diesen Resultaten wurde das femD Gen kloniert,
sequenziert und charakterisiert.

Material & Methoden 19
2. Material & Methoden
2.1. Bakterien und Plasmide
Die Staphylococcus aureus- und E. co/i-Stämme sowie die Vektoren und
die rekombinanten Plasmide können den Tabellen 2, 3 und 4 entnommen
werden.
2.2. Medien
LB-Bouillon für Flüssigkulturen
LB-Agar für Platten
LB Softagar für Transduktionen
SOC-Medium
Müller-Hinton-Agar-Platten
20 g/1 LB Broth (Difco, Detroit, USA)
20 g/1 LB Broth
1.5% w/v Agar (Difco, Detroit, USA)
20 g/1 LB Broth
0.6% w/v Agar
20 g/1 Trypton (Difco, Detroit, USA)
5 g/1 Yeast Extract (Difco, Detroit, USA)
2.5 mM KCl
lOmMNaCl
lOmMMgCl,
10mMMgSO4
20 mM Glucose
21 g/1 Müller-Hinton Broth
(Difco, Detroit, USA)

Tabelle
2:Staphylococcus
aureusStämme
S.aureus
relevanterGenotyp
relevanterPh
änot
ypHerkunft,
Stämme
Bemerkungen
BB255
8325
ue
(13)
BB270
8325mec
Mcr
(13)
BB270AsigB
8325mec,AsigBOp
eron
Mcr,Tef
(61)
PG27
8325,
ClUFglmM::Tn55I
Mcs,Emr,kleine
Kolonien
dieseArbe
it;TransduktionvonQ12FglmM::Tn551von
BB591
inBB270
PG108
PG100
8325mec,
012FglmM::Tn551
8325mec,
Mcs,Em1
Mcr,Emr
dieseArbeit;TransduktionvonQ12FglmM::Tn551von
BB591
inBB270
dieseArbe
it;McrSu
ppre
ssor
MutantevonPG108
PG105
PG106
PG79
012FglmM::Tn551hmrDl
8325mec,
012Fg/mM::Tn55i
8325mec,
D.X2VglmM::ln551
8325mec,
Mcs,Emr
Mcs,Emr
Mcr,Emr,Tef
dieseArbeit;TransduktionvonQ12Fgl
mM::
Tri5
51von
PG100inBB270
dieseArbeit;TransduktionvonQ12Fgl
mM::
Tii5
51von
PG108
inCOL
dieseArbe
it;TransduktionvonPG108
PG231
Q12Fg/mM::TnJ5i;
8325mec,
pPG76
Mcr,Emf,Cm1
dieseArbeit;TransduktionvonPG108
PG232
Q12Fg/mM::Tn55i;
8325mec,
pHK4176
Mcr,Emr,Cm'
dieseArbeitTransduktionvonPG100
PG217
012Fg/mM::Tn557;
pHK4176
8325mec,
Q12Fg/mM::Tn55i;
hmrDl
pAW8
Mcs,Enf,Tef
dieseArbeit;TransduktionvonPG108
TO a.
EL
9p Ci Ero & a too

RN4220
PG77
PG78
BB1000
BBlOOOAs/gß
BB1163
BB1164
BB1165
BB1166
COL
COL-TS111
COL-TS111
pHK4357
COL-TS111
pHK4069
RN450
RN450-TS111
KSA8
Ha56
Ha57
8325-r"
8325-f
;pAW8
8325-r
;pPG76
Newman
Newman,AsigBOperon
$325Aa
grv.
tet
8325
Aagr
v.te
t,sar::7n917LTVl
8325mec,Aa
grv.
tet
8325mec,Aagrv.te
t,sar::Tn917LTVl
COL
COLfmtB::Tn551
Restriktion,
transformierbardurch
Elek
trop
orat
ion
Rest
rikt
ion"
;Tef
Rest
rikt
ion"
;Tef
Enf
Mc\Tef
Mcs,Tef,Emr
Mcr,Tef
Mcr,Tef,Em1
Mcr,Oxr
Me',Emr
COLfmtB::Tn551
;pHK4357
Ox\Emr,Cmr
COL
fintB:
:Tn55i
;pHK4069
Oxr,
Emr,Cm'
8325-4
8325-4frntBv.7n551
mec,fintBpartiel]
8325mec,
fmtB::Tn551
8325mec,
fmtB::7n551
Mcs
Mcs,Em'
Oxr,
Mcr
Mcs,Emr
McT,Emr
(59)
dieseArbeit;ElektroporationvonRN4220
dieseArbeit;ElektroporationvonRN4220
erhaltenvon
P.Vaudaux,Genf
(61)
(26)
(26)
(26)
(26)
(58)
COL
muta
genisiertmitTnJJ/
infintB;
erhaltenvon
Komatsuzawa
erhaltenvonKomatsuzawa
erhaltenvonKomatsuzawa
erhaltenvonKomatsuzawa
erhaltenvonKomatsuzawa
KlinischesIsolâtvonKomatsuzawa
dieseArbeit;TransduktionvonfintB::7n551
COL-TSlllinBB270
dieseAr
beit;TransduktionvonfmtB::Tn5'51
COL-TSlllinBB270

BB1395
8325mec,
Mc',Em'
dieseArbeit
;TransduktionvonfintB::7n551
fmtBv.7n551
COL-TSlllinBB270
BB1399
8325mec,
Mcs,Em'
dieseArbeit;TransduktionvonfintB::7n551
fmtBv.7n551
COL-TSlllinBB270
s. Rp
Cm',Chlo
ramphenicol-resistent;
Em',Er
ythromycin-resistent;Mc',Methiciliin-resistent;
Mcs,Methicillin-empfmdlich;Tef,Tetracyclin¬
resistent;
Ox',
Oxacillin-resistent.
o S

Tabelle
3:E.
coliStämmeundVektoren
E.coli
relevanterGenotyp
Herkunft
Stämme
Bemerkungen
DH5oc
supE44,AlacU169,FSOdlacZ,AMI5,hsdR17,
recAl,endAl,gyrA99,
thi-
1,relAl
Gibco,Basel,Schweiz
DH10B
F",mcrA,galKF,
rpsL,
(68),
A(mrr-hsdRMS-mcrBC)FSOdlacZ,AM15,
Stammzum
KlonierengrosserFragmente
AlacX74,deoR,recA,endAl,araD139,
A(ar
a,le
u)76
9,galU,nupG
CD
o 3
Vektor
Grösse
Phänotyp
Herkunft
Bemerkungen
pAW8
5.1kb
Tef
Wada,
unpublizie
rt,E.coli-S.aureus
shuttleVektor
pTZ18R
2.9kb
Ampr
USB
Corp
.,Cleveland,Ohio,USA
pUC18
2.69kb
Ampr
Boeh
ring
erMannheim,Schweiz
pLI50
5.266kb
Ampr,Cmr
(66);E.coli-S.aureus
shuttleVektor
pGC2
5.78kb
Ampr,Cmr
E.coli-S.aureus
shuttleVektor,
erhaltenvon
P.Matthews
Ampr,
Ampicillin-resistent;Cmr,Chloramphenicol-resistent;
Tef,
Tetracyclin-
resi
sten
t.

Tabelle4:
RekombinantePlasmide
rekombinante
Vektor,
Schnittstelle
Insertgrösse,Stamm,
Herkunft
Plasmide
Schn
itts
tell
e,Gene
Bemerkungen
pPG76
pAW8,
Pstl
5kb(BB270)
Pstl
,ar
gl-o
rfl-
orfl-glmM
dieseArbeit
pBBB106
pTZ18R,Xbal
10.5kb(BB591)XM,
JR(ai2Fg/mM;:Tn55/)
Gustafson,un
publ
izie
rt
pPGl.l
pUC18,Smal
4.1kb(BB270)£coRV
femX-orfl
dieseArbeit
pHK4176
pLI50
1.8kbPCR-FragmentmitglmM
erhaltenvonKomatsuzawa
pHK4069
pGC2,Xhal
8.9kb(KSA8)XbalFragment
mitfmtB
erhaltenvonKomatsuzawa
pHK4357
pGC2
7.8kbPCR-Fragment
ausCOL
erhaltenvonKomatsuzawa
Cmr,Chloramphenicol-resistent;
Emr,Er
ythr
omyc
in-r
esis
tent
;Mcr,
Methicil
iin-
resi
sten
t;Mcs,
Methicillin-
empfindlich;
Tef,
Tetr
acyc
lin-resistent;
Oxr,
Oxacillin-resistent;

Material & Methoden 25
2.3. Chemikalien und molekularbiologische Produkte
Die Chemikalien und Lösungsmittel wurden, wenn nicht speziell
angegeben, über die Firmen Fluka (Buchs, Schweiz), Sigma (Buchs, Schweiz)
oder BioRad (Glattbrugg, Schweiz) bestellt. Radiochemikalien wie a35S-dATP,
Y^P-dATP und a32P-dCTP wurden bei Amersham Life Science (Zürich,
Schweiz) bezogen.
2.3.1. Synthetische Oligonucleotide
Synthetische Oligonukleotide wurden durch Microsynth (Balgach,
Schweiz) hergestellt.
2.3.2. Restriktionsendonukleasen und DNA/RNA modifizierende
Enzyme
Restriktionsendonukleasen und DNA/RNA modifizierende Enzyme
wurden (wenn nicht speziell angegeben) bei den Firmen Boehringer Mannheim
(Rotkreuz, Schweiz), MBI Fermentas (Gebrüder Mächler AG, Basel, Schweiz)
oder Gibco Life Technologies (Basel, Schweiz) bezogen. Die AmplitaqGold
DNA Polymerase und die pfu Polymerase wurden bei Perkin Eimer (Rotkreuz,
Schweiz) bestellt. Die Sequenzierungskits wurden von Amersham Life Science
(Zürich, Schweiz) erhalten.
2.3.3. Molekulare Grössen und Gewichtsmarker
Der molekulare Grössenmarker für DNA (1 kb Plus DNA ladder) wurde
bei Gibco (Basel, Schweiz) bezogen. Der molekulare Grössenmarker für RNA
(RNA Molecular Weight Marker I, 0.3-7.4 kb) wurde von Boehringer
Mannheim (Rotkreuz, Schweiz) erhalten. Der vorgefärbte molekulare Protein-
Gewichtsmarker (Prestained SDS/PAGE Standards, low range) und der

Material & Methoden 26
molekulare Protein-Gewichtsmarker (SDS/PAGE Standards, broad range)
wurden bei Bio Rad (Glattbrugg, Schweiz) bestellt.
2.4. Allgemeine molekularbiologische Methoden
Alle grundlegenden molekularbiologischen Methoden wurden den
Protokollen von Ausubel et al. (3) und Maniatis et al. (72) entnommen.
2.5. DNA-Isolierungsmethoden
2.5.1. Isolierung chromosomaler DNA aus S. aureus
Das Pellet einer 30 ml ÜN-Kultur wurde in 3 ml Lysis-Mix (0.1 M Tris-
HCl, pH 7.5; 0.1 M EDTA; 0.15 M NaCl) mit 1 Ltg/ml Lysozym und 0.2 u.g/ml
Lysostaphin suspendiert und für 30 min bei 37 °C inkubiert. Um vollständige
Lyse zu erreichen wurden anschliessend 0.3 ml 5% SDS in 50% EtOH
zugegeben, kurz gevortext und 5 min bei RT inkubiert. Nach Zugabe von
Phenol/Chloroform/Isoamylalkohol (25:24:1) und 10 min Zentrifugation bei
40000g wurde die chromosomale DNA mit zwei Volumen EtOH versetzt, auf
einen Glasstab aufgespult, 2 x in 100% Ethanol gewaschen und in 1 ml TE
Puffer (0.01 M Tris-HCl, pH 8; 1 mM EDTA) über Nacht bei RT gelöst. Um
die RNA abzubauen wurden 0.5 q,g DNase-freie RNase dazugegeben.
Die DNA-Konzentration wurde mit einem Photospektrometer
(Amersham Pharmacia Biotech, Dübendorf, Schweiz) bei 260 nm bestimmt.

Material & Methoden 27
2.5.2. Isolierung von Plasmid DNA
Die Isolierung von Plasmid DNA aus E. coli ÜN-Kulturen (25» 100ml)
erfolgte unter Verwendung des QIAGEN Midiprep/Maxiprep Systems
(QIAGEN, Basel, Schweiz) nach Anleitung des Herstellers.
2.5.3. Isolierung von X Phagen DNA
X Phagen DNA wurde wie durch Patterson und Dean (87) beschrieben
isoliert.
2.5.4. Isolierung von DNA Fragmenten aus Agarose Gel
Mit einem Restriktionsenzym verdaute chromosomale DNA wurde über
ein 0.8% präparatives Agarose Gel (Low Melt Agarose, Sigma, Buchs,
Schweiz) bei (2.6 V/cm) nach Bandengrösse aufgetrennt. Der entsprechende
Bandenbereich wurde mit einem sauberen Skalpell ausgeschnitten und die DNA
enthaltende Agarose in ein Eppendorf-Röhrchen gegeben. Dieses Eppendorf-
Röhrchen wurde anschliessend für 2 min in Trockeneis/EtOH
schocktiefgefroren und dann für 10 min bei 16000g abzentrifugiert. Der
erhaltene Überstand wurde in ein separates Eppendorf überführt und die
verbleibende Agarose erneut schocktiefgefroren und zentrifugiert. Der
Überstand wurde gesammelt und zur restlichen Agarose 100 Lil TE-Puffer
hinzugefügt. Die Agarose wurde an der Eppendorfwand zerquetscht und von
neuem schocktiefgefroren und zentrifugiert.
Die gesamten gesammelten Überstände wurde dann erst mit einem
Volumen Phenol/Chloroform/Isoamylalkohol und anschliessend mit einem
Volumen Chloroform extrahiert. Zum resultierenden Überstand wurden 1/10
Volumen 3 M Natrium-Acetat pH 4.8 und zwei Volumen Ethanol zugegeben,
gut gemischt, für 10 min auf Eis gefällt und abzentrifugiert. Nach Waschen mit
80% Ethanol und Trocknen wurde die DNA in 0.1 x TE aufgenommen.

Material & Methoden 28
2.6. RNA-Isolierungsmethoden
50-100 ml LB-Medium mit oder ohne Antibiotikum wurden mit 1/100
Volumen einer S. aureus ÜN-Kultur angeimpft. Bei 37 °C wurde diese
Hauptkultur bis zur gewünschten OD600 wachsen gelassen. Wenn die Zellen mit
EtOH gestresst werden sollten, wurde nach Erreichen einer OD600 von 0.3 6%
v/v 100% EtOH zugegeben und die Zellen daraufhin bis zu einer OD600 von 0.6
wachsen gelassen. 35 ml wurden für 5 min bei 11000g zentrifugiert, das Pellet
in 500 Ltl kaltem TKM-Puffer (10 mM Tris-HCl, pH 7.4; 10 mM KCl; 5 mM
MgCl2) resuspendiert und nach Zugabe von 10 ul Lysozym (10 mg/ml) und 10
}il Lysostaphin (10 mg/ml) bis zur sichtbaren Lyse bei 37 °C unter Schütteln
inkubiert. Unmittelbar nach der Lyse wurde die Suspension in
Trockeneis/EtOH schocktiefgefroren und bis zur Weiterverarbeitung bei -80
°C aufbewahrt.
Das Lysat wurde bei 65 °C für 1 min aufgetaut, bevor es mit je 40 pl
20% SDS und 3 M NaOAc (pH 5.2) und mit 600 jLtl saurem auf 65 °C
erwärmten Phenol vermischt und kräftig gevortext wurde. Nach 4 min
Inkubation bei 65 °C wurde erneut gevortext und anschliessend abzentrifugiert.
Nach Wiederholung der Extraktion mit saurem Phenol folgte eine
herkömmliche Phenol/Chloroform Extraktion, gefolgt von einer Chloroform
Extraktion. Die Fällung erfolgte mit 2 Volumen 100% EtOH während 30 min
bei -20 °C. Schliesslich wurde die RNA einmal mit 80% EtOH gewaschen, in
50-100 ml DEPC-behandeltem Wasser resuspendiert und die Konzentration im
Photospektrometer gemessen.
2.7. Northern- und Southern-Analyse
2.7.1. Herstellung von DNA Sonden
Die Herstellung radioaktiv markierter DNA-Sonden erfolgte unter
Verwendung des Random Primed DNA Labeling Kits (Boehringer Mannheim).

Material & Methoden 29
9 pT DNA (30-100 ng) wurde für 5 min bei 95 °C denaturiert und sofort
eisgekühlt. In einem 20 p.1 Ansatz wurde die DNA mit 2 \il Reaktionsmix, 3 pl
Nukleotidmix (0.5 M dATP; 0.5 M dGTP; 0.5 M dTTP), 0.5 units Klenow-
Fragment und 5 p.1 a32P-dCTP (50 pCi) für 30 min bei 37 °C inkubiert und
von nicht eingebauten Nukleotiden über eine G-50-Sepharose-Säule
(Boehringer Mannheim) abgetrennt. Vor Zugabe zum Hybridisierungs-Mix
wurde zur markierten DNA 100 Lil Heringssperma-DNA (10 mg/ml;
Boehringer Mannheim) zugegeben und die Mischung anschliessend während 5
min bei 95 °C denaturiert.
Nicht-radioaktive Sonden wurden mittels PCR unter Verwendung des
markierten Nukleotids DIG-11-dUTP (Digoxigenin-ll-2'-deoxy-uridine-5'-
triphosphate, Boehringer Mannheim, Rotkreuz, Schweiz) hergestellt.
2.7.2. Northern Blot
10 pig Gesamt-RNA wurde über ein 1.2% Formaldehyd-Agarose-Gel
(21.4 ml 37% Formaldehyd, 23.6 ml 5x MOPS Puffer pH 7, 0.96 g Agarose in
75 ml DEPC behandeltem Wasser) bei 6.2 V/cm elektrophoretisch aufgetrennt
und mittels der von Münch (75) beschriebenen "downward-alkaline-transfer"
Methode auf eine Nylonmembran (FALL, Biodyne® Nylon Membranes, East
Hills, NY, USA) übertragen. Nach dem Transfer wurde die Membran kurz in
2 x SSC gewaschen und im Anschluss daran wurde die transferierte RNA im
UV-Stratalinker® 1800 (Stratagene, Zürich, Schweiz) durch UV-Licht (254
nm, 120 mJ) auf der noch feuchten Membran fixiert.
2.7.3. Southern Blot
Chromosomale DNA aus 5. aureus (je 1 jig) wurde mit verschiedenen
Restriktionsenzymen verdaut und über ein 0.8% Agarose-Gel elektrophoretisch
aufgetrennt. Die erhaltenen Gele wurden jeweils 10 min in 0.25 M HCl
depuriniert, für 20 min in 0.4 M NaOH denaturiert und anschliessend auf eine

Material & Methoden 30
mit 0.4 M NaOH vorbehandelte Nylonmembran transferiert. Nach dem
Transfer wurde die Membran für fünf Minuten in 2 x SSC gewaschen und im
Anschluss daran wurde die DNA im UV-Stratalinker® 1800 (Stratagene) durch
UV-Licht (254 nm, 120 ml) auf der noch feuchten Membran fixiert.
2.7.4. Northern Hybridisierung
Die 30 minütige Prähybridisierung und die Hybridisierung über Nacht
erfolgte in Hybridisierungspuffer (0.5 M NaHP04-Puffer, pH 7.2; 1%
lyophilisiertes Rinderserumalbumin; 1 mM EDTA; 7% SDS) bei 65 °C. Als
Sonden wurden mittels Random primed labeling kit (Boehringer Mannheim)
markierte PCR Fragmente verwendet. Nach über Nacht erfolgter
Hybridisierung wurde die Membran für 5 min mit Wash 1 (40 mM NaHP04-
Puffer, pH 7.2; 0.5% lyophilisiertes Rinderserumalbumin; 1 mM EDTA; 5%
SDS) gewaschen, danach so lange für je 3 min mit Wash 2 (40 mM NaHP04-
Puffer, pH 7.2; 1 mM EDTA; 1% SDS), bis die Waschlösung nicht mehr
radioaktiv war. Nach dem Entfernen von SDS mit Wash 3 (100 mM NaHP04-
Puffer, pH 7.2) wurden die Filter bei -70 °C auf Röntgenfilmen (Fuji X-Ray,
Fuijfilm, Dielsdorf, Schweiz) exponiert.
2.7.5. Southern Hybridisierung
Die Hybridisierungsschritte waren, sofern radioaktiv gearbeitet wurde,
die gleichen wie bei der Northern Analyse.
Wenn mit DIG-Sonden hybridisiert wurde, wurde nach der im „The DIG
System User's Guide for Filter Hybridization" vom DIG-Kit Lieferanten
Boehringer Mannheim vorgeschlagenen Methode vorgegangen (The DIG
System User's Guide for Filter Hybridization, Boehringer Mannheim).

Material & Methoden 31
2.8. Klonieren von glmM
Ein 10.5 kb chromosomales Xbal Fragment aus dem Stamm PG108, das
die rechte Seite (Junction right, JR) des Transposons Tn55i enthielt, wurde mit
Tn55/ als Sonde identifiziert und in den Vektor pTZ18R (Genescribe-Z,
USB Corp., Cleveland, Ohio, USA) kloniert. Dieses Fragment wurde
verwendet um in einem chromosomalen Pstl Verdau des Stammes BB270 ein 5
kb grosses Fragment zu identifizieren, welches das gesamte glmM Operon
enthielt. Dieses Fragment wurde in den Vektor pAW8 subkloniert, woraus
schliesslich der Vektor pPG76 resultierte.
2.9. Sequenzierungen
2.9.1. Nicht-radioaktive Sequenzierung
Das 5 kb grosse klonierte Insert von pPG76 wurde mit verschiedenen
Restriktionsenzymen verdaut und in den E.coli Vektor pTZ18R (Genescribe-
Z, USB Corp.) subkloniert. Anschliessend wurden die verschiedenen
Subklone mit dem halbautomatischen ALF-Sequenator (Pharmacia, Uppsala,
Schweden) unter Verwendung des Thermo Sequenase fluorescent-labeled
primer cycle sequencing kit (Amersham Life Science) sequenziert.
2.9.2. Radioaktive Sequenzierung
Für die radioaktive Sequenzierung mit a35S-dATP wurde der T7
Sequenase Kit vs. 2.0 (Amersham Life Science) verwendet und nach den
Angaben des Herstellers vorgegangen.

Material & Methoden 32
2.10. PCR (Polymerase Chain Reaction)
Polymerase Kettenreaktion (PCR, (95)) wurden nach den Vorschriften
der Hersteller der verwendeten Polymerasen in einem Techne Progene
(Techne, Cambridge, Grossbritannien) oder DNA Thermal Cycler (Perkin
Eimer Cetus, Rotkreuz, Schweiz) durchgeführt. Als Polymerasen wurde die
AmphtaqGold DNA Polymerase (Perkin Eimer) oder die pfu Polymerase
(Perkin Eimer) verwendet.
2.11. RT-PCR (Reverse Transcriptase Polymerase Chain Reaction)
Für die Reverse Transkriptions Polymerase Ketten Reaktion (RT-PCR)
wurden 3 p,g Gesamt-RNA während 5 min bei 65 °C inkubiert um
Sekundärstrukturen zu zerstören. Die entsprechenden Primer wurden während
10 min bei RT anhybridisiert. Die reverse Transkription erfolgte während
einer Stunde bei 42 °C unter Verwendung der Reversen Transkriptase
SuperScript II (Gibco, Life Technologies AG). Anschliessend wurde der ganze
Ansatz für 5 min bei 95 °C inkubiert um die Reverse Transkriptase zu
inaktivieren und schliesslich für 5 min auf Eis gekühlt.
Die erhaltene cDNA wurde mittels PCR unter Verwendung
entsprechender Primer amplifiziert. Um eine Verunreinigung der Gesamt-
RNA Ansätze mit chromosomaler DNA auszuschliessen wurden dieselben
Gesamt-RNA Ansätze ohne vorhergehende RT-PCR mittels PCR amplifiziert.
Die Sequenz der Primer kann der Tabelle 5 entnommen werden.
2.12. Primer-Extension
Die für die Primer Extenison verwendeten Primer wurden mit y^P-
dATP (Amersham Life Science) unter Verwendung einer T4

Material & Methoden 33
Tabelle 5: für PCR und RT-PCR verwendete Primer
Primersequenz (5'-»3') Position des 5' Nukleotids
im glmM-Operon
Nummer
in Fig. 8
TGATTTAGGACCAACAGC
ACCTAATGAAACCGCCTG
AGGGAACTAAAGCGATAC
GGGGCTTGAGATTTATTG
CAACTGGATTATGAGAGG
TTACTTTGAAGGGGC
TTTATCTGTTACGCG
CAGACAATGGCAAATAAC
107 1
968 2
1307 3
2025 4
3286 5
3426 6
4116 7
1786 8
Primersequenz (5'—>3') Position des 5' Nukleotids
in fmhB
Nummer
in Fig. 26
TCAGAAACAAAGAAATTAACTG
TAAATCGTTAAAGCCGTACTTC
1
2
Tabelle 6: für Primer Extension verwendete Oligonukleotide
Oligonukleotidsequenz (5'—>3') Position des 5' Nukleotids im
g/mM-Operon; Fig. 9
GAAAAATCCATAACATCTCCTC
AAAGACCGTGATGAGAAG
TAGTTCCCTTAAAGACCGTGATGAG
ATAAAAAGAAAAACAATGCCAAAAG
GTTTGCGACACCTCTTACTCC
CTAGAACATAGCCACCGTATCTTCC
TTTCACCTTTATTATGTGCTAGAAC
1209
1305
1315
2072
3012
3070
3088

Material & Methoden 34
Polynukleotidkinase endmarkiert und über Sephadex®G-25 Säulen (Pharmacia
Biotech, Uppsala, Schweden) gereinigt. Die RNA (40 pig) und der Primer (500
cps) wurden zuerst für 5 min auf 85°C erhitzt und während 15 min bei 55°C
wurde der Primer anhybridisiert, kurz auf Eis abgekühlt und dann nach
Zugabe von Reverser Transkriptase (SuperScript II RT, Gibco, Life
Technologies AG, Basel, Schweiz) 1 h bei 42 °C inkubiert. Anschliessend
wurde die cDNA gefällt, in 9 jllI Stop Solution (Sequenase Stop Solution,
Amersham Life Science) aufgenommen, denaturiert und auf einem 6%
Sequenziergel zusammen mit der radioaktiven Sequenzierreaktion des gleichen
Primers aufgetrennt.
Die Sequenz der Primer kann der Tabelle 6 entnommen werden.
2.13. Herstellung eines transduzierenden Phagenlysats
0.3 ml einer ÜN-Kultur des Donorstammes wurde mit 5 mM CaCl2
versetzt und mit 0.1 ml verschiedener Verdünnungen des Phagen 80a infiziert.
Damit der Phage an die Bakterienzellen adsorbieren konnte, wurde der Ansatz
während 15 min bei RT inkubiert. Anschliessend wurde der Ansatz mit 4 ml
Softagar und 5 mM CaCl2 vermischt, auf Schafblut-Agarplatten ausplattiert
und ÜN bei 30 °C inkubiert. Am nächsten Tag wurde die Platte mit
konfluenter Lyse mit 1 ml LB versetzt mit 5 mM CaCl2 überschichtet, die
Softagarschicht zusammen mit dem 1 ml LB abgekratzt, kurz gevortext und
für 10 min bei 10000g zentrifugiert. Der Überstand wurde durch einen 0.45
p.m Filter (Millipore S.A., Molsheim, Frankreich) gefiltert und bei 4 °C
gelagert.
2.14. Transduktion von S. aureus
Für Transduktionen wurden 0.3 ml einer ÜN-Kultur des
Akzeptorstammes mit 5 mM CaCl2 versetzt und mit 0.1 ml der verschiedenen

Material & Methoden 35
Verdünnungen des transduzierenden Phagen 80a infiziert. Nach 15 min
Inkubation bei RT wurde die Transduktionsmischung mit 4 ml Softagar, der 20
mM Natriumeitrat enthielt, gemischt und sofort auf LB-Agar-Platten mit 20
p.g/ml Erythromycin ausplattiert.
Nach 48 h Inkubation bei 37 °C konnten Transduktanten gepickt werden.
2.15. Kompetente Zellen
2.15.1. Herstellung DMSO-kompetenter E. coli Zellen
Die Gewinnung DMSO-kompetenter E. coli Zellen wurde nach der
Methode von Inoue et al. (45) vorgenommen. Im allgemeinen wurde der E.
coli Stamm DH5a (Gibco Life Technologies AG) verwendet.
2.15.2. Herstellung elektrokompetenter DH10B E. coli Zellen
Von einer ÜN-Kultur wurden 100 ml LB 1:100 angeimpft und bis zu
einer OD600 von 0.5-0.6 wachsen gelassen. Die für 10 min bei 2000g
abzentrifugierten Zellen wurden in gleichem Volumen eiskaltem Wasser
resuspendiert und nochmals 10 min bei 2000g zentrifugiert. Darauf wurde das
Pellet in 0.5 Volumen eiskaltem Wasser resuspendiert und wieder
zentrifugiert. Nach Aufnahme in 1-1.5 ml eiskaltem Wasser wurde die
Zellsuspension nochmals kurz abzentrifugiert und das Zellpellet in 500 pl
Wasser eiskaltem Wasser aufgenommen und bis zur Transformation auf Eis
stehen gelassen oder bei -70 °C aufbewahrt.
2.15.3. Herstellung elektrokompetenter S. aureus RN4220
500 ml LB wurden mit einer ÜN-Vorkultur auf eine OD600 von 0.05
angeimpft und bei 37 °C unter starkem Schütteln inkubiert, bis eine OD600 von

Material & Methoden 36
0.5-0.6 erreicht wurde. Bei 2000xg wurden die Zellen bei 4 °C pelletiert.
Anschliessend wurden die Zellen viermal mit eiskalter 0.5 M Saccharoselösung
gewaschen und zwar mit jeweils 0.5, 0.25, 0.125 und 0.0625 des
Kulturvolumens. Nach dem zweiten Waschgang wurden die resuspendierten
Zellen für 2-4 h auf Eis stehengelassen. Zuletzt wurden die Zellen in 2.5 ml
eiskaltem, filtersterilisiertem 10% Glycerin resuspendiert, in 50 pf Aliquots im
Trockeneis/EtOH-Bad schocktiefgefroren und bei -70 °C aufbewahrt.
2.16. Transformationen
2.16.1. Transformation von DMSO-kompetenten E. coli Zellen
Die Transformation DMSO-kompetenter E. coli Zellen mit
zirkularisierter Plasmid-DNA erfolgte wie bei Inoue et al. (45) beschrieben.
2.16.2. Transformation elektrokompetenter E. coli Zellen
Zu 40-80 pJ elektrokompetenten Zellen wurden 1-2 u.1 des
Ligationsansatzes in vorgekühlten 0.2 cm Elektroporationsküvetten (BioRad,
Glattbrugg, Schweiz) zugegeben und für 1 min auf Eis belassen. Nach
erfolgtem Puls (Gene, BioRad, Glattbrugg, Schweiz: 2.5 kV, 200 O, 25 uF)
wurde sofort 1 ml SOC-Medium zugegeben und der Ansatz für 1 h bei 37 °C
leicht geschüttelt. Anschliessend wurden 100-150 pl des Ansatzes auf
entsprechende Selektivplatten ausgestrichen und ÜN bei 37 °C inkubiert. Der
Rest des Ansatzes wurde bei 4 °C aufgehoben.
2.16.3. Transformation elektrokompetenter S. aureus RN4220
50 pl kompetente Zellen wurden auf Eis aufgetaut, mit einer
entsprechenden Menge Plasmid-DNA gemischt (300-1000 ng) und für 30 min

Material & Methoden 37
auf Eis inkubiert. Das Zellen-DNA-Gemisch wurde in eine vorgekühlte 0.1 cm
Elektroporationsküvette (BioRad) transferiert und anschliessend wurden die
Zellen mit einem Pulser (Gene Pulser, BioRad; 1.5 kV, 100 D, 25 jiF)
elektroporiert. Sofort wurde 0.9 ml eiskaltes SOC-Medium zugegeben und der
Ansatz für 1 h bei 37 °C inkubiert, bevor 100-150 jil auf entsprechende
Selektivplatten ausgestrichen wurde. Der Rest wurde bei 4 °C aufbewahrt.
2.17. Nachweis der Penicillin-Bindeproteine (PBP)
Zellmembran-Proteine wurden nach Curtis et al. (23) mittels
verschiedener Zentrifugationsschritten aus mit Lysostaphin (Sigma, Buchs,
Schweiz) lysierten ÜN-Kulturen gewonnen. Die mit 10 mg/ml (Phenyl-4(n)~
3H)Benzylpenicillin (Endkonzentration; 11.9 Ci/mmol, Amersham Life
Science) für 10 min bei 30 °C markierten PBP wurden über ein 12% SDS-Gel
aufgetrennt. Nach der Auftrennung wurde das Gel mit Coomassie Blau gefärbt
und photographiert. Anschliessend wurde das Gel für 20 min in Enhancer
(EN3-Hance, New England Nuclear, Boston, USA) inkubiert. Der Nachweis
der PBP erfolgte mittels Fluorographie. Das Gel wurde während vier Wochen
bei -70 °C auf einem Röntgenfilmen (Fuji X-Ray, Fuijfilm, Dielsdorf,
Schweiz) exponiert.
2.18. Western Blot Analyse
Für die Western Blot Analyse wurden zytoplasmatische Proteine und
Membranproteine aus mittels Lysostaphin lysierten Kulturen gewonnen und
nach Standardprotokollen mittels SDS-Gel Elektrophorese aufgetrennt und
geblottet.
Anti GlmM Antikörper wurden von H. Komatsuzawa, University School
of Dentristy, Hiroshima, Japan, erhalten.

Material & Methoden 38
2.19. Resistenzbestimmungen
Minimale Hemmkonzentrationen (MHK) von den entsprechenden
Antibiotika wurden mittels E-Test (AB BIODISK, Solna, Schweden) auf
Müller-Hinton-Agar-Platten oder mittels der Mikrodilution-Methode nach
NCCLS (79) in LB-Medium durchgeführt. Die Zellzahl wurde auf den 0.5
McFarland Trübungsstandard eingestellt, was 0.5 x 108 Bakterienzellen/ml
entspricht.
2.20. Populationsanalysen
Populationsanalysen von entsprechenden Bakterienstämmen wurden von
ÜN-Kulturen gemacht indem Aliquote von 0.1 ml aus einer Verdünnungsreihe
von 10"1 bis 10"9 auf frisch gegossene LB-Agar-Platten mit steigender
Antibiotika-Konzentration ausgestrichen wurde. Nach 48 h Inkubationszeit bei
35 °C wurden die gewachsenen Kolonien gezählt und graphisch dargestellt.
2.21. Autolyse
Um die spontane Autolyse zu bestimmen wurden exponentiel! wachsende
Zellen (OD600 = 0.6) geerntet, zweimal mit physiologischem Phosphatpuffer pH
7.0 gewaschen und in 0.05 M Tris-HCl pH 7.5 auf eine OD600 von 0.5
resuspendiert. Die resuspendierten Zellen wurden bei 37 °C inkubiert und die
mit der Zeit wegen der Zunahme lysierter Zellen abnehmende Trübung
gemessen und entsprechend graphisch dargestellt.

Material & Methoden 39
2.22. Wachstumskurven
Von einer S. aureus ÜN-Kultur wurde eine Probe entnommen,
entsprechend verdünnt und die OD600 photospektrometrisch gemessen. Anhand
des erhaltenen Wertes wurde in einem Gesamtvolumen von 100 ml LB eine
OD600 zwischen 0.02 und 0.05 eingestellt. Die Kulturen wurden anschliessend
bei 37 °C geschüttelt, von Zeit zu Zeit Proben entnommen und ihre OD600
gemessen.

Resultate 40
3. Resultate
3.1. Das glmM-System
glmM ist ein fem Faktor und wurde mittels Transposon-
Insertionsinaktivierung mit Tn55i auf dem Smal Fragment I lokalisiert. Seine
Inaktivativierung erhöhte die Empfindlichkeit gegenüber ß-Laktamen, wobei
es aber immer noch zur Ausbildung hochresistenter Subpopulationen kam.
Weiter führte die Inaktivierung zu einer Telcoplanin-Hyperempfindlichkeit.
3.1.1. Phänotypische Charakterisierung der Insertion Q12F
glmM::Tn551
Populationsanalyse, Minimale Hemm-Konzentrationen (MHK)
Charakteristisch für MRSA ist die heterogene Expression der
Methicillin-Resistenz. Alle Zellen des MRSA-Stammes BB270 besitzen eine
bestimmte Grundresistenz von etwa 4-8 pg/ml Methicillin. Eine kleine
Subpopulation ist aber fähig, bei Konzentrationen von über 100 pg/ml
Methicillin zu wachsen. Bei einigen hochresistenten MRSA-Stämmen wie COL
zeigen alle Zellen eine hohe Grundresistenz; solche Stämme werden auch
homogen hochresistente MRSA genannt. Die Höhe der Resistenz ist durch
chromosomale Faktoren bedingt und nicht abhängig von der Menge an PBP2'
(94). Über 40 solcher auf dem Chromosom verteilter Gene sind bereits mittels
Tn551 Insertionsinaktivierung identifiziert worden (13, 25, 111). Ursprünglich
sind diese Gene fem oder aux Faktoren genannt worden, werden aber, sobald
sie charakterisiert sind, nach ihrer eigentlichen Funktion umbenannt.
Die Inaktivierung des Phosphoglukosamin-Mutase Gens glmM,
ursprünglich femD, mittels Tn557 Insertion, verleiht eine erhöhte
Empfindlichkeit gegenüber Methicillin. Dabei wird sowohl die Grundresistenz,

Resultate 41
als auch die Anzahl von Subklonen, die in Anwesenheit von mehr als 10 pg/ml
Methicillin wachsen können, reduziert. Die Insertion D12F glmM::Tn551
reduzierte die MHK von 6 u.g/ml im Methicillin-resistenten Ausgangsstamm
BB270 auf 0.38 pg/ml Methicillin in der Mutante PG108 (Tab. 7). Die MHK
beim Stamm PG108 lag damit sogar unter derjenigen des isogenen MSSA
Stammes BB255. Dennoch bildete PG108 eine kleine Subpopulation (10"2-10"7),
die bei höheren Methicillinkonzentrationen von bis 128 u.g/ml Methicillin
wachsen konnte (Fig. 3). Diese hochresistenten Subklone waren immer noch
Erythromycin-resistent, was schliessen Hess, dass sie die Tn55i Insertion noch
trugen. Einer dieser hochresistenten Subklone, Stamm PG100, wurde
eingehender untersucht. Der Stamm PG100 zeigte im Vergleich zum
Ausgangsstamm BB270 eine erhöhte Resistenz gegenüber allen ß-Laktamen
und brachte auch eine grössere hochresistente Subpopulation hervor (Fig. 3).
Dieses charakteristische Resistenzprofil von Stamm PG100 blieb auch nach
Subkultivieren in LB ohne Antibiotikazugabe, sowie nach Lagerung bei -70 °C
erhalten, so dass angenommen werden konnte, dass es sich dabei um eine
Mutante handelte.
Um herauszufinden, ob die hohe Methicillin-Resistenz in PG100 mit der
Erythromycin-Resistenz kotransduzierbar ist, wurde die Erythromycin-
Resistenz von PG100 in BB270 zurücktransduziert. Die Transduktanten,
repräsentiert durch den Stamm PG105 wurden aber Methicillin-empfindlich
wie PG108 (Tab. 7; Fig. 3). Hybridisierungen von i7mdIII geschnittener
chromosomaler DNA der Stämme PG108, PG100 und PG105 mit Tn55i-DNA
als Probe ergaben die gleichen Bandenmuster. Damit wurde gezeigt, dass
glmM in allen drei Stämmen, auch in PG100, immer noch durch Tn55i
unterbrochen war (Fig. 4B; Fig. 5). Daher wurde folgendes postuliert: eine
suppressorische oder kompensatorische Mutation, hmrDl genannt für ,,/ngh-
level methicillin-resistant suppressor mutant of femD", scheint für die hohe
Resistenz in PG100 verantwortlich und ist, da nicht kotransduzierbar, nicht mit
glmM verknüpft.

Resultate 42
Tabelle 7: Minimale Hemmkonzentrationen verschiedener Zellwand¬
gerichteter Antibiotika gegen verschiedene glmM Mutanten und ihre
parentalen Stämme.
Stämme MHK(ug/ml)
VA TP OX ME IP
BB255 1.5 1.0 0.19 0.75 0.032
PG27 1.5 0.25 0.064 0.25 0.012
BB270 1.5 1.0 12 6 0.25
PG108 0.75 0.064 0.094 0.38 0.032
PG100 1.0 0.064 >256 >256 f^" ^J a£j
PG105 0.75 0.064 0.19 0.75 0.047
COL 1.5 2 >256 >256 >32
PG106 1.0 0.25 0.19 0.75 0.047
PG79 1.5 1.0 16 12 0.125
PG231 1.5 0.125 16 6 0.125
PG232 1.5 0.38 >256 >256 >32
PG217 0.75 0.19 0.38 0.38 0.047
VA, Vancomycin; TP, Teicoplanin; OX, Oxacillin; ME, Methicillin; IP, Imipenem.

O
"sa
1 10 100 1000 10000
Methicillin pg/ml
Figur 3: Populationsanalyse von Uebernachtkulturen. Kolonie¬
formende Einheiten nach 48 h Inkubation auf LB-Agar Platten mit
zunehmender Methicillin-Konzentration. #, BB255; , BB270;
, PG108; A, PG100; A, PG105; O, PG79

Resultate 44
abcdefghi abcdefghi
<!=3
A B
Figur 4: Southern Blot von Hindlll verdauter chromosomaler DNA
verschiedener Wildtypen und ihren g//;/A/-Mutanten hybridisiert mit (A)einem g/mM-Fragment und (B) mit Tn55 7. a, COL; b, BB255; c,
BB270; d, PG106; e, PG27; f, PG108; g, Marker; h, PG100; i, PG105.
Pfeil zeigt auf das ungefähr 1 kb grosse Tn557 interne Hindlll
Fragment (Fig. 5).

Resultate 45
Tn551
CAH
ermB
KA SB HAC H X B AC
lkb
Figur 5: Restriktionskarte von Tn55i. A, Aval; B, Bglll; C,
CM; H, HindlU; H, Hpal; S, Sali; X, Xbal.

Resultate 46
Der MSSA Stamm BB255 erwies sich im Vergleich zu den mec
tragenden MRSA Stämmen in Transduktionen als schlechterer Rezipient. Es
wurden häufig Erythromycin-resistente Kolonien erhalten, die sich bei
genauerer Untersuchung als abortive Transduktanten erwiesen. Erst nach
mehreren Versuchen und längerer Inkubation der Transduktanten konnten
stabile BB255 glmM Mutanten erhalten werden, die sich aber phänotypisch
auffällig verhielten. Die Kolonien zeigten entweder eine gelb-orange
Verfärbung oder sie waren sehr klein. Die repräsentative BB255 glmM
Mutante PG27 zeigte bei der Hybridisierung mit Tn55i das gleiche
Bandenmuster wie alle anderen glmM Mutanten (Fig. 4B). Die Tn55i Insertion
hatte aber erwartungsgemäss weniger Einfluss auf die Empfindlichkeit gegen
Methicillin als bei einem MRSA Stamm (Tab. 7).
Die Transduktion von glmM::Tn551 in den homogen hochresistenten
Stamm COL ergab den Stamm PG106 und führte ebenfalls zu einer erhöhten
Methicillin-Empfindlichkeit (Tab. 7).
Neben den verschiedenen getesteten ß-Laktam Antibiotika Methicillin,
Oxacillin und Imipenem wurde der Einfluss der glmM Inaktivierung auf das
Verhalten gegen zwei weitere gegen die Zellwand gerichtete Antibiotika
untersucht. Die Insertion D12F (glmM::Tn551) hatte auf die Empfindlichkeit
gegen Vancomycin keine Wirkung (Tab. 7); die Empfindlichkeit gegen ein
weiteres Glykopeptid, Teicoplanin, nahm jedoch in allen MRSA glmM
Mutanten, auch im hochresistenten Suppressor Stamm PG100, zu (Tab. 7). Die
postulierte hmrDl Mutation beeinflusst demnach nur die Methicillin-Resistenz,
wohingegen die erhöhte Teicoplanin-Empfindlichkeit mit dem immer noch
inaktivierten glmM in Zusammenhang stehen muss.
Autolyse
Die spontane Autolyse zeigte nur einen geringen Unterschied zwischen
dem Wildtyp BB270 und seiner glmM Mutante PG108 (Fig. 6). Die Tn557
Insertion in glmM scheint auf die Autolyse keinen grossen Einfluss zu haben.

Resultate 47
o
o
O
o
0.45-
0.35
0.25 t r
0 40 80 120 160 200
min
Figur 6: Autolyse. Spontane Autolyse der Stämme, BB270; ,
PG108undA,PG100.

Resultate 48
Die Suppressor Mutante PG100 hingegen autolysierte im Vergleich dazu
deutlich langsamer, was mit der postulierten Mutation, die zu der erhöhten
Methicillin-Resistenz geführt hatte, zusammenhängen könnte.
3.1.2. Das glmM Operon
Klonieren und Sequenzieren
Um das Wildtyp Allel von glmM zu klonieren wurde Pstl-verdaute
chromosomale Wildtyp BB270 DNA mit dem 2.2 kb grossen CM Fragment
aus dem Plasmid pBBB106, das sowohl die Jr von Tn557 als auch 10 kb der
distalen Region von glmM enthielt, hybridisiert (Fig. 7). Die resultierende
Bande war um die 5 kb gross. Entsprechend wurde der 5 kb Bereich Pstl-
verdauter chromosomaler Wildtyp BB270 DNA in den Pstl geschnittenen
Shuttle Vektor pAW8 kloniert. Daraus resultierte das Plasmid pPG76, das
später auch zur Komplementation verwendet wurde.
Der gesamte Insert wurde mittels primer walking Methode sequenziert
und analysiert. Er beinhaltete vier offene Leseraster (ORF), die alle die gleiche
Orientierung aufwiesen (Fig. 8).
Analysieren der verschiedenen offenen Leserastern
Der erste ORF beginnt bei Nukleotid 107 und endet bei Nukleotid 1013
(Fig. 9). Er kodiert für ein 302 Aminosäuren (AS) grosses Protein mit einem
errechneten Molekulargewicht von 33 kDa. Datenbankvergleiche mit diesem
302 AS grossen Protein zeigten in einem 300 AS überlappenden Bereich eine
Identität von 53% und eine Ähnlichkeit von 73% mit der AS-Sequenz der
Arginase von Bacillus caldovelox (Swissprot accession no. p53608) und eine
Identität von 52% und eine Ähnlichkeit von 70% mit der von Bacillus subtilis
(Swissprot accession no. p39138). Das Enzym Arginase katalysiert den Abbau
von Arginin zu Ornithin und Harnstoff. Ein Vergleich der im aktiven Zentrum

2.2kbClalFragment
JrTn551
YA
XB
AvCH
PH
CP
PP
AX
kb
Figur
7:Ausschnitt
ausdem
10.5kbXbal
Insert
inPlasmidpBBBlOö.
Das
alsProbe
verwendete
2.2kb
grosseC/al-Fragment
istangegeben.A,AccI;Av,Aval;B,Bg
lll;
C,
Clal
;
H,Hindll;P,
Pstl:X,Xbal.

tTn55i
j
14
argl
orfJ
orß
glmM
AH
HC
HHN
HN
kb
Figur
8:RestriktionskartederRegiondesglmMOperons.
P,Ps
tl;A,Asp718;H,Hindll;C,
Clal
;
N,
Nsil.DieoffenenLeserastersindmitPfeilenangegeben,sowiediePositionundOr
ient
ieru
ngdesTn557.Diekleinennummerierten
Pfeilezeigen
diePositionundOr
ient
ieru
ngderfürdieRT-
PCR
verwendeten
Primer.
Die
daraus
resultierenden
Produkte
sind
oberhalb
als
Linien
angegeben.
Die
fürdieNorthem
Blotsverwendetenundge
zeigte
nProben
sind
alsDoppellinien
angegeben.

TTTTTTTATGAAGTAATAAGATAAACAGATTATATGTGTAGCTATTGCTT GATAATGGTA
1 * * * * * * 60
AGCC-CATACATTATTCTTTAATTACATAGAGCAAAGGGGGACGCTTATGACÄAAGACAAA
61 * * * SD* * * 120
1 M T K T K 5
AGCAATTGATATTATAGGTGCACCATCAACATTTGGACAAAGAAAATTAGGTGTTGATTT
121 ****** ]_go
6AIDIIGAPSTFGQRKLGVDL 25
AGGACCAACAGCAATTAGATATGCTGGATTAATTTCAAGATTAAAGCAATTAGACCTTGA
181 ****** 240
26GPTAIRYAGLISRLKQLDLD 45
TGTATATGACAAGGGGGATATTAAGGTACCTGCTGTGAACATTGAAAAATTTCATAGTGA
241 ****** 3oo
46VYDKGDIKVPAVNIEKFHSE 65
ACAAAAAGGATTAAGAAATTATGATGAAATTATAGATGTTAATCAAAAATTAAATAAAGA
301 * * * * * * 360
66QKGLRNYDEIIDVNQKLNKE 85
GGTTTCAGCATCAATTGAAAATAACAGATTTCCTCTAGTTCTTGGTGGAGATCATTCTAT
361 ****** 420
86VSASIENNRFPLVLGGDHSI 105
TGCGGTAGGTTCAGTATCAGCAATAAGTAAACATTATAATAATTTAGGTGTTATTTGGTA
421 ****** 4gQ
106 AVGSVSAISKHYNNLGVTWY 125
TGATGCACATGGTGATTTAAATATACCTGAAGAGTCACCAAGTGGAAATATTCATGGTAT
481 * * * * * * 540
126 DAHGDLNIPEESPSGNIHGM 145
GCCTCTAAGGATTTTGACAGGCGAAGGTCCCAAAGAACTTTTAGAATTAAATAGTAATGT
541 ****** 600
146 PLRILTGEGPKELLELNSNV 165
AATCAAGCCAGAAAACATCGTACTAATTGGTATGAGAGATTTAGATAAAGGTGAAAGACA
601 ****** 660
166 IKPENIVLIGMRDLDKGERQ 185
ATTTATCAAAGATCATAATATTAAAACATTTACTATGTCAGATATTGATAAATTGGGGAT
661 ****** 720
186 FIKDHNIKTFTMSDIDKLGI 205
AAAGGAAGTAATTGAAAATACAATAGAATATTTGAAGTCACGCAATGTTGATGGCGTTCA
721 ****** 73o
206KEVÏENTIEYLKSRNVDGVH 225

Resultate
TTTATCTTTAGATGTTGATGCTTTAGATCCGCTTGAAACGCCAGGCACTGGTACTAGAGT
781 ****** 340
226 LSLDVDALDPLETPGTGTRV 245
TTTGGGTGGTCTTAGTTATAGAGAAAGCCATTTTGCATTGGAATTACTGCATCAATCACA
841 * * * * * * 900
246 LGGLSYRESHFALELLHQSH 265
TTTAATTTCCTCAATGGATTTAGTTGAAGTAAATCCATTGGTTGACAGTAATAATCATTC
gOl * * * * * * 960
266 LISSMDLVEVNPLVDSNNHS 285
TGCTGAACAGGCGGTTTCATTAGGTGGAACATTTTTTGGTGAACCTTTATTATAAATAAA
961 ******1020
286AEQAVSLGGTFFGEPLL* 302
TGATTTGTAGTGAAAAAGTATÄTTTTGCTTTTGCACTACTTTTTTTAATCACTAAAATGA
1021 ******1080
TTAAGGGTAGTTATATC TTTAAAATAATTTTTTTCTATTAAAATATATGTCGT ATGA CAG
1081 ***** 1140
TGATGTAAATGATTGGTATAATGGGTATTATCGAAAAATATTCCCGGAGGAGATGTTATG
1141 * * * * SD* * 120Q
M 1
GATTTTTCCAACTTTTTTCAAAACCTCAGTACGTTAAAAATTGTAACGAGTATCCTTGAT
1201 ******1260
2DFSNFFQNLSTLKIVTSILD 21
TTACTGATAGTTTGGTATGTACTTTATCTTCTCATCACGGTCTTTAAGGGAACTAAAGCG
1261 ******1320
22LLIVWYVLYLLITVFKGTKA 41
ATACAATTACTTAAAGGGATATTAGTAATTGTTATTGGTCAGCAGATAAGTATGATATTG
1321 ******13S0
42IQLLKGILVIVIGQQISMIL 61
AACTTGACTGCAACATCTAAATTATTCGATATCGTTATTCAATGGGGGGTATTAGCTTTA
1381 ******1440
62NLTATSKLFDIVIQWGVLAL 81
ATAGTAATATTCCAACCAGAAATTAGACGTGCGTTAGAACAACTTGGTAGAGGTAGCTTT
1441 ******1500
82IVIFQPEIRRALEQLGRGSF 101
TTAAAACGCTATACTTCTAATACGTATAGTAAAGATGAAGAGAAATTGATTCAATCGGTT
1501 ******156û
102 L ?" R Y T S N T Y S K D E E K L I Q S V 121

Resultate 53
TCAAAGGCTGTGCAATATATGGCTAAAAGACGTATAGGTGCATTAATTGTCTTTGAAAAA
1561 ******1620
122 SKAVQYMAKRRIGALIVFEK 141
GAAACAGGTCTTCAAGATTATATTGAAACAGGTATTGCAATGGATTCAAATATTTCGCAA
1621 ******16gQ
142 ETGLQDYIETGIAMDSNISQ 161
GAACTTTTAATTAATGTCTTTATACCTAACACACCTTTACATGATGGTGCAATGATTATT1681 ******1740
162 ELLINVFTPNTPLHDGAMII 181
CAAGGCACGAAGATTGCAGCAGCAGCAAGTTATTTGCCATTGTCTGATAGTCCTAAGATS1741 ******1800
182 QGTKIAAAASYLPLSDSPKI 201
TCTAAAAGTTTGGGTACAAGACATAGAGCTGCGGTTGGTATTTCAGAAGTATCTGAGGCC
1801 ******186o
202 SKSLGTRHRAAVGISEVSEA 221
TTTACCGTTATTGTATCTGAAGAAACTGGTGATATTTCGGTAACATTTGATGGAAAATTS1861 ******1920
222 FTVIVSEETGDISVTFDGKL 241
CGACGAGACATTTCAAACGAAATTTTTGAAGAGTTGCTTGCTGAACATTGGTTTGGCACA1921 ******1930
242 RRDISNEIFEELLAEHWF_GT 261
CGCTTTCAAAAGAAAGGTGTGAAATAATATGCTAGAAAGTAAATGGGGCTTGAGATTTAT1981 ******2040
262/1R FQKKGVK* MLESKWGLRFI 269/11
TGCCTTTCTTTTGGCATTGTTTTTCTTTTTATCTGTTAACAATGTTTTTGGAAATATCTT2041 ******2100
12AFLLALFFFL SVNNVFGNI F 31
TAACACTGGTAATCTTGGTCAAAAGTCTAGTAAAACGATTCAAGATGTACCCGTTGAAAT2101 ******2160
32NTGNLGQKSSKTIQDVPVEI51
TCTTTATAACACTAAAGATTTGCATTTAACAAAAGCGCCTGAAACAGTTAATGTGACTAT2161 ******2220
52LYNTKDLHLTKAPETVNVTI71
TTCAGGACCACAATCAAAGATAATAAAAATTGAAAATCCAGAAGATTTAAGAGTAGTGAT2221 ******2280
72SGPQSKIIKIENPEDLRVVI91

Resultate 54
TGATTTATCAAATGCTAAAGCTGGAAAATATCAAGAGAAGTATCAAGTTAAAGGGTTAGC
2281 ******2340
92DLSNAKAGKYQEKYQVKGLA 111
TGATGACATTCATTATTCTGTAAAACCTAAATTAGCAAATATTACGCTTGAAAACAAAGT
2341 ******2400
112 DDIHYSVKPKLANITLENKV 131
AACTAAAAAGATGACAGTTCAACCTGATGTAAGTCAGAGTGATATTGATCCACTTTATAA
2401 ******2460
132 TKKMTVQPDVSQSDIDPLYK 151
AATTACAAAGCAAGAAGTTTCACCACAAACAGTTAAAGTAACAGGTGGAGAAGAACAATT
2461 ******2520
152 ITKQEVSPQTVKVTGGEEQL 171
GAATGATATCGCTTATTTAAAAGCCACTTTTAAAACTAATAAAAAGATTAATGGTGACAC
2521 ******2580
172 NDIAYLKATFKTNKKINGDT 191
AAAAGATGTCGCAGAAGTAACGGCTTTTGATAAAAAACTGAATAAATTAAATGTATCGAT
2581 ******2640
192 KDVAEVTAFDKKLNKLNVSI 211
TCAACCTAATGAAGTGAATTTACAAGTTAAAGTAGAGCCTTTTAGCAAAAAGGTTAAAGT
2641 ******2700
212 QPNEVNLQVKVEPFSKKVKV 231
AAATGTTAAACAGAAAGGTAGTTTAGCAGATGATAAAGAGTTAAGTTCGATTGATTTAGA
2701 ******2760
232 NVKQKGSLADDKELSSIDLE 251
AGATAAAGAAATTGAAATCTTCGGTAGTCGAGATGACTTACAAAATATAAGCGAAGTTGA
2761 ******2820
252 DKEIEIFGSRDDLQNISEVD 271
TGCAGAAGTAGATTTAGATGGTATTTCAGAATCAACTGAAAAGACTGTAAAAATCAATTT
2821 ******2880
272 AEVDLDGISESTEKTVKINL 291
ACCAGAACATGTCACTAAAGCACAACCAAGTGAAACGAAGGCTTATATAAATGTAAAATA
2881 ******2940
292PEHVTKAQPSETKAYINVK 310
AATAGCTAAATTAAAGGAGAGTAAACAATGGGAAAATATTTTGGTACAGACGGAGTAAGA
2941 * SD * * * * 3ooo
1 * M G K Y F G T D G V R 11

Resultate 55
GGTGTCGCAAACCAAGAACTAACACCTGAATTGGCATTTAAATTAGGAAGATACGGTGGC
3001 ******3060
12GVANQELTPELAFKLGRYGG 31
TATGTTCTAGCACATAATAAAGGTGAAAAACACCCACGTGTACTTGTAGGTCGCGATACT
3061 ******3120
32YVLAHNKGEKHPRVLVGRDT 51
AGAGTTTCAGGTGAAATGTTAGAATCAGCATTAATAGCTGGTTTGATTTCAATTGGTGCA
3121 ****** 3180
52RVSGEMLESALIAGLISIGA 71
GAAGTGATGCGATTAGGTATTATTTCAACACCAGGTGTTGCATATTTAACACGCGATATG
3181 ******3240
72EVMRLGIISTPGVAYLTRDM 91
GGTGCAGAGTTAGGTGTAATGATTTCAGCCTCTCATAATCCAGTTGCAGATAATGGTATT
3241 ****** 3300
92 G A E LGVMI SASHNPVADNGÏ 111
AAATTCTTTGGATCAGATGGTTTTAAACTATCAGATGAACAAGAAAATGAAATTGAAGCA
3301 ****** 3360
112 KFFGSDGFKLSDEQENEIEA 131
TTATTGGATCAAGAAAACCCAGAATTACCAAGACCAGTTGGCAATGATATTGTACATTAT
3361 ******3420
132 LLDQENPELPRPVGNDIVHY 151
TCAGATTACTTTGAAGGGGCACAAAAATATTTGAGCTATTTAAAATCAACAGTAGATGTT
3421 ******3480
152 SDYFEGAQKYLSYLKSTVDV 171
AACTTTGAAGGTTTGAAAATTGCTTTAGATGGCGCAAATGGTTCAACATCATCACTAGCG
3481 ******3540
172 NFEGLKIALDGANGSTSSLA 191
CCATTCTTATTTGGTGACTTAGAAGCAGATACTGAAACAATTGGATGTAGTCCTGATGGA
3541 ****** 3g00
192 PFLFGDLEADTETIGCSPDG 211
TATAATATCAATGAGAAATGTGGCTCTACACATCCTGAAAAATTAGCTGAAAAAGTAGTT
3601 ******3660
212 YNINEKCGSTHPEKLAEKVV 231
GAAACTGAAAGTGATTTTGGGTTAGCATTTGACGGCGATGGAGACAGAATCATAGCAGTA
3661 ******3720
232 ETESDFGLAFDGDGDRIIAV 251

Resultate 56
GATGAGAATGGTCAAATCGTTGACGGTGACCAAATTATGTTTATTATTGGTCAAGAAATG
3721 ****** 3780
252 DENGQIVDGDQIMFIIGQEM 271
CATAAAAATCAAGAATTGAATAATGACATGATTGTTTCTACTGTTATGAGTAATTTAGGT
3781 ******3840
272 HKNQELNNDMIVSTVMSNLG 291
TTTTACAAAGCGCTTGAACAAGAAGGAATTAAATCTAATAAAACTAAAGTTGGCGACAGA
3841 ******39oo
292 FYKALEQEGIKSNKTKVGDR 311
TATGTAGTAGAAGAAATGCGTCGCGGTAATTATAACTTAGGTGGAGAACAATCTGGACAT
3901 ******396o
312 YVVEEMRRGNYNLGGEQSGH 331
ATCGTTATGATGGATTACAATACAACTGGTGATGGTTTATTAACTGGTATTCAATTAGCT3961 ******4020
332 IVMMDYNTTGDGLLTGIQLA 351
TCTGTAATAAAAATGACTGGTAAATCACTAAGTGAATTAGCTGGACAAATGAAAAAATAT4021 Tn551 ******408O
352 SVIKMTGKSLSELAGQMKKY 371
CCACAATCATTAATTAACGTACGCGTAACAGATAAATATCGTGTTGAAGAAAATGTTGAC4081 ******4140
372 PQSLINVRVTDKYRVEENVD 391
GTTAAAGAAGTTATGACTAAAGTAGAAGTAGAAATGAATGGAGAAGGTCGAATTTTAGTA4141 ******4200
392 VKEVMTKVEVEMNGEGRILV 411
AGACCTTCTGGAACAGAACCATTAGTTCGTGTCATGGTTGAAGCAGCAACTGATGAAGAT4201 ******4260
412 RPSGTEPLVRVMVEAATDED 431
GCTGAAAGATTTGCACAACAAATAGCTGATGTGGTTCAAGATAAAATGGGATTAGATAAA4261 ******4320
432 AERFAQQIADVVQDKMGLDK 451
TAAÄTACTGTATTACAAATGAGCCGATGCGTATGCATCGGTTTTTTGTGTTTGTAGAAAT4321 ******43g0
AATTTATAGTACAAACGTAAAATGATATAAACAAAATAAAAACAAAGTAATCAATATGTA4381 ****** 444Q
ATATAAAATACACTGGTACTCAATATATAATGATGATAAAATTAATTTTAATTAGATAGA4441 ****** 4500

Resultate 57
GTTGCTTTGTGTTTTTAACGCAGATGCTACTACTTATCTTAACAGTTGATTAAGTGAAAT
4501 ******4560
CATTTAACAGCGAGAATAATCAACCAGGAGGATGACTTAATGAATTTATTCAGACAACAA
4561 ****** 4620
AAATTTAGTATCAGAAAATTTAATGTCGGTATTTTTTCAG
4621 * * * * 4660
Figur 9: Sequenz der Arginase argl und dem glmM Operon mit den ORF
orfl, orfl und glmM. Die möglichen Promotorsequenzen sind
fettgedruckt und unterstrichen, die Transkriptionsstartpunkte fettgedruckt
und kursiv geschrieben, Shine-Dalgarno Sequenzen sind kursiv
geschrieben, die Startkodons ATG sind fettgedruckt und die Inverted
Repeats sind unterstrichen. Die „active site" von GlmM ist fettgedruckt.
Tn557 Insertionsstelle Nukleotidposition 4022/4023.

Resultate 58
liegenden AS ergab eine Identität von über 70%. Daher wurde dieser ORF argl
genannt. Distal zu argl folgte ein Abschnitt von sieben Thymidin-Resten mit
vorgestelltem Inverted Repeat von 14 Nukleotiden, der für einen Faktor¬
unabhängigen RNA-Polymerase Terminator charakteristisch ist (Fig. 9) (17).
AG für diesen Terminator beträgt -3.3 kcal/mol.
Der zweite ORF, orfl genannt, startet bei Nukleotid 1198 und endet bei
Nukleotid 2005 (Fig. 9). Er kodiert für ein 269 AS grosses Protein mit einem
errechneten Molekulargewicht von 29.6 kDa. Dieses Protein hatte eine Identität
von 56% und eine Ähnlichkeit von 76% mit der AS-Sequenz von YbbP, einem
hypothetischen, von B. subtilis kodierten Protein. Zwei hydrophobe Regionen
im N-terminalen Bereich lassen vermuten, dass dieses Protein, dessen Aufgabe
noch nicht klar ist, membranassoziiert sein könnte (Fig. 10).
Der dritte ORF, orfl, von orfl nur durch vier Nukleotide getrennt,
beginnt bei Nukleotid 2009 und endet bei Nukleotid 2939 (Fig. 9). Er kodiert
für ein 310 AS grosses Protein mit einem errechneten Molekulargewicht von
34 kDa. Seine AS-Sequenz wies eine Identität von 28% und eine Ähnlichkeit
von 53% mit der AS-Sequenz von YbbR, einem wie YbbP hypothetischen, von
B. subtilis kodierten Protein auf. Bei orf2 konnte im Gegensatz zu allen
anderen ORF weder eine mögliche Promotor- noch eine wahrscheinliche
Shine-Dalgarno Sequenz ausfindig gemacht werden (Fig. 9).
Der vierte und letzte ORF, glmM, war 29 Nukleotide von orfl entfernt
und kodiert für ein 451 AS grosses Protein mit einem errechneten
Molekulargewicht von 49.6 kDa. Die glmM DNA Sequenz ist identisch mit der
des Phosphoglukosamin-Mutase Gens glmM von COL (117) (48) ausser einem
Nukleotidaustausch T zu C an der Position 3179, was zu einem
Aminosäurenaustausch von Valin zu einem Alanin an der Position 251 führte.
Die Tn55i Insertion unterbricht glmM im Stamm PG108 im C-terminalen
Bereich an Position 4022/4023 und verkürzt GlmM um 99 Aminosäure-Reste.
Trotz der Tn557 bedingten Unterbrechung von glmM, beinhaltet GlmM immer
noch das aktive Zentrum, das eine Phosphogluko- und Phosphomanno-Mutase
Phosphoserin Signatur enthält. Diese charakteristische Aminosäuren liegen

hydrophilic
I120 160 200
amino acid position
Figur 10: Hydrophobizitätsdiagramm von Orfl. Stark hydrophober N-
terminaler Bereich.

Resultate 60
zwischen den Aminosäuren 91 und 95, 95 und 110 und 111 und 115. Ein
Abschnitt von Thymidin-Resten mit vorgestelltem palindromischem
Sequenzabschnitt charakteristisch für einen Faktor-unabhängigen RNA-
Polymerase Terminator (Fig. 9) (17) folgte distal von glmM. AG für diesen
Terminator beträgt -10.3 kcal/mol.
3.1.3. Transkription der glmM Region
Die Analyse der vier verschiedenen ORF liess vermuten, dass argl
unabhängig von den ORF orfl, orfl und glmM transkribiert würde. Unklar
war allerdings, ob argl Teil eines grösseren, proximal gelegenen Operons ist
und ob orfl, orfl und glmM nur als dreizistronisches Transkript vorliegen
würde. Mittels Northern Blot Hybridisierungen und Primer-Extension
Analysen wurde die Transkription dieser Region untersucht.
Northern Blot
RNA von exponentiellen, spät-exponentiellen und stationären
Wachstumsstadien wurden vom MRSA BB270, MSSA BB255, von der glmM
Mutante PG108 und der hmrDl glmM Suppressor Mutante PG100 extrahiert
und mittels Northern Blot mit verschiedenen Proben (Fig. 8) analysiert.
Hybridisierung mit einer 0.69 kb langen g/mM-internen Probe, die die
Tn55i Insertionsstelle bedeckte, ergab bei allen Wachstumsstadien das gleiche
Bandenmuster. Bei den parentalen Stämmen BB255 und BB270 erhielt man
jeweils Transkripte von 3.2, 2.2 und 1.3 kb Grösse mit zunehmender
Bandenintensität (Fig. IIA). Abgeleitet von ihrer Grösse wurde davon
ausgegangen, dass das 3.2 kb Transkript die drei ORF orfl-orfl-glmM
repräsentierte. Das 1.3 kb Transkript schien vom glmM zu stammen, da es in
den Tn557-inaktivierten glmM Mutanten fehlte. Dafür war bei den Insertions-
Mutanten eine Bande von 4.7 kb Grösse vorhanden, bei der es sich
wahrscheinlich um ein „Readthrough" Produkt in das Tn55i handelte. Die 2.2
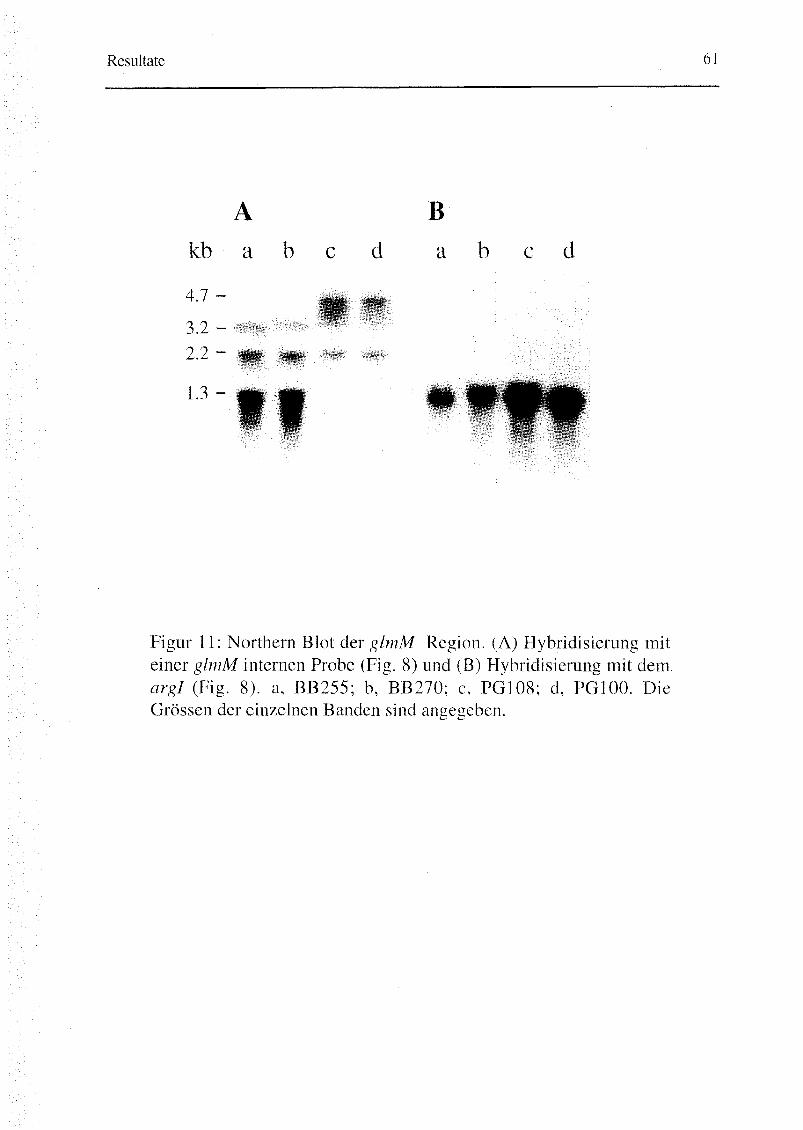
A
kb a b c
4.7 -
3.2 - »^
1.3-
B
a b c d
Figur 11 : Northern Blot der glmM Region. (A) Hybridisierung mit
einer glmM internen Probe (Fig. 8) und (B) Hybridisierung mit dem
argl (Fig. 8). a, BB255; b, BB270; c. PG108; d, PG100. Die
Grössen der einzelnen Banden sind angegeben.

Resultate 62
kb grosse Bande schien der Länge nach einem zweizistronischen Transkript
ausgehend vom orfl zu entsprechen. Dem widerspricht die Tatsache, dass eine
Bande in genau derselben Grösse bei den glmM Mutanten vorkommt, was
aufgrund der Tn55i Insertion nicht der Fall sein dürfte.
Aufgrund der Sequenzanalyse wurde postuliert, dass argl unabhängig
von den ORF orfl, orfl und glmM transkribiert werde. Daher wurde auch mit
einer argl internen Probe (Fig. 8) hybridisiert um dieses Postulat zu
überprüfen. Die erhaltene Bande hatte eine Grösse von 1.25 kb und war somit
um die 300 Nukleotide grösser als argl (Fig. IIB). Ihre Grösse und das
Vorhandensein sowohl in den Wildtypen und Mutanten haben gezeigt, dass argl
offenbar nicht zum glmM Operon gehört. Weshalb die Bandenintensität vom
argl Transkript in den Mutanten stärker war als im Wildtyp konnte nicht
geklärt werden.
Reverse Transkription
Einen weiteren Hinweis darauf, dass argl unabhängig vom postulierten
glmM Operon transkribiert wird, hat die Analyse der mRNA mittels reverser
Transkription und anschliessender PCR-Amplifikation geliefert. Sowohl eine
Amplifikation mit den Primern 1 und 5, als auch eine mit den Primern 1 und 8
lieferte kein RT-PCR Produkt, woraus geschlossen werden konnte, dass
anscheinend keine mRNA existierte, die sowohl argl als auch das glmM
Operon beinhaltete (Fig. 12). Die RT-PCR Analyse des glmM Operons im
Wildtyp BB270 und der entsprechenden glmM Mutante PG108 mit den
Primern 3 und 5 ergab sowohl im Wildtyp als auch in der Mutante ein 1969 bp
langes Produkt, das den grössten Teil von orfl, orfl und den Anfang von
glmM beinhaltete (Fig. 8). Mit den Primern 4 und 7 resultierte nur im Wildtyp
ein 2091 bp grosses, orfl-glmM abdeckendes Fragment, während in der glmM
Mutante, die das 5.2 kb grosse Tn55i Insert trug, kein Transkript zu sehen war
(Fig. 8). Da aber sowohl mit den Primern 3 und 5 und den Primern 4 und 5
bei der Mutante und beim Wildtyp ein Transkript zu sehen war und somit die

kb abcdefghi
Figur 12: RT-PCR mit verschiednen Primern von mRNA von
PG108 (Spuren a-d) und BB270 (Spuren f-i).
a, Primerpaar 5/3; b. Kontrolle von Primerpaar 5/3; c, Primerpaar
5/4; d, Kontrolle von Primerpaar 5/4; e. Marker; f, Kontrolle von
Primerpaar 5/3; g. Primerpaar 5/3; h, Kontrolle von Primerpaar II
4; i, Primerpaar 7/4.

Resultate 64
transkriptionelle Verbindung zwischen den drei ORF gezeigt war, konnte orfl-
orfl-glmM als dreizistronisches Operon bestätigt werden (Fig. 8; Fig. 12).
Transkriptionsstart-Bestimmung mittels Primer-Extension
Bei der Hybridisierung der Northern Blots mit der glmM internen Probe
erhielt man beim Wildtyp BB270 drei Banden verschiedener Grössen. Das 3.2
kb grosse Transkript wurde postuliert, die drei ORFs orfl-orfl-glmM zu
beinhalten (Fig. IIA). Das 1.3 kb grosse Transkript stammte, da es in den
Mutanten fehlte, von glmM ab (Fig. IIA). Dies deutete auf mindestens zwei
verschiedene Transkriptionsstart hin: einen am Anfang des Operons vor orfl
und einen zweiten vor glmM. Diese Annahme wurde mittels Primer-Extension
überprüft. Die dazu verwendeten Primer sind in Material und Methoden
aufgelistet.
Die getroffenen Annahmen konnten experimentell bestätigt werden. So
wurden kurz vor dem orfl zwei, nur durch drei Nukleotide getrennte
transkriptioneile Starts identifiziert und zwar an Position 1167 und Position
1171 (Fig. 9; Fig. 13). Diese beiden Starts wurden mit allen drei eingesetzten
Primern bestätigt. Ebenfalls gefunden wurde der transkriptioneile Start 5' von
glmM. Dieser befand sich an Position 2954, 13 Nukleotide entfernt vom
Startkodon von glmM (Fig. 9; Fig. 13).
Hingegen konnte, auch mit verschiedenen Primern, vor orfl kein
transkriptioneller Start identifiziert werden. Zudem konnte keine mögliche
Promotor- oder Shine-Dalgarno Sequenz vor dem orfl identifiziert werden.
Der orfl scheint demnach nur im Zusammenhang mit dem ganzen Operon
transkribiert zu werden. Bei der 2.2 kb grossen Bande, die bei der
Hybridisierung der Northern Blots erhalten wurde, scheint es sich daher um
ein Abbauprodukt des grösseren Transkripts zu handeln.
Weiter wurde in der Promotor Region des orfl-orfl-glmM Operons
eine für den Stressfaktor SigB spezifische Promotorsequenz identifiziert (Fig.
14) (39). Allerdings befindet sich, wenn man von diesem SigB Promotor

Resultate65
G A T C G A T C
B
Figur 13: Transkriptionsstarts. Die Transkriptionsstarts des glmM
Operon wurde mittels Primer Extension bestimmt. Die Spuren G, A, T
und C zeigen die Sequenz der entsprechenden Region des glmM
Operons. (A) zeigt den Transkriptionsstart von orfl, (B) denjenigen
von glmM. Die Transkriptionsstarts sind jeweils mit einem Stern
gekennzeichnet.

VV
TGTCGTATGACAGTGATGTAAATGATTGGTATAATGGGTATTATGGAAAAATATTCCCGGAGGAGATGTTATGGA
35
-10
^#
TGTCGTATGACAGTGATGTAAATGATTGGTATAATGGGTATTATGGAAA
-35
-10
TGTCGTATGACAGTGATGTAAATGATTGGTATAATGGGTATTATGGAAA
B
Figur
14:PromotorRegiondesorfl-orß-glmMOperons.
(A)gesamtePromotorRegiondesglmM
Operons:
v,Transkript
ions
star
tpun
kte:
I1
,Shine-Dalgarno
Sequenz;
unte
rstr
iche
n,Startkodon
ATG.
(B)
vorgeschlagene
Promotor
Region
desglmM
Operons
mit
den
beiden
Transkriptionsstartpun
kten
undSigB-ErkennungssequenzinnerhalbderPromotorRegion.

Resultate 67
ausgeht, der erste Transkriptionsstart A mitten im -10 Bereich des Promotors
und der zweite Transkriptionsstart nur zwei Nukleotide vom letzten Nukleotid
von der -10 Region entfernt. Trotzdem wurde mRNA aus sigB Mutanten und
gestressten Wildtypen isoliert, um einen ersten Eindruck einer möglichen
Wirkung des SigB Promotors zu erhalten. Die mit 6% v/v 100% EtOH
gestressten Stämme (siehe auch Kapitel 2.6.) zeigten zwar kein anderes
Bandenmuster als deren Wildtypen, aber es waren Unterschiede in den
Intensitäten der Banden festzustellen (Fig. 15). Es hat sich gezeigt, dass bei den
sigB Mutanten und den mit EtOH gestressten Zellen die 1.3 kb glmM Bande
eine grössere Intensität aufwies. Vor allem bei den sigB Mutanten war die
glmM Bande viel intensiver als beim entsprechenden Wildtyp.
Die Zellwand von der glmM Mutante PG108 unterschied sich nicht
wesentlich von der Zellwand des Wildtyps BB270, ausser durch eine geringe
Menge an durch ein D-Ala substituierte Interpeptidkette. Wurde jedoch die
glmM Mutante in Anwesenheit von Erythromycin wachsen gelassen, erschienen
Monomer- und Dimer-Muropeptide, die auf eine erhöhte Endopeptidase-
Aktivität schliessen lassen (Roos, persönliche Mitteilung). Es schien so, als
würde Erythromycin bei PG108 in der Lage sein, regulatorisch auf eine
Endopeptidase zu wirken. Um zu testen, ob Erythromycin einen Einfluss auf
die Transkription des glmM Operons hat, wurden daher PG108 und PG100 in
Erythromycin wachsen gelassen und daraus die mRNA isoliert. Es konnten
allerdings keine Veränderungen der Bandenmuster festgestellt werden (Fig.
15), so dass angenommen wird, dass Erythromycin keinen Einfluss auf die
Transkription des glmM Operons hat.
3.1.4. Komplementation der glmM Mutation
Die Insertionsinaktivierung von glmM führte zu einer Reduktion der
Grundresistenz. Um zu überprüfen, ob die Abnahme der Grundresistenz auf
die Transposon-Insertion zurückzuführen ist, wurde die glmM Mutante PG108
mit dem Plasmid pPG76 (argl-orfl-orß-glmM) und dem Plasmid pHK4176

abcdefghij
kl
m
Figu
r15:NorthernBlotverschiedenerglmM-und
s/gZ
?-Mu
tant
enhy
brid
isie
rtmitglmM
internenFragment
(Fig
.8).
a,PG100
(Em);
b,PG100;
c,BB591
(Em);
d,BB591;
e,
PG108
(Em);
f,PGJ08;
g,COLAsigB;
h,COL
(EtOH);
i,COL;
j,BB270AsigB;
k,BB270
(EtOH);
1,BB270;m,PG79.(EtOH):ab
einerbestimmtenOD600mit6%
v/v100%EtOH
bis
zurgewünschten
OD600
gest
ress
t;(E
m):
Zellen
inLB
mit20pg/ml
Erythromycin
gewachsen.

Resultate 69
(glmM) in trans komplementiert, das heisst, es wurde untersucht, ob dank dem
transduzierten Plasmid wieder MHK-Werte des Wildtyps BB270 erreicht
werden können.
Komplementation in trans
Die Komplementation der glmM Mutation wurde sowohl mit dem
Plasmid pHK4176 (glmM), als auch mit dem Plasmid pPG76 (argl-orfl-orß-
glmM) durchgeführt.
Die Komplementation von PG108 mit pPG76 ergab den Stamm PG79,
der dieselbe Resistenz gegen ß-Laktame sowie Teicoplanin aufwies wie der
parentale Stamm BB270 (Tab. 7). Hingegen konnte in PG108 mit dem Plasmid
pHK4176 (Stamm PG231) nur die ß-Laktam Resistenz nicht aber die
Teicoplanin-Resistenz wieder hergestellt werden. Auch im Suppressor Stamm
PG100 hatte das Plasmid pHK4176 (Stamm PG232) keinen Einfluss auf die
Teicoplanin-Resistenz. Dies führte zur Vermutung, dass möglicherweise orfl
und/oder orß einen Einfluss auf die Teicoplanin Resistenz ausüben könnten.
Phänotypische Komplementation
Die Syntheseschritte, die in E. coli zu GlcNAc-1-P, einem
zytoplasmatischen Vorläufer der Peptidoglykansynthese, führen, sind vor
kurzem veröffentlicht worden (Fig. 16) (90). Aufgrund dieser Daten wurde
versucht glmM durch externe Zugabe eines Vorläufers von GlcNAc-1-P
phänotypisch zu komplementieren. Der Einfluss von Mannosamin,
Glukosamin, Glukose, Glukose-1,6-di-Phosphat oder Glukose-1-Phosphat auf
die MHK von Methicillin wurde bestimmt.
Nur Glukosamin konnte die Methicillin-Resistenz von Stamm PG108 auf
die Werte des Ausgangsstammes BB270 erhöhen (Tab. 8). Die anderen
getesteten Zucker hatten keinen Einfluss auf die Methicillin-Resistenz und die

NANA
ManNAc-
GlcNAc-
GlcN-
NanT NanA
NANA ManNAc + Pyruvat
NanK
ManNAc-6-P
NanE
GlcNAc-6-P,
NagA*
NagB
GlcN-l-P
GlmU
tGlcNAc-1-P
GlmU
Glc-6-P
aussen mnen UDP-GlcNAc
Figur 16: Synthese von GlcNAc-1-P in E. coli nach Piumbridge et al. (90),verändert. Ausgezogene Pfeile: in E. coli identifizierte Stoffwechselschritte;
gestrichelte Pfeile: postulierte Wege in S. aureus; NANA: N-
Acetylneuraminsäure; ManNAc: N-Acetylmannosamin; GlcN: Glukosamin;
GlcNAc: N-Acetylglukosamin; Fru: Fruktose; Glc: Glukose. NanT: NANA-
Transporter; NanA: NANA-Lyase; NanK: ManNAc Kinase; ManXYZ:
Phosphotransferase System (PTS); NanE: Epimerase; NagG: GlcNAc
spezifischer PTS Transporter; NagA: Deacetylase; NagB: Deaminase;
GlmS: Amidotransferase; GlmM: Phosphoglukosamin-Mutase; GlmU:
bifunktionelles Enzym, Acetyltransferase und UDP-Pyrophosphorylase.

Tabelle
8:EinflussverschiedenerGlcNAc-1-PVorläuferaufdieMHK
von
MethicillinundTeicoplanin
inverschiedenen
Stämmenim
Mikrodilutionsverfahren.
MethicillinMHK
pg/ml
TeicoplaninMHK
pg/ml
Stämme
+10mM
Glukose
+10mM
Glukosamin
+5mM
Glukose
1,6-
di-P
+10mM
Glukose-1-P
+5mM
Mannosamin
+10mM
Glukosamin
PG108
42
>256
0.5
40.5
0.032
0.032
BB270
128
128
>256
232
41
0.5
Ha56
22
80.5
20.5
0.032
0.032
BB1395
44
256
44
10.25
0.25
Ha57
32
32
256
4128
80.25
0.125
BB1399
64
64
16
0.5
128
64
0.5
0.25
COL
>256
>256
>256
32
256
64
1.5
1
COL-TSlll
0.5
132
<0.25
20.5
0.063
0.25
P:Phosphat

Resultate 72
phosphorylierten Zucker hatten sogar eine hemmende Wirkung auf das
Wachstum und die MHK-Werte.
Bei Teicoplanin war mit keiner der Vorläufersubstanzen eine
Veränderung in der Empfindlichkeit gegen dieses Glykopeptid festzustellen
(Tab. 8).
3.1.5. Wachstumsverhalten
Nachdem bekannt war, dass Glukosamin die Methicillin-Resistenz in
glmM Mutanten erhöhte, wurde dessen Einfluss auf die
Wachstumsgeschwindigkeit untersucht. Interessanterweise hatte die Zugabe von
10 mM Glukosamin in der exponentiellen Wachstumsphase bei allen getesteten
Stämmen einen hemmenden Einfluss, wie dies bei den anderen getesteten
Substanzen bereits bei der MHK Bestimmung festgestellt werden konnte. Dies
drückte sich auch in den höheren Generationszeiten aus. So betrug die
Generationszeit von PG108 71 min, währenddessen die Generationszeit
desselben Stammes bei Zugabe von Glukosamin auf 81 min anwuchs. Noch
deutlicher fiel dieser hemmende Einfluss bei PG100 aus: ohne Glukosamin
betrug die Generationszeit rund 94 min und mit Glukosamin etwa 120 min.
Auch BB270 war von einer Generationszeitzunahme unter Glukosamineinfluss
betroffen: hier stieg die Zeit von 67 auf 85 min. Dafür erreichten die mit
Glukosamin wachsenden Zellen die höhere OD600als diejenigen Zellen, die ohne
Glukosamin wuchsen (Fig. 17). Diese erhöhte OD könnte auf die erhöhte C-
Quelle durch die externe Glukosaminzugabe zurückzuführen sein, während der
hemmende Einfluss der externen Glukosaminzugabe auf das Wachstum auf die
Katabolitrepression zurückzuführen ist.
3.1.6. Proteinanalysen
MRSA Stämme besitzen das Strukturgen mecA, das für ein zusätzliches,
hochmolekulares PBP, das PBP2', kodiert, das für die Methicillin-Resistenz

min
mm
mm
B
Figur
17:Wachstumskurvenmit/ohne
10mM
Glukosaminim
LB-Medium.D,
repr
äsen
tier
tLB-
MediumohneGlukosamin;A,
repräsentiertLB-MediummitGlukosamin.A,BB270;B,PG108;
C,
PG100.

Resultate 74
verantwortlich ist. PBP-Profile der verschiedenen Stämme zeigten, dass glmM
keinen Einfluss auf die PBP2' Produktion hatte. Die typische PBP2' Bande war
sowohl beim Wildtyp BB270 als auch bei der glmM Mutante PG108 und der
Suppressor Mutante PG100 deutlich zu sehen (Fig. 18B). Im Vergleich dazu
fehlte diese Bande beim MSSA Stamm BB255 (Fig. 18B, Spur a). Liess man
den Wildtyp BB270 ÜN in Anwesenheit von Methicillin wachsen, so zeigte
sich, dass alle PBP ausser dem PBP2' und einem kleinen Anteil des PBP2 mit
Methicillin abgesättigt waren und daher nicht mit dem radioaktiven
(3H)Benzylpenicillin markiert werden konnten (Fig. 18B, Spur c). Das
Membranprotein Muster der Suppressor Mutante PG100 zeigte zudem eine 49
kDa grosse Bande (Fig. 18A, Spur d) von starker Intensität verglichen mit den
übrigen Stämmen. Diese 49 kDa Bande könnte möglicherweise mit der
postulierten Suppressor Mutation hmrDl zusammenhängen.
Anhand der aus den Northern Blots erhaltenen Resultate wurde davon
ausgegangen, dass PG108 und PG100 kein GlmM produzieren würden. Dies
konnte in Western Blots bestätigt werden (Fig. 19). Sowohl beim Stamm
PG108 als auch beim Suppressor Stamm PG100 konnte mit den GlmM-
Antikörpern keine Phosphoglukosamin-Mutase mehr nachgewiesen werden,
während bei den MSSA und MRSA Wildtypen und beim komplementierten
Stamm PG79 ein Signal detektiert werden konnte. Das Molekulargewicht der
GlmM Bande betrug etwa 57 kDa und war somit um 7.4 kDa grösser als die
errechnete. Bei den zusätzlichen Banden von geringerem Molekulargewicht
beim komplementierten Stamm handelt es sich wahrscheinlich um unspezifische
Produkte, da mit ungereinigten Antikörpern hybridisiert wurde.
Da eine mögliche SigB abhängige Promotorsequenz vor dem glmM
Operon identifiziert wurde (Fig. 14), wurde überprüft, ob das glmM Operon
durch einen der bekannten globalen Regulatoren beeinflusst wird. Es wurden
sowohl sigB-, sar- als auch agr- Mutanten auf ihre GlmM Expression
untersucht. Bei den aus ÜN-Kulturen gewonnen Proteinen zeigte sich zwischen
den Wildtypen und den entsprechenden Mutanten Unterschiede in der Intensität
der GlmM Bande (Fig. 20A und 20B). Wiederum war ein Unterschied
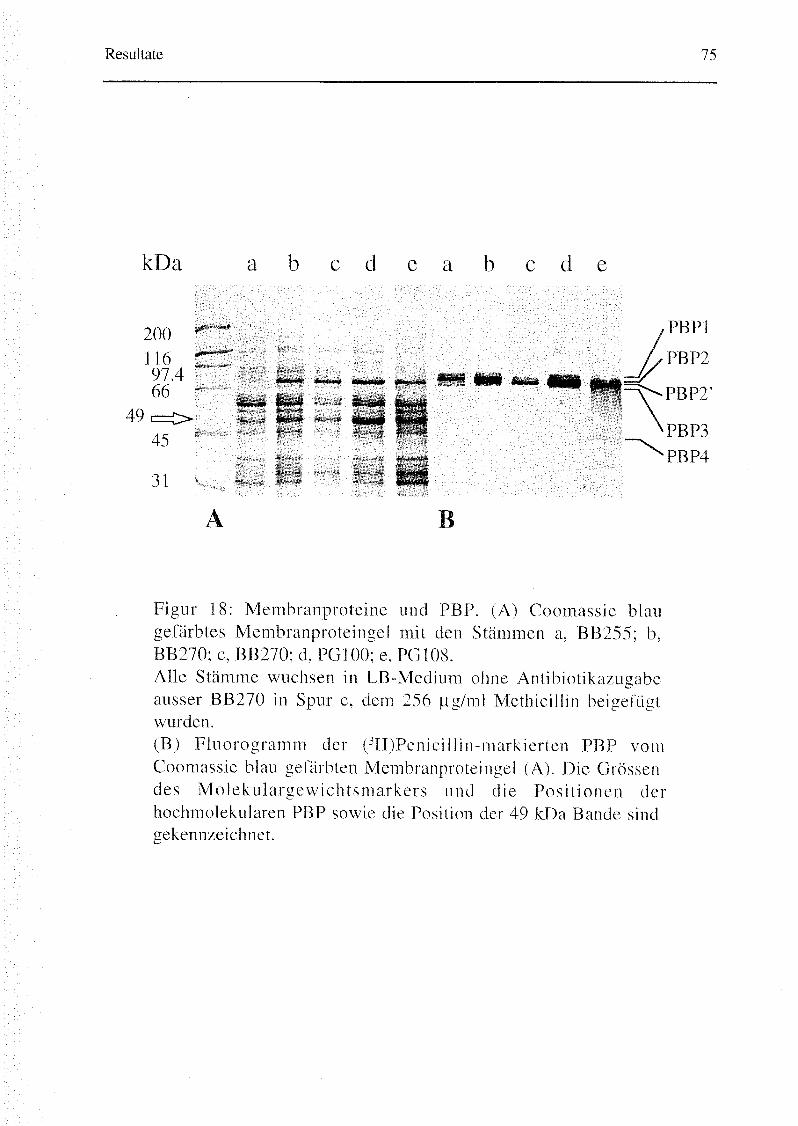
Resultate 75
kDa abcdeabcde
200^»*w!(**
A
^W-VYsfe
O
PBP3
PBP4
Figur 18: Membranproteine und PBP. (A) Coomassie blau
gefärbtes Membranproteingel mit den Stämmen a, BB255; b,
BB270; c, BB270; d, PG100; e, PG108.
Alle Stämme wuchsen in LB-Medium ohne Antibiotikazugabeausser BB270 in Spur c, dem 256 pg/nü Methicillin beigefügtwurden.
(B) Fluorogramm der (Tf)PeniciHin-markierten PBP vom
Coomassie blau gefärbten Membranproteingcl (A). Die Grössen
des Molekulargewichtsmarkers und die Positionen der
hochmolekularen PBP sowie die Position der 49 kDa Bande sind
gekennzeichnet.

Resultate 76
kDa a b c d e f
Figur 19: Nachweis der Phosphoglukosamin Mutase m verschiedenen
Stammen im Western-Blot. Zytoplasmatische Zeil-Extrakte von UN-
Kulturen wurden mit Anti-GlmM Antikörpern hybridisiert, a, PG79; b,
PG100; c, PG108; d. BB255; e. BB270, t, lekombinantes GlmM. Der
Pfeil zeigt auf GlmM

Resultate 77
a b c d e f g
*Pf| ^—y
l\
a b c d e f g
B
Figur 20: Nachweis der Phosphoglukosamin Mutase in
verschiedenen sigB- (A), agr- und «//--Mutanten (B) im Western-
Blot. Zytoplasmatische Zeil-Extrakte von ÜN-Kulturen wurden
mit Anti-GlmM Antikörpern hybridisiert. (A): a, BBIOOO; b,
BB WOOAsigB; c. COL ; d, COL AsigB; e, BB255; f, BB255A.s7##;g, PGI08. (B): a. BB1164; b, BB1163; c, BB1 166; d, BB1165; e,
BB270; f. Molekluargewichtsmarker; g, PGJ08. Pfeil zeigt auf
GlmM.

Resultate 78
zwischen dem Wildtyp und seiner sigB Mutante festzustellen, wie dies schon
bei den Northern Blots zu sehen war. Die sigB Mutante schien jeweils mehr
GlmM zu produzieren als der Wildtyp (Fig. 20A). Dieser Unterschied war vor
allem zwischen BB255 und seiner sigB Mutante zu erkennen. Bei der agrisar
Doppelmutante BB1164 schien weniger GlmM produziert zu werden als bei
der agr Mutante BB1163 (Fig. 20B). Wie relevant diese Resultate allerdings
sind lässt sich erst sagen, wenn die GlmM Expression in den verschiedenen
Stämmen über die gesamte Wachstumsphase überprüft würde. Einzig bei der
sigB Mutation scheint ein Zusammenhang zu bestehen zwischen der sigB
Mutation und der Menge an transkribierten glmM (Fig. 15) und der Menge an
produziertem GlmM.
GlmM ist in den Biosyntheseweg der Peptidoglykan-Vorläufer
involviert. Daher wurde angenommen, dass GlmM dementsprechend nur im
Zytoplasma gefunden würde. Wie Figur 21 zeigt, konnte GlmM zwar in der
Membranproteinfraktion nicht nachgewiesen werden, dafür wie erwartet in der
Zytoplasmafraktion.
3.2. Das fmtB-System
Die meisten Methicillin-Resistenz beeinflussenden Faktoren wurden
mittels Tn55I Insertions-Mutagenese und anschliessender Zunahme der
Methicillin-Empfindlichkeit identifiziert. Nicht-inhibitorische Konzentrationen
des nicht-ionischen Detergens TritonX-100 reduzieren die Oxacillin Resistenz
in MRSA ohne dabei das Wachstum, die Lebensfähigkeit oder die PBP2'
Produktion zu beeinflussen (55, 56). Der Einfluss von TritonX-100 auf die
Zunahme der Empfindlichkeit ist von Stamm zu Stamm unterschiedlich und
scheint vom genetischen Hintergrund des jeweiligen Stammes abzuhängen.
Wenn nebst der Zunahme der Empfindlichkeit gegenüber Oxacillin auch noch
die Zunahme der Empfindlichkeit gegenüber TritonX-100 als Kriterium zur

abed efghiWW
A
abed efgh
B
Figur 21: Nachweis der Phosphoglukosamin Mutase in
verschiedenen glmM- und ////fB-Mutanten im Western-Blot.
Entsprechende Zeil-Extrakte aus ÜN-Kulturen wurden mit
Anti-GlmM Antikörpern hybridisiert. (A) zytoplasmatischeFraktion: a, Ha56; b, Ha57; c. Marker; d, COL; e, COL-TS111 ;
f, BB270; g, PG108; h. PG100; i, PG79.
(B) Membranprotein Fraktion: a, Ha56; b, Ha57; c, COL; d,
COL-TS111; e. BB270; f, PG108; g. PG100; h, PG79.
Der Pfeil zeigt auf GlmM.

Resultate 80
Identifikation von Mutanten eingesetzt wird, können andere Arten von
Faktoren gefunden werden, die die Resistenz beeinflussen können.
Distal von glmM liegt fmtB, das von Komatsuzawa identifiziert, kloniert
und sequenziert wurde. fmtB kodiert für ein 263 kDa grosses Peptidoglykan-
assoziiertes Protein. Es hat die für ein Peptidoglykan-assoziiertes Protein
typische positive Ladung im C-Terminus, gefolgt von einer Signalsequenz, dem
Zellwand-sortierenden LPXTG-Motif, einer im zentralen Bereich von FmtB
gelegenen 17 fachen Tandem-Repeat-Region von 75 AS sowie einem 687 AS
langen N-terminalen Teil (Fig. 22). Komatsuzawa konnte proximal vom
Startkodon von fintB eine mögliche Shine-Dalgarno Sequenz identifizieren und
mittels Primer-Extension 14 bp proximal zum Startkodon den
Transkriptionsstart bestimmen.
3.2.1. Phänotypische Charakterisierung der Tn55I Insertion in
fmtB
Komatsuzawa konnte zeigen, dass eine Tn557 Insertion in fmtB im
homogen hoch Methicillin-resistenten Stamm COL eine starke Reduktion der
Oxacillin-Resistenz (Tab. 9) und eine Reduktion der Resistenz gegenüber
TritonX-100 bewirkte (Komatsuzawa et al., Manuskript eingereicht).
Rückkreuzungen in zwei genetisch unterschiedliche aber ebenfalls homogen
hoch resistente MRSA bestätigten die starke Reduktion der Oxacillin-Resistenz
durch die Insertion fmtB::Tn551. In MSSA Stämmen hatte die Tn557 Insertion
in fmtB weder einen Einfluss auf die Oxacillin- noch auf die Triton-XIOO
Empfindlichkeit. Die Transduktion von fintB::Tn3'51 in BB270 ergab zwei
Typen von Transduktanten. Transduktanten vom Typ I, repräsentiert durch die
beiden Stämme Ha57 und BB1399, waren Methiciliin-resistent und erschienen
bereits nach zwei Tagen, während Transduktanten vom Typ II, repräsentiert
durch die beiden Stämme Ha56 und BB1395, Methicillin-empfindlich waren
und erst nach vier Tagen Inkubation erschienen. Das Verhältnis Typl : TypII
betrug 1:10 nach sechs Tagen Inkubation. Transduktanten vom Typ I zeigten

Tn55i
glmM
nght R]
-R17
LPXTG
EfmtB
PB
CP
PEC
P
1kb
Figur
22:
RestriktionskartederfmtB
Region.
B,BamHl;
C,
Clal
;E,EcoRV;
P,Pstl.Die
Positionund
Orientierung
desTn551
istangegeben.R1-R17:
Inverted
Repe
ats.
LPXTG:
Zellwand-sortierendeMotifimC-terminalenBereich..Die
fürdenNorthernBlotverwendete
internefmtBProbe
istalsdo
ppel
teLinieangegeben.

Resultate 82
Tabelle 9: Minimale Hemmkonzentrationen von Oxacillin und Teicoplanin
gegen verschiedene ymtö-Mutanten.
Stämme MHK(ag/ml)OX TP
COL >256Jmi
COL-TSlll 0.13 0.19
COL-TSlll
pHK4357 (fmtBvon COL) 0.13* nb
COL-TSlll
pHK4069 (fmtBvon KSA8)COL-TSlll
pHK4176 (glmM)COL-TSlll
pHK4251(glmM+fmtB)
0.13*
128*
256*
nb
nb
nb
RN450 1 1.5
RN450-TS111 0.75 1.5
BB270 12 1
Ha56 0.38 0.38
BB1399 0.75 0.38
Ha57 12 1.5
BB1395 12 1
OX: Oxacillin; TP: Teicoplanin; *: Werte von Komatsuzawa; nb: nichtbestimmt

Resultate 83
dicke, fette Kolonien, wohingegen die Transduktanten vom Typ II schwach
wuchsen und nur kleine Kolonien bildeten. Beide Typen von Transduktanten
waren Hämolyse positiv. Obwohl sich die zwei Typen in ihrer Resistenz
gegenüber Methicillin unterschiedlich verhielten, enthielten sie die
fmtB::Tn551 Insertion wie durch Southern Blots bestätigt werden konnte
(Daten nicht gezeigt).
3.2.2. Komplementation der fmtB Mutation
Komplementation in trans
Interessanterweise konnte Komatsuzawa die fmtB Mutation weder mit
fmtB aus COL noch mit fmtB aus KSA8, einem weiteren MRSA,
komplementieren. Die Oxacillin Empfindlichkeit war dieselbe wie in der
ursprünglichen fintB Mutante COL-TSlll (Tab. 9). D'à fmtB nur wenige
hundert Nukleotide distal vom glmM Operon liegt, wurde vermutet, dass die
Tn55i Insertion möglicherweise einen polaren Effekt auf das gesamte glmM
Operon oder zumindest auf glmM ausüben könnte und die Resistenz gegen
Oxacillin aus diesem Grund herabgesetzt wurde. Die Komplementation von
fintB: :Tn551 mit glmM alleine führte zu einem 1000 fachen Anstieg der MHK
verglichen mit der Mutante (Tab. 9). Wurde fmtB::Tn551 mit glmM plus fmtB
komplementiert, wurden fast die MHK Werte des Wildtyps erreicht.
Phänotypische Komplementation
Da die Zugabe des Zellwandvorläufers Glukosamin bei PG108 die
Resistenz wiederherstellen konnte und glmM in trans die fmtB Mutation
komplementieren konnte, wurde der Einfluss von Glukosamin und N-
Acetylglukosamin auf die Methicillin-Resistenz bei fmtB Mutanten untersucht.
Die Zugabe sowohl von Glukosamin als auch von N-Acetylglukosamin

Resultate 84
erhöhten die Resistenz (Tab. 10), wobei die ursprüngliche Oxacillin-Resistenz
des Wildtyps mit beiden Zellwandvorläufern nicht ganz erreicht wurde.
3.2.3. Wachstumsverhalten
Wie bei den glmM Mutanten wurde auch bei den fmtB Mutanten der
Einfluss von Glukosamin auf das Wachstum untersucht. Während die
Generationszeit bei COL um die 66 min war, war diejenige von COL-TSlll
rund 73 Minuten. Interessanterweise hatte Glukosamin eine eher hemmende
Wirkung auf die Verdopplungszeit, auch wenn der Einfluss nicht so stark
ausfiel wie bei PG100. Unter Einfluss von Glukosamin erreichten die
Generationszeiten einen Wert von 83 min bei COL bzw. 87 min bei seiner
entsprechenden fmtB Mutante (Fig. 23). Bei den von BB270 abstammenden
fmtB Transduktanten war die Generationszeit in etwa gleich wie bei ihrem
Ausgangsstamm. Sie betrug bei der TypI/mfß Transduktante Ha57 71 min und
bei der TypII fmtB Transduktante 68 min (Fig. 24). Offenbar hatte die fmtB
Mutation in BB270 nur einen geringen Einfluss auf die Generationszeit. Aber
auch bei den beiden Typen von BB270 ^«^-Transduktanten hatte die Zugabe
von 10 mM Glukosamin in der exponentiellen Wachstumsphase einen
hemmenden Einfluss. Dafür erreichten die Ansätze mit Glukosamin ebenfalls
höhere Endkonzentrationen.
3.2.4. Transkription der fmtB Region
Die DNA-Sequenz der Region zwischen glmM und fmtB Hess vermuten,
dass die beiden ORF vermutlich unabhängig voneinander transkribiert werden.
Da glmM die fmtB Mutation in trans komplementieren konnte, wurde
untersucht ob diese Tn55i Insertion möglicherweise einen Einfluss auf die
glmM Transkription hat. Die Transkription war jedoch identisch im Wildtyp
und in der fmtB Mutante (Fig. 25). Es konnte in keinem Fall ein Transkript

Resultate85
Tabelle 10: Einfluss von N-Acetylglukosamin und Glukosamin auf die MHK
von Oxacillin (Werte erhalten von Komatsuzawa).
Oxacillin MHK
pg/ml
Stämme +10mMN-
Acetylglukosamin
+10 mM
Glukosamin
COL
COL,fmtB::Tn551
COL glmM::Tn551
512
0.25
4
512
64
64
1024
32
64

min
A
mm
B
Figur
23:Wachstumskurvenmit/ohne
10mM
Glukosaminim
LB-Medium.,
repr
äsen
tier
tLB-
MediumohneGlukosamin;A,
repräsentiertLB-MediummitGlukosamin.A,COL;B,COL-TSl
11.

mm
B
mm
mm
Figur24:Wachstumskurvenmit/ohne
10mM
Glukosaminim
LB-Medium.,
repr
äsen
tier
tLB-
MediumohneGlukosamin;A,
repräsentiertLB-MediummitGlukosamin.A,BB270;B,Ha57;
C,
BB1399.

Resultate 88
kb abed
Figur 25: Transkription von glmM. Northern Blot der fmtBMutanten und ihren entsprechenden Wildtypen hybridisiert mit
einem glmM internem Fragment (Fig. 8). a, COL; b, COL
fmtB::Tx\551; c, RN450; d, RN450,/mfi::Tn557. Die Grössen der
einzelnen Banden sind angegben.

Resultate 89
grösser als 3.2 kb nachgewiesen werden, was bei einer Kotranskription von
glmM undfmtB der Fall sein müsste.
Die fmtB Transkription wurde mittels Northern Blots und einer Probe
aus dem N-terminalen Bereich von fmtB (Fig. 22) analysiert. Da nur diffuse
Abbauprodukte und keine Transkripte von der zu erwartenden Grösse
nachgewiesen werden konnten, ist nicht bekannt, ob fmtB Teil eines grösseren
Operons ist oder allenfalls monozistronisch transkribiert wird (Daten nicht
gezeigt).
3.2.5. Proteinanalysen
Da die Komplementation von fintB;:Tn551 mit fmtB keine Zunahme der
Oxacillin-Resistenz zur Folge hatte, mit glmM hingegen erfolgreich war,
drängte sich die Frage auf, ob in den fintB Mutanten eventuell die GlmM
Produktion durch Tn557 Insertion beeinflusst worden ist. Western Blots mit
entsprechenden GlmM-Antikörpern zeigten, dass immer noch GlmM
produziert wurde, unabhängig davon, ob das fmtB unterbrochen war oder
nicht (Fig. 21). Die reduzierte Oxacillin-Resistenz in fmtB Mutanten schien
daher nicht direkt mit der Menge von produziertem GlmM
zusammenzuhängen.
3.3. Das femX-System (fmhB-System)
3.3.1. Identifizierung des fem Faktors femX (fmhB)
FemA und FemB sind in die Synthese der Pentaglycin Seitenkette
involviert. Sie kanalisieren oder kontrollieren den Einbau von Gly2-Gly3
(FemA) und Gly4-Gly5 (FemB). Ein dritter Faktor, FemX, der für den Einbau
des ersten Glycins verantwortlich ist wurde bereits postuliert (57). Es wurde
vermutet, dass FemX Sequenzähnlichkeit mit FemA oder FemB haben könnte

Resultate 90
und dass femX für das Überleben der Zelle essentiell ist. Eine potentielle
Sequenz fmhB, die femX entsprechen könnte, wurde durch Tschierske
beschrieben (110).
Anhand der zum Teil bekannten Sequenz von fmhB wurde eine Probe
hergestellt (Fig. 26) um das fmhB Allel und die umliegende Region aus dem
Stamm BB270 zu klonieren. Ein 4 kb grosses EcoRV Fragment wurde in einen
Smal geschnittenen pUC18 Vektor Moniert. Das daraus resultierende Plasmid
pPGl.l mit einem Insert von ungefähr 4 kb Grösse wurde vollständig
durchsequenziert.
Analysieren der Sequenz
Der ORF des fmhB beginnt bei Nukleotid 1 und endet bei Nukleotid
1263 (Fig. 27). fmhB kodiert für ein 421 AS grosses Protein mit einem
errechneten Molekulargewicht von 46.3 kDa. FmhB hat eine Identität von 27%
zu FemA und eine Identität von 26% zu FemB (110). Distal zu fmhB folgte ein
Abschnitt von sieben Thymidin-Resten mit vorgestelltem Inverted Repeat von 9
Nukleotiden, der für einen Faktor-unabhängigen RNA-Polymerase Terminator
charakteristisch ist (17). DG für diesen Terminator beträgt -10.2 kcal/mol.
In einem Abstand von 173 Nukleotiden 3' von fmhB konnte das 5' Ende
eines ORFs identifiziert werden, der über den Monierten Bereich
hinausreichte. Er startet bei Nukleotid 1437 und bricht aufgrund der Grösse
des klonierten Fragments bei Nukleotid 3858 ab. Die Grösse des partiellen
Proteins beträgt 807 AS und hat ein Molekulargewicht von 88.7 kDa.
Datenbankvergleiche des partiellen Proteins ergaben die grösste
Homologie mit YerP, einem Protein von B. subtilis, welches Ähnlichkeit zu
einem Akriflavin-Resistenz Protein hat. Zu diesem YerP wurde eine Identität
von 42% und eine Ähnlichkeit von 63% gefunden.

fmhB
t/—
EBC
BHA
CC
1kb
Figur
26:
RestriktionskartederMoniertenfmhB
Region.Die
offenen
Leseraster
sindmit
Pfeilenangegeben.DieMeinennummerierten
Pfeilegeben
Orientierung
und
Positionder
PCR
Primer,mitdenen
dieProbe
hergestelltwurde
an.DieProbe
istdurchdieDoppellinie
angegeben.A,AccI;B,BamHl;C,
Clal
;E,EcoKV;H,Hindlll.

Resultate 92
ATGGAAAAGATGCATATCACTAATCAGGAACATGACGCATTTGTTAAATCCCACCCAAAT]_ ******
gg
1MEKMHITNQEHDAFVKSHPN 20
GGAGATTTATTACAATTAACGAAATGGTCAGAAACAAAGAAATTAACTGGATGGTACGCG61 * * * * * * 120
21GDLLQLTKWSETKKLTGWYA 40
CGAAGAATCGCTGTAGGTCGTGACGGTGAAGTTCAGGGTGTTGCGCAGTTACTTTTTAAA
121 ****** !80
41RRIAVGRDGEVQGVAQLLFK 60
AAAGTACCTAAATTACCTTATACGCTATGTTATATTTCGCGTGGTTTTGTTGTTGATTAT
181 ****** 240
61KVPKLPYTLCYISRGFVVDY 80
AGTAATAAAGAAGCGTTAAATGCATTGTTAGACAGTGCAAAAGAAATTGCTAAAGCTGAG
241 ****** 300
81SNKEALNALLDSAKEIAKAE 100
AAAGCGTATGCAATTAAAATCGATCCTGATGTTGAAGTTGATAAAGGTACAGATGCTTTG
301 ****** 350
101 KAYAIKIDPDVEVDKGTDAL 120
CAAAATTTGAAAGCGCTTGGTTTTAAACATAAAGGATTTAAAGAAGGTTTATCAAAAGAC361 * * * * * * 420
121 QNLKALGFKHKGFKEGLSKD 140
TACATCCAACCACGTATGACTATGATTACACCAATTGATAAAAATGATGATGAGTTATTA421 ****** 480
141 YIQPRMTMITPIDKNDDELL 160
AATAGTTTTGAACGCCGAAATCGTTCAAAAGTGCGCTTGGCTTTAAAGCGAGGTACGACA481 * * * * * * 540
161 NSFERRNRSKVRLALKRGTT 180
GTAGAACGATCTGATAGAGAAGGTTTAAAAACATTTGCTGAGTTAATGAAAATCACTGGG541 ****** 600
181 VERSDREGLKTFAELMKITG 200
GAACGCGATGGCTTCTTAACGCGTGATATTAGTTACTTTGAAAATATTTATGATGCGTTG601 ****** 660
201 ERDGFLTRDI SYFENIYDAL 220

Resultate 93
CATGAAGATGGAGATGCTGAACTATTTTTAGTAAAGTTGGATCCAAAAGAAAATATAGCG661 ****** 720
221 HEDGDAELFLVKLDPKENIA 240
AAAGTAAATCAAGAATTGAATGAACTTCATGCCGAAATTGCTAAATGGCAGCAGAAGATG721 ****** 780
241 KVNQELNELHAEIAKWQQKM 260
AAAACATCTGAAAAGCAAGCTAAAAAAGCGCAAAATATGATTAATGATGCGCAAAATAAA
781 ****** 840
261 KTSEKQAKKAQNMINDAQNK 280
ATTGCTAAAAATGAAGATTTAAAACGAGACCTAGAAGCTTTAGAAAAGGAACATCCTGAA
841 ****** goo
281 IAKNEDLKRDLEALEKEHPE 300
GGTATTTATCTTTCTGGTGCACTATTAATGTTTGCTGGCTCAAAATCATATTACTTATAT
901 ****** 960
301 GIYLSGALLMFAGSKSYYLY 320
GGTGCGTCTTCTAATGAATTTAGAGATTTTTTACCAAATCATCATATGCAGTATACGATG961 ******102Q
321 GASSNEFRDFLPNHHMQYTM 340
ATGAAGTATGCACGTGAACATGGTGCAACAACTTACGATTTCGGTGGTACAGATAATGAT1021 ******1080
341 MKYAREHGATTYDFGGTDND 360
CCAGATAAAGACTCAGAACATTATGGATTATGGGCATTTAAAAAAGTGTGGGGAACATAC
1081 ***** li40
361PDKDSEHYGLWAFKKVWGTY 380
TTAAGTGAAAAGATTGGTGAATTTGATTATGTATTGAATCAGCCATTGTACCAATTAATT1141 ******1200
381 LSEKIGEFDYVLNQPLYQLI 400
GAGCAAGTTAAACCGCGTTTAACAAAAGCTAAAATTAAAATATCTCGTAAATTAAAACGA1201 ******1260
401 EQVKPRLTKAKIKISRKLKR 420
AAATAGATTAACGACTGAAATCTGAACGCTCATAAGACTGTCATTTGCGTTCAGATTTTT1261 ******j_320
421 K *

Resultate 94
TTACACAATATAGAATGGTTGAGTAAAATATTTTTGAATATAGTGAAAGAGGGGGAAGTA
1321 ******1380
CTGTGATAAAAAAGCTATTACAATTTTCTTTAGGGAATAAGTTTGCTATCTTTTTAATGG
1381 * * * SD * * * 1440
MV 2
TTGTTTTAGTTGTCTTGGGCGGTGTATATGCGAGTGCTAAATTGAAATTAGAATTACTAC
1441 ****** 1500
3 VLVVLGGVYASAKLKLELLP 22
CAAATGTACAAAATCCAGTTATTTCAGTTACAACAACAATGCCGGGTGCAACGCCACAAA
1501 ******1560
23 NVQNPVISVTTTMPGATPQS 42
GTACCCAAGATGAAATAAGTAGTAAAATTGACAATCAAGTAAGATCATTGGCATATGTGA
1561 ******1620
43 TQDEISSKIDNQVRSLAYVK 62
AAAATGTTAAAACGCAATCCATACAAAATGCTTCAATTGTAACAGTTGAATATGAAAATA
1621 ******16go
63 NVKTQSIQNASIVTVEYENN 82
ATACAGATATGGATAAAGCAGAAGAACAGCTTAAAAAAGAAATCGATAAAATTAAATTTA
1681 ******1740
83 TDMDKAEEQLKKEIDKIKFK 102
AAGATGAAGTTGGTCAACCAGAATTAAGACGTAATTCGATGGATGCTTTTCCGGTTTTAG
1741 ******180o
103 DEVGQPELRRNSMDAFPVLA 122
CATATTCATTTTCAAATAAAGAGAATGACTTGAAAAAAGTAACGAAAGTACTGAATGAAC
1801 ******1860
123 YSFSNKËNDLKKVTKVLNEQ142
AATTAATACCAAAATTGCAAACGGTAGATGGTGTGCAAAATGCGCAATTAAATGGGCAGA
1861 ******192o
143 LIPKLQTVDGVQNAQLNGQT162
CGAACCGTGAAATCACCCTTAAATTTAAGCAAAATGAACTTGAAAAATATGGGTTGACTG
1921 ******193o
163 NREITLKFKQNELEKYGLTA182
CTGATGATGTAGAAAACTATCTAAAAACGGCAACAAGAACAACGCCACTTGGATTGTTCC
1981 ******2040
183 DDVENYLKTATRTTPLGLFQ 202

Resultate
AATTTGGTGATAAAGATAAATCAATTGTTGTTGATGGTCAATATCAATCTGTTGATGCTT
2041 ******2100
203 FGDKDKSIVVDGQYQSVDAF222
TTAAAAACATAAATATTCCATTAACGTTGGCAGGAGGACAAGGGCAATCTCAATCCCAAA
2101 ******2160
223 KNINIPLTLAGGQGQSQSQS 242
GTGACAATAAACAAAATTCAGCCATGTCAGACGTTAATCAGGCATCACCACAGCAAAATT
2161 ******2220
243 DNKQNSAMSDVNQASPQQNS 262
CAAAAGCGTCAGCACCAAATAATATAAGTGGTATGCCGACAGCGAAACTAGGAGATTTAG
2221 ******2280
263 KASAPNNISGMPTAKLGDLA282
CTGATATTACAGTTGGTGATGTGCGTACTTCTATTTCTAAAACGAATGGAAAGGATGCGG
2281 ******2340
283 DITVGDVRTSISKTNGKDAV302
TTAATCTACAAATAACTAAAGCTCAAGATGCCAATACAGTTCAAGTAGCCAAAGATGTAC
2341 ******2400
303 NLQ ITKAQDANTVQVAKDVQ 322
AACGTAAAATTGATACATTCGTTGATGAAAATAAAGATTTTAATGTCACAAAAACAATGG
2401 ******2460
323 RKIDTFVDENKDFNVTKTMD 342
ATACTGCAAAGCCTGTTGAGAAATCACTTTATACGATGGTTGAAAAAGCATCATTAGGTA
2461 ******2520
343 TAKPVEKSLYTMVEKASLGT 362
CAATCGTGGCAATTATAGTTATTTTGCTGTTTTTAAGAAACATTCGTACGACGGCAATTT
2521 ******2580
3 63 IVAIIVILLFLRNIRTTAIS382
CTATTATATCGATTCCGTTATCACTTCTTATGGCGCTTATTGCTCTGAAATTGAGTGATG
2581 ******2640
383 IISIPLSLLMALIALKLSDV 402
TTTCATTGAATATACTAACGTTAGGTGCATTAACAGTAGCGATTGGACGTGTGATAGACG
2641 ******2700
403 SLNILTLGALTVAIGRVIDD 442

Resultate 96
ATTCGATTGTAGTTGTTGAAAATATTTATCGACGCTTAACAGATTCAGAAGAACAACTAA
2701 ******2760
443 SIVVVENIYRRLTDSEEQLK462
AAGGTGAAAATTTAATTATCAGTGCGACAACTGAAGTATTTAAACCAATAATGTCATCGA
2761 ******2820
463 GENLIISATTEVFKPIMSST 482
CACTAGTTACTATTATCGTCTTCTTACCACTTGTGTTTGTATCAGGTTCAGTAGGCGAAA
2821 ******2880
483 LVTI IVFLPLVFVSGSVGEM 502
TGTTTAGACCTTTTGCATTGGCTATTGCATTTAGTTTATTAGCATCGTTATTAGTGTCAA
2881 ******2940
503 FRPFALAIAFSLLASLLVSI522
TTACACTCGTTCCAGCGTTGGCAGCTACACTATTTAAAAAAGGCGTTAAACGTCGTAATA
2941 ******3000
523 TLVPALAATLPKKGVKRRNK542
AACAACATCAAGAAGGATTAGGTGTTGTTAGTACAACTTATAAAAAAGTATTACATTGGT
3001 ******3060
543 QHQEGLGVVSTTYKKVLHWS 562
CATTAAATCATAAGTGGATTGTAATTATATTAAGTACATTAATTTTGGTTGCAACTATTG
3061 ******3120
563 LNHKWIVI ILSTLILVATIV 582
TATTTGGAGGACCGAGACTAGGCACTAGCTTTATTTCAGCAGGTGACGATAAATTTTTAG
3121 ******3180
583 FGGPRLGTSFISAGDDKFLA 602
CTATTACTTATACACCGAAGCCTGGTGAAACGGAGCAAGCAGTGTTGAATCATGCGAAAG3181 ******3240
603 ITYTPKPGETEQAVLNHAKD 622
ATGTTGAAAAATATTTAAAACAGAAAAAGCATGTAAAAACAATTCAATACTCAGTTGGCG3241 ******33oo
623 VEKYLKQKKHVKTIQYSVGG 642
GTAGTAGTCCAGTAGATCCAACGGGTAGTACAAATAGTATGGCAATCATGGTTGAATATG
3301 ******3360
643 SSPVDPTGSTNSMAIMVEYD662

Resultate 97
ATAATGACACGCCTAATTTTGATGTAGAAGCGGATAAGGTTATTAAACATGCAGATGGCT
3361 ******3420
663 NDTPNFDVEADKVIKHADGF682
TTAAACATCCTGGAGAGTGGAAAAATCAAGATTTAGGAACAGGTGCAGGTAATAAATCTG
3421 ******3480
683 KHPGEWKNQDLGTGAGNKSV702
TAGAGGTTACTGTAAAAGGTCCATCAATGGATGCCATAAAATCAACTGTAAAAGATATTG
3481 ******3540
703 EVTVKGPSMDAIKSTVKDIE 722
AACAGAAAATGAAACAGGTTAAAGGACTAGCCAATGTCAAATCTGATTTATCGCAAACAT
3541 ******36oo
723 QKMKQVKGLANVKSDLSQTY 742
ATGATCAGTATGAAATTAAAGTCGATCAAAATAAAGCGGCAGAAAATGGTATTTCTGCAA
3601 ******3660
743 DQYEIKVDQNKAAENGISAS 762
GTCAACTTGCAATGCACTTGAATGAAAACTTACCAGAAAAAACAGTTACGACTGTTAAAG
3661 ******372Q
763 QLAMHLNENLPEKTVTTVKE 782
AAAATGGTAAAACTGTTGATGTTAAAGTCAAACAAAATAAGCAAACAGACTGGTCAGAAG
3721 ******3780
783 NGKTVDVKVKQNKQTDWSED 802
ATAAGTTGAATAATATAACTTTGAAAAAGCCGACTGGTGGTACGATTAAATTGGGAGATG
3781 ******3840
803 KLNNITLKKPTGGTIKLGDG 822
GGTACCGAGCTCGAATTC
3841 *
3858
823 Y R A R I 827
Figur 27: Sequenz von fmhB und dem sequenzierten Teil des proximalen orf
Die möglichen Promotorsequenzen sind fettgedruckt und unterstrichen, Shine-
Dalgarno Sequenzen sind kursiv geschrieben, die Startkodons ATG sind
fettgedruckt und die Inverted Repeats sind unterstrichen.

Resultate 98
3.3.2. Northern Blot
Untersuchung der fmhB Transkription in Northern Blots mit der fmhB
internen Probe (Fig. 26) ergab drei Transkripte von 4.1, 2.9 und 1.7 kb (Fig.
28) mit zunehmender Intensität. Die kleinste Bande ist um etwa 400 Nukleotide
länger als fmhB. Interessanterweise war die Intensität der Banden bei den
entsprechenden sigB Mutanten und den durch Ethanol gestressten Stämmen
stärker als bei den Wildtypen. Dieses Phänomen konnte auch bei glmM in
Northern Blots beobachtet werden (Fig. 15).
Ob fmhB als Teil eines noch unbekannten Operons oder als
monozistronische mRNA abgelesen muss noch mit Primer-Extension überprüft
werden.
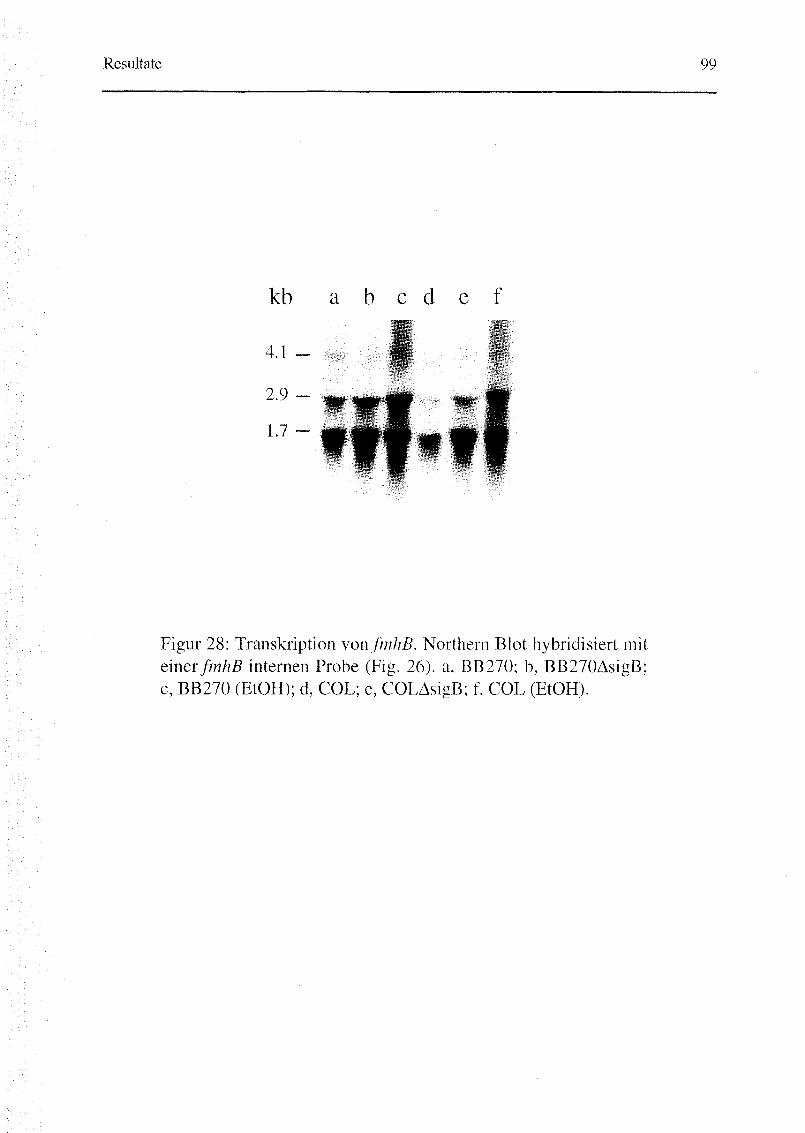
Resultate
kb a b c d e f
Figur 28: Transkription von fmhB. Northern Blot hybridisiert mit
einer fmhB internen Probe (Fig. 26). a, BB270; b, BB270AsigB;c, BB270 (EtOH); d, COL; e, COLAsigB; f. COL (EtOH).

Diskussion 100
4. Diskussion
4.1. Die Phosphoglukosamin-Mutase GlmM
GlmM katalysiert die Umlagerung des C6-Phosphats von Glukosamin-6-
Phosphat an das Cl zu Glukosamin-1-Phosphat (48, 116). Diese Umlagerung
wird als einer der ersten Schritte betrachtet, die schliesslich zur Bildung des
Nukleotidzuckers UDP-N-Acetylglukosamin führen, der in die Biosynthese des
Peptidoglykans involviert ist (Fig. 1). Die endgültige Zusammensetzung und
Struktur des Peptidoglykans in S. aureus glmM Mutanten wird dadurch kaum
beeinflusst. glmM Mutanten haben eine 5% niedrigere Quervernetzung des
Peptidoglykans verglichen mit der des Wildtyps (93). Neben den
Pentaglycinseitenketten kommen auch Seitenketten vor, die an Stelle von Glyl
ein Ala! enthalten. Die Alanin enthaltenden Seitenketten sind in glmM Mutanten
nicht mehr nachweisbar (93). Der Einfluss einer solch geringen Änderung auf
die Methicillin-Empfindlichkeit lässt sich möglicherweise dadurch erklären,
dass die Produktionsrate von Peptodiglykan-Vorläufern aufgrund der
glmM::Tn551 Insertion herabgesetzt wurde und dass die noch produzierte
Menge nicht ausreicht, um den von PBP2' ausgehenden Schutz gegen
Methicillin voll ausnützen zu können. Es war bereits bekannt, dass in E. coli
das Gen glmM für das Überleben des Bakteriums entscheidend ist (74). Da
aber das Peptidoglykan in S. aureus trotz der glmM::Tn551 Insertion kaum
beeinflusst und im Gegensatz zu E. coli eine Inaktivierung des glmM toleriert
wird, nimmt man an, dass in S. aureus für die Glukosamin-1-Phophat Synthese
möglicherweise ein alternativer Weg vorhanden ist, der bis jetzt noch nicht
identifiziert wurde. Diese Hypothese wird insofern von den Resultaten
unterstützt, als dass die phänotypischen Komplementierungsexperminte mit
Glukosamin, das dem Wachstumsmedium zugegeben wurde, ganz klar zeigen
konnten, dass die Methicillin-Empfindlichkeit in den glmM Mutanten deutlich
abnahm und beinahe das Resistenz-Niveau des Wildtyps erreichte. Das

Diskussion 101
zugegebene Glukosamin muss, um als Substrat für die Bildung von UDP-N-
Acetylglukosamin zur Verfügung zu stehen, zuerst am C6 phosphoryliert
werden und anschliessend muss die Phosphatgruppe an das Cl umgelagert
werden. Alternativ könnte Glukosamin direkt am Cl phosphoryliert werden.
Beim ersten Weg wäre eine Phosphoglukosamin-Mutase notwendig um eine
entsprechende Umlagerung ablaufen zu lassen. Da aber in den glmM Mutanten
GlmM nicht exprimiert wird, muss diese Aufgabe von einem anderen, noch
unbekannten Enzym oder mehreren Enzymen übernommen werden, die Teil
des postulierten alternativen Weges zur Bildung von UDP-N-Acetylglukosamin
wären (Fig. 16).
Da GlmM in die ersten Schritte der Zellwandbiosynthese involviert ist,
schien es zumindest denkbar, dass die glmM::Tn551 Insertion einen Einfluss
auf die Teichonsäuren Substitution des Peptidoglykans haben könnte (30). Es
zeigte sich jedoch, dass sowohl die Menge an Teichonsäuren, ihre
Zusammensetzung und N-Acetylglukosaminyl Substitution in Stationärphasen
Kulturen von PG108 und BB270 (46) als auch die „plating efficiencies" der
Phagen, die von N-Acetylglukosaminyl Modifikationen der Wandteichonsäure
abhängt (22), genau gleich waren (46).
4.1.1. Sequenz des Phosphoglukosamin-Mutase Operons
glmM wurde auf dem Smal I Fragment des NCTC8325 Chromosoms
lokalisiert. Es liegt am Ende eines dreizistronischen Operons das sich aus den
ORFs orfl-orß-glmM zusammensetzt. Dieses Operon resultiert in zwei
Transkripten, einem dreizistronischen bestehend aus allen drei ORFs und
einem monozistronischen glmM Transkript. Letzteres Transkript war in den
glmM Mutanten nicht mehr vorhanden; dafür erschien ein längeres aber
schwächeres Transkript von etwa 4.7 kb. Im Gegensatz zu den femAB oder
femC Mutanten, in welchen die Tn55i unterbrochenen Transkripte
entsprechend der Grösse vom Tn557 um 5.2 kb zunahmen, nahmen die
Transkripte in den glmM Mutanten weniger an Grösse zu. Der Unterschied

Diskussion 102
rührt wahrscheinlich von einr im Gegensatz zu den femAB undfemC Mutanten
unterschiedlichen Prozessierung der mRNA her.
Aus der Sequenzanalyse des glmM Operons ging hervor, dass das Tn557
im C-terminalen Bereich von glmM integriert hatte. Das aktive Zentrum
wurde dadurch nicht unterbrochen, da es sich im N-terminalen Bereich
befindet. Das möglicherweise daraus resultierende Transkript könnte vielleicht
für ein partielles GlmM kodieren, was allerdings durch die Western Blots
widerlegt werden konnte, da weder in PG108 noch in PG100 GlmM-Produkte
nachgewiesen werden konnte. In B. subtilis konnte eine dem glmM Operon
orfl-orß-glmM ähnliche Genorganisation ybbP-ybbR-ybbT gefunden werden
(Fig. 29). Entsprechend hoch waren die Identitäten der von den beiden
Operons kodierten Genen untereinander. YbbP, YbbR und YbbT zeigten zu
Orfl, Orf2 und GlmM eine Identität von 56, 28 und 67%. Da aber auch in B.
subtilis die Aufgabe der beiden Proteine YbbP und YbbR noch nicht klar ist,
konnten noch keine Rückschlüsse auf 5. aureus gezogen werden.
4.1.2. Suppressor Mutante PG100
Eine charakteristische Eigenschaft der mec-bedingten Resistenz der
MRSA Stämme ist deren heterogene Expression (29). Diese heterogene
Expression der Resistenz blieb auch nach der glmM;:Tn551 Insertion erhalten,
auch wenn die Grund-Resistenz abnahm. So konnte der Stamm PG100 isoliert
werden, der trotz der glmM Mutation sogar eine höhere MHK aufwies als
BB270. Die postulierte Suppressor Mutation hmrDl führte zur
Wiederherstellung oder sogar Erhöhung der Resistenz gegenüber den ß-
Laktamen Methicillin, Oxacillin und Imipenem. Um die Möglichkeit einer
möglichen Kotransduktion von hmrDl zusammen mit glmM::Tn551 zu
überprüfen, wurde mittels Phagen, gewachsen auf PG100, erneut BB270
transduziert. Der resultierende Stamm zeigte aber das gleiche Resistenz-Profil
wie PG108, was schliessen liess, dass die Mutation hmrDl, die die hohe

?
ybbP
ybbR
ybbT
glmS
?.9
orfl
orß
glmM
Ikb
Figur29:Vergle
ichderglmMRegionvon
S.aureusmitderRegion
inB.
subtilis,derenGene
fürProteinekodieren,
diehoheHomologie
zudenenvon
S.aureus
zeig
ten:
OrflzuYbbP
56%
Identitätund76%
Ähnl
ichk
eit;
Orf2zuYbbR28%
Identitätund53%
Ähnl
ichk
eit;
GlmM
zuYbbT65%
Identitätund80%
Ähnl
ichk
eit.9,
Terminator.

Diskussion 104
Resistenz gegenüber Methicillin verursacht mehr als 40 kb von glmM entfernt
sein muss.
Die offenbar durch die Tn551 Insertion in glmM verursachte Zunahme
der Empfindlichkeit gegenüber dem Glykopeptid Teicoplanin blieb in der
Suppressor Mutante unverändert. Das Glykopeptid Teicoplanin inhibiert
sowohl die Transglykosylase- als auch die Transpeptidase-Reaktion indem es
sich an das terminale D-Ala-D-Ala des Peptidoglykanvorläufers bindet. Diese
Eigenschaft des Teicoplanins wird zusätzlich durch seinen Membrananker
verstärkt (6). Daher wird vermutet, dass neben der fehlenden GlmM Aktivität
zusätzlich eine Membran Komponente in den glmM Mutanten betroffen sein
könnte. Das Hydropathie Profil von Orfl weist auf ein möglicherweise
Membran-assoziiertes Protein hin. Es ist also denkbar, dass die glmM;:Tn551
Insertion nicht nur die GlmM Aktivität beeinflusst.
Zudem zeigte PG100 im Vergleich zu PG108 und BB270 eine
schwächere spontane Autolyse. Es ist bereits bekannt, dass die Inaktivierung
von lytH, das für ein Autolysin kodiert, heterogene Methicillin-Resistenz in
hohe homogene Methicillin-Resistenz umwandelt (32). Bei PG100 führt die
postulierte hmrDl Mutation zu einer hohen aber nicht zu einer hohen
homogenen Methicillin-Resistenz. Trotzdem könnte bei der postulierten
Mutation hmrDl ein oder mehrere Gene betroffen sein, die die spontane
Autolyse beeinflussen.
Die Produktion eines 49 kDa grossen Membran Proteins scheint in
PG100 hochreguliert zu sein. Kompensatorische Mutationen, die zu hoher
Methicillin-Resistenz führen, wurden auch in femC Mutanten beschrieben,
wobei der Ursprung dieser Art von Mutation noch nicht bekannt ist (106). Wie
bei der postulierten hmrDl Mutation ist die kompensatorische Mutation, die in
femC Mutanten zu einer hohen homogenen Resistenz führt nicht mit der
/emC::Tn55/ Insertion kotransduzierbar (106). Es unterstützt damit die
Hinweise, wonach offenbar verschiedene Mechanismen zu dieser hohen
Methicillin-Resistenz beitragen können (106).

Diskussion 105
4.1.3. Transduktion von glmM::Tw551
Die Transduktionseffizienz ist in mec Determinante tragenden Stämmen erhöht
(9, 36). So erwies sich der MSSA BB255 als schlechterer Rezipient für
glmM::Tn551 als MRSA BB270. Da die seltenen stabilen BB255 glmM
Mutanten zudem einen zusätzlichen auffälligen Phänotyp aufwiesen, wurde
angenommen, dass eine zusätzliche Mutation notwendig ist, damit die
Inaktivierung von glmM durch Tn55/ in einem MSSA möglich ist. Ein
weiteres Indiz, dass die Transduktion von glmM::Tn551 in MSSA Stämme
durch zusätzliche chromosomale Mutationen beeinflussbar zu sein scheint, ist
die Transduktion von glmM::Tn551 eines in einen in vitro auf Methicillin-
Resistenz selektionierten BB255. Obwohl die Transduktionsfrequenz immer
noch niedriger war als die eines MRSA Stammes, hatten die Transduktanten
keine auffällige Veränderungen phänotypischer Merkmale (36).
Möglicherweise kompensierte in diesem Fall eine zusätzliche Mutation den
schädlichen Einfluss der glmM Mutation und ermöglichte so die Integration,
während in MRSA die mec Determinante diesen schützenden Effekt übernahm.
4.1.4. Komplementation der glmM Mutation
Die Komplementationen von glmM::Tn551 einerseits mit dem gesamten
glmM Operon, andererseits mit glmM allein haben gezeigt, dass offenbar
glmM ausreicht um die ß-Laktam-Resistenz und die MHK für Vancomycin
wiederherzustellen, nicht aber die MHK für Teicoplanin. Die MHK für
Teicoplanin blieb sowohl beim komplementierten PG108 als auch beim
komplementierten PG100 unverändert tief. Betrachtet man das Hydropathie
Profil von Orfl, so kann vermutet werden, dass es sich dabei um ein
Membran-assoziiertes Protein handeln könnte, da im N-terminalen Bereich ein
stark hydrophober Bereich ausgemacht werden konnte. Da Teicoplanin sich an
das terminale D-Ala-D-Ala des Peptidoglykanvorläufers bindet und auch mit

Diskussion 106
der Membran interagiert, könnte Orfl in die Teicoplanin-Resistenz involviert
sein.
Da GlmM von S. aureus eine E. coli glmM Mutante komplementieren
konnte und für GlmM Phosphoglukosamin-Mutase Aktivität nachgewiesen
werden konnte (48), stellte sich die Frage, ob durch Zugabe bestimmter
Zucker die glmM Mutation phänotypisch umgangen werden könnte. Es zeigte
sich, dass durch Zugabe von N-Acetylglukosamin oder Glukosamin ins
Medium die MHK für Methicillin wieder auf das Niveau von BB270 erhöht
wurde. Interessanterweise sind aber beide Substanzen nur Vorläufer und
müssen erst durch enzymatische Schritte in Glukosamin-1-Phosphat
umgewandelt werden. Da aber das für diesen Schritt notwendige Enzym GlmM
in den entsprechenden glmM Mutanten nicht nachgewiesen werden konnte,
wurde ein sekundärer Weg postuliert, der die Aufgabe von GlmM übernimmt
und kompensiert.
Die glmM::Tn551 Insertion reduziert nur die basale Resistenz von BB270, bei
E. coli hingegen ist eine Insertion in glmM letal. Bei S. aureus muss daher ein
sekundärer Weg existieren, der es dem Organismus ermöglicht trotz fehlender
Phosphoglukosamin-Mutase zu überleben. Dabei nimmt zwar die Resistenz ab,
aber es bleibt die Möglichkeit zur Ausbildung von hochresistenten
Subpopulationen bestehen. Die empfindlich gewordenen glmM Mutanten wie
PG108 verfügen offenbar über einen sekundären Weg um weiterhin
Zellwandvorläufer zu produzieren. Dieser sekundäre Weg scheint aber
weniger effizient zu sein, da die Resistenz in glmM Mutanten herabgesetzt ist.
Gibt man von aussen Glukosamin oder N-Acetylglukosamin zu, so nimmt die
Resistenz wieder zu und erreicht gleiche MHK-Werte wie in BB270. Offenbar
scheinen diese Substrate einen Einfluss auf den postulierten sekundären Weg zu
haben. Entweder bewirkt die grössere Menge an Substraten eine zusätzliche
Aktivierung der in den sekundären Weg involvierten Proteine oder aber deren
möglicherweise niedrigere Spezifität für diese Substrate wird durch die
erhöhte Menge überdeckt.

Diskussion 107
4.1.5. Proteine
Wie sich gezeigt hat, hatte die glmM::Tn551 Insertion keinen Einfluss
auf die PBP2' Expression da in den Mutanten gleich viel PBP2' exprimiert
wurde wie im Wildtyp. Auch bei der heterogen hochresistenten Suppressor
Mutante PG100 konnte keine Zunahme der PBP2' Produktion festgestellt
werden. Dass die postulierte hmrDl Mutation, die dem PG100 ein hohe
Methicillin-Resistenz verleiht, keinen Einfluss auf die PBP2' Produktion zu
haben scheint, korreliert mit der Beobachtung, dass zwischen der Menge an
produziertem PBP2' und der Höhe der Methicillin-Resistenz kein direkter
Zusammenhang besteht, und mit der Hypothese, dass noch andere Genprodukte
mit dem PBP2' kooperieren müssen. Das in PG100 in der Membranfraktion
stärker exprimierte Protein von 49 kDa Grösse könnte mit der erhöhten
Resistenz in Zusammenhang stehen, da gleichzeitig die spontane Autolyse in
PG100 reduziert war. Diese mögliche Korrelation muss aber weiter
experimentell bestätigt werden.
Die glmM::Tn551 Insertion hatte zur Folge, dass kein GlmM mehr
exprimiert wurde wie in den Western Blots gezeigt wurde. Im Wildtyp konnte
das GlmM nur in der zytoplasmatischen Fraktion nachgewiesen werden. In der
Membranprotein Fraktion konnte GlmM nicht nachgewiesen werden. GlmM
scheint also nur im Zytoplasma vorzukommen, was aufgrund seiner Aufgabe
innerhalb der Synthese der Zellwandvorläufer naheliegend ist.
4.2. Das fmtB-System
4.2.1. Transduktion und Wirkung
Die fmtB::Tn551 Insertion führte zu einer starken Abnahme der
Oxacillin-Resistenz und machte MRSA Stämme empfindlich gegen alle ß-
Laktame. Im homogen hochresistenten Stamm COL bewirkte eine fmtB

Diskussion 108
Mutation, dass die Mutante ein heterogenes Resistenz-Profil zeigte. Die
Transduktion von fmtB::Tn551 in den heterogen Methicillin-resistenten Stamm
BB270 führte einerseits zu TypI-Transduktanten, die Methiciliin-resistent
blieben, und andererseits zu TypII-Transduktanten, die wie COL-TS111
Methicillin-empfindlich wurden. Interessanterweise erschienen Transduktanten
vom Typl bereits nach zwei Tagen, die Transduktanten vom TypII hingegen
erst nach vier Tagen. Möglicherweise trugen die Transduktanten vom Typl
bereits vor der Transduktion eine chromosomale Mutation, die es erlaubte, die
fmtB::Tn551 Insertion zu kompensieren, so dass bereits nach zwei Tagen
Transduktanten wachsen konnten. Die empfindlich gewordenen Transduktanten
vom TypII brauchten im Gegensatz zu Typl länger um den Metabolismus auf
die Insertion in fintB einzustellen. Trotz dieser Diskrepanz zeigten beide Typen
mit Tn551 als Probe das gleiche Hybridisierungsmuster wie COL-TSlll, was
daraufhinweist, dass die fmtB: :Tn551 Insertion in beiden Typen stattgefunden
hatte. Die Transduktion von fmtB::Tn551 in empfindliche Stämme
funktionierte nicht ausser in RN450. Aus bisher noch unbekannten Gründen
hatte aber die Inaktivierung von fmtB in RN450 keinen Effekt auf dessen
Resistenz-Profil. Vermutlich zerstörte die fmtB::Tn551 Insertion die
Lebensfähigkeit der Zellen oder aber die wenigen Mutanten, die gefunden
werden konnten besassen ähnlich den resistenten fintB Mutanten vom Typl eine
sekundäre chromosomale Mutation, die der fmtB Mutation entgegenwirkt.
Da die Insertion von fmtB::Tn551, wie mittels Southern Blots gezeigt
werden konnte, in beiden Typen stattgefunden hatte, wurde folgende Hypothese
aufgestellt: Transduktanten vom Typl besassen eine /m?J5-kompensierende
chromosomale Mutation, während TypII Transduktanten, da sie erst nach vier
Tagen erschienen und Methicillin-empfindlich waren, diese postulierte
Mutation nicht enthielten, aber trotzdem einen Stoffwechsel induzieren müssen,
um mit der fintB Mutation überleben zu können

Diskussion 109
4.2.2. FmtB-Protein
fmtB kodiert für ein Protein noch unbekannter Funktion, das offenbar
Zellwand-assoziiert ist. Dies zeigt sich einerseits in seiner AS-Sequenz, die das
charakteristische Zellwand-sortierende Motif LPXTG beinhaltete, andererseits
in der Detektion von FmtB mittels Western Blot in der Membran-Protein-
Fraktion und dessen Fehlen sowohl im Kulturüberstand als auch in der
zytoplasmatischen Fraktion. Das Protein selbst besitzt vermutlich wichtige
Eigenschaften, die möglicherweise bei der Pathogenität eine Rolle spielen
könnten. So konnte in der Repeat Region von 75 AS Ähnlichkeit mit dem EF
Protein in Streptococcus suis (103) und mit dem Lmpl in Mycobacterium
hominis (65) gefunden werden. Proteine, die innerhalb ihrer AS-Sequenz
mehrfache Repeats enthalten, sind oft in die Bindung von Liganden involviert
(28, 43, 101, 114). Manchmal sind die AS-Sequenzen innerhalb solcher AS-
Repeatshoch konserviert oder zeigen Homologie Variationen (49, 115). Diese
Variationen wurden schon beim M-Protein von Streptococcus pyogenes (49),
in der Glukosyltransferase von Streptococcus downei und Staphylococcus
mutans und im Toxin A von Clostridium difficile (115) beschrieben. Bei FmtB
besitzen die 17 Repeats untereinander schwache Ähnlichkeit. Vom ersten bis
zum zwölften Repeat sind jeweils vier AS konserviert, während die letzten fünf
weniger gute Ähnlichkeiten untereinander zeigen. Ihre Bedeutung für FmtB ist
noch nicht klar (Komatsuzawa et al, Manuskript eingereicht).
4.2.3. Offene Fragen über den Effekt der fmtB Inaktivierung
Komatsuzawa konnte mit Western Blots zeigen, dass in der mit dem
Plasmid pHK4357 komplementierten fmtB Mutante COL-TSlll tatsächlich
FmtB exprimiert wurde (Komatsuzawa et al., Manuskript eingereicht). Er
konnte ebenfalls zeigen, dass es sich beim FmtB Protein um ein Zellwand-
assoziiertes Protein handelte, da die FmtB Bande nur im Ganz-Zell-Lysat und
im Zellwand-Extrakt, nicht aber im Kulturüberstand oder in der

Diskussion 110
zytoplasmatischen Fraktion nachgewiesen werden konnte. Wurde versucht die
fmtB Mutation entsprechend mit einem plasmidkodierten fmtB zu
komplementieren, so konnte die Resistenz nicht wiederhergestellt werden,
obwohl mittels Western Blot nachgewiesen wurde, dass im komplementierten
Stamm grosse Mengen an FmtB gebildet wurde. FmtB selbst ist daher nicht
direkt in die Methicillin-Resistenz involviert, sondern die Insertion hat
möglicherweise einen polaren Effekt auf andere, die Methicillin-Resistenz
beeinflussende Gene. Es wäre aber auch denkbar, dass FmtB zwar korrekt
gebildet wird und zur Zellwand gelangt, dass der Einbau aber nicht korrekt
erfolgt und somit die Aktivität ausbleibt. Andererseits konnte beobachtet
werden, dass der MRSA KSA8, der nur partielles FmtB ohne Zeliwanddomäne
und Zellwandanker herstellt, hochresistent ist. Dieses partielle FmtB konnte
nur im Kulturüberstand und in der Zytoplasma-Fraktion beobachtet werden.
Da aber eine fmtB;:Tn551 Insertion in KSA8 ebenfalls zur gleichen Reduktion
der Resistenz gegen Oxacillin und TritonX-100 führte wie bei COL-TSlll,
kann davon ausgegangen werden, dass FmtB selbst keinen direkten Einfluss auf
die Resistenz hat, was wiederum auf einen möglichen polaren Effekt Tn55i auf
andere Gene hinweist. In unmittelbarer Nähe zu fmtB weniger als 400
Nukleotide entfernt liegt proximal das Gen glmM. Obwohl Northern und
Western Blots gezeigt haben, dass die fmtB::Tn551 Insertion keinen Einfluss
auf die Transkription und Translation von GlmM hatte, konnte eine
Komplementation mit einem plasmidkodierten glmM die Oxacillin-Resistenz
quasi wieder herstellen, was bei einer fmtB Komplementation nicht der Fall
war. Nach der glmM kodierenden Sequenz folgt ein rho-unabhängiger
Terminator. Die mit einer glmM Probe detektierten Transkripte waren auch
alle zu kurz um zusätzlich fmtB zu beinhalten. Da aber nie ein fintB Transkript
nachgewiesen werden konnte, ist ebenfalls nicht auszuschliessen, dass es
möglicherweise doch ein paar wenige g/mM-/mr5-Transkripte geben könnte.
Neben der Möglichkeit die Oxacillin-Resistenz mittels glmM
Komplementation fast wieder herzustellen, konnte durch Zugabe von
Glukosamin oder N-Acetylglukosamin die Resistenz fast wieder auf Wildtyp

Diskussion 111
Niveau angehoben werden. Bei beiden Substanzen handelt es sich um Vorläufer
des UDP-N-Acetylglukosamin, einem Baustein des Peptidoglykans (Fig. 1).
GlmM wandelt Glukosamin-6-Phosphat in Glukosamin-1-Phosphat um, welches
dann sukzessive für die Zellwandbiosynthese weiterverarbeitet wird (Fig. 1).
Damit überhaupt Glukosamin-1-Phosphat entsteht ist also entweder GlmM
notwendig oder es existiert ein sekundärer Weg, der es ermöglicht, dass glmM
Mutanten das von aussen zugegebene Glukosamin verwerten können. Die
Zugabe eines plasmidkodierten glmM erhöhte in fmtB Mutanten die Oxacillin-
Resistenz fast auf Wildtyp-Niveau. Dies ist vermutlich auf die Überproduktion
von GlmM zurückzuführen, was zu einer effizienteren Umwandlung von
vorhandenen Glukosamin-6-Phosphat zu Glukosamin-1-Phosphat führte.
Andererseits lässt sich die fmtB Mutation gleich der glmM Mutation durch
Zugabe von Glukosamin oder N-Acetylglukosamin biochemisch
komplementieren. Im Gegensatz zu den glmM Mutanten besitzen die fmtB
Mutanten nach wie vor ein intaktes GlmM. Vermutlich wird das zugegebene
Glukosamin oder N-Acetylglukosamin in fmtB Mutanten nicht wie in den
glmM Mutanten über den postulierten sekundären Weg metabolisiert, sondern
die grössere Menge an Vorläufersubstanzen wird vom GlmM metabolisiert.
Die fmtB::Tn551 Insertion scheint offenbar keinen Einfluss auf GlmM zu
haben, da plasmidkodiertes glmM die Resistenz wieder heraufsetzen konnte. Es
ist anzunehmen, dass die fmtB: :Tn551 Insertion möglicherweise einen Einfluss
auf den postulierten sekundären Weg hat und die zugegebenen Substrate vom
chromosomalen glmM metabolisiert werden.
Die Aufgabe, die FmtB ausübt, ist noch nicht klar. Offenbar wirkt aber
FmtB in irgendeiner Form auf die Transkription oder Expression weiterer
chromosomaler Gene oder auf den Einbau von N-Acetylglukosamin-1-P in
UDP-NAG. Dabei kann ausgeschlossen werden, dass die fintB::Tn551 Insertion
einen Einfluss auf die GlmM Transkription oder Translation haben könnte, da
sowohl mit Northern als auch mit Western Blots glmM Transkripte und GlmM
nachgewiesen werden konnten. Möglicherweise beeinflusst FmtB die distal
liegenden Gene. Nach fintB konnte kein rho unabhängiger

Diskussion 112
Transkriptionsterminator gefunden werden und der nächste orf ist 1 kb
entfernt und wird divergent transkribiert. Dass aber nicht das vollständige
FmtB notwendig ist um Methicillin-Resistenz auszuprägen, zeigte sich in der
Wirkung der fmtB::Tn551 Insertion im MRSA KSA8, der für ein
unvollständiges FmtB kodiert.
Da ein plasmidkodiertes FmtB keinen Einfluss auf die Oxacillin-
Resistenz hat, wird angenommen, dass FmtB nur in eis wirken kann.
4.3. Das femX-System (fmhB-System)
Wenige Nukleotide distal zu fmhB konnte ein Gen identifiziert werden,
dessen Protein Ähnlichkeit zu einem Protein zeigt, das einen Einfluss auf die
Akriflavin-Resistenz hat. Es war bekannt, dass E. coli, die Mutationen in einem
Lokus, acrA genannt, tragen, hyperempfindlich gegen basische Farbstoffe,
Detergentien und Antibiotika sind (78). Die Akriflavin-Resistenz vermittelnden
Gene sind am besten in E. coli beschrieben (33). Die erhöhte Empfindlichkeit
ist jedoch nicht auf eine früher aufgestellte Hypothese zurückzuführen, die
besagte, dass eine acrAB Deletion die Permeabilität der äusseren Membran
erhöhe, sondern nach neuesten Ergebnissen auf das Fehlen des aktiven
Exportes der inhibierenden Substanzen. Nun besitzt ein von B. subtilis
kodiertes Protein, YerP, Ähnlichkeit mit dem AcrB, das ein Teil der Pumpe
repräsentiert. Zu diesem Protein YerP wiederum besitzt das unbekannte
Protein, das vom distalen ORF von fmhB kodiert wird, eine Identität von 42%
und eine Ähnlichkeit von 63%.

Literaturverzeichnis 113
1. Archer, G. L., D. M. Niemeyer, J. A. Thanassi, and M. J.
Pucci. 1994. Dissemination among staphylococci of DNA sequences associated
with methicillin resistance. Antimicrob Agents Chemother 38:447-454.
2. Arthur, M., P. E. Reynolds, F. Depardieu, S. Evers, S.
Dutkamalen, R. Quintiliani, and P. Courvalin. 1996. Mechanisms of
glycopeptide resistance in enterococci. J Infect 32:11-16.
3. Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G.
Seidman, J. A. Smith, and K. Struhl. 1987. Current protocols in
molecular biology, Greene Publishing Associates, New York.
4. Barber, M. 1947. Staphylococcal infections due to penicillin resistant
strains. British Medical Journal 2:863-865.
5. Barber, M. 1961. Methicillin-resistant staphylococci. Journal of
Clinical Pathology 14:385-393.
6. Beauregard, D. A., D. H. Williams, M. N. Gwynn, and D. J.
C. Knowles. 1995. Dimerization and membrane anchors in extracellular
targeting of vancomycin group antibiotics. Antimicrob Agents Chemother
39:781-785.
7. Beck, W. D., B. Berger-Bächi, and F. H. Kayser. 1986.
Additional DNA in methicillin-resistant Staphylococcus aureus and molecular
cloning of mec-specific DNA. J Bacteriol 165:373-378.
8. Beise, F., H. Labischinski, and P. Giesbrecht. 1988. Selective
inhibition of penicillin-binding proteins and its effects on growth and
architecture of Staphylococcus aureus. FEMS Microbiol Lett 55:195-202.

Literaturverzeichnis 114
9. Berger-Bächi, B. 1983. Increase in transduction efficiency of Tn557
mediated by the methicillin resistance marker. J Bacteriol 154:533-535.
10. Berger-Bächi, B. 1992. Methicillinresistenz bei Staphylococcus
aureus. Chemother J 2:58-64.
11. Berger-Bächi, B. 1996. Update on methicillin resistance mechanisms
in staphylococci. Chemother 42:19-26.
12. Berger-Bächi, B., L. Barberis-Maino, A. Strässle, and F. H.
Kayser. 1989. FemA, a host-mediated factor essential for methicillin
resistance in Staphylococcus aureus: molecular cloning and characterization.
Molec Gen Gen 219:263-269.
13. Berger-Bächi, B., and M. L. Köhler. 1983. A novel site on the
chromosome of Staphylococcus aureus influencing the level of methicillin
resistance: genetic mapping. FEMS Microbiol Lett 20:305-309.
14. Berger-Bächi, B., A. Strässle, J. E. Gustafson, and F. H.
Kayser. 1992. Mapping and characterization of multiple chromosomal factors
involved in methicillin resistance in Staphylococcus aureus. Antimicrob Agents
Chemother 36:1367-1373.
15. Berger-Bächi, B., and M. Tschierske. 1998. Role of Fem factors
in methicillin resistance. Drug Res Updates 1:325-335.
16. Brandis, H., W. Köhler, H. J. Eggers, and G. Pulverer. 1994.
Lehrbuch der Medizinischen Mikrobiologie, Gustav Fischer Verlag, Stuttgart,
Jena, New York.

Literaturverzeichnis 115
17. Brendel, V., and E. N. Trifonov. 1984. A computer algorithm for
testing potential prokaryotic terminators. Nucleic Acids Res 12:4411-27.
18. Brown, D. F. G., and P. E. Reynolds. 1980. Intrinsic resistance to
ß-lactam antibiotics in Staphylococcus aureus. FEBS Lett 122:275-278.
19. Chambers, H. F. 1988. Methicillin-resistant staphylococci. Clin
Microbiol Rev 1:173-186.
20. Chambers, H. F., and C. J. Hackbarth. 1987. Effect of NaCl and
nafcillin on penicillin-binding protein 2a and heterogeneous expression of
methicillin resistance in Staphylococcus aureus. Antimicrob Agents Chemother
31:1982-1988.
21. Cheung, A. L., and S. J. Projan. 1994. Cloning and sequencing of
sarA of Staphylococcus aureus, a gene required for the expression of agr. J
Bacteriol 176:4168-4172.
22. Coyette, J., and J. M. Ghuysen. 1968. Structure of the cell wall of
Staphylococcus aureus, strain Copenhagen. IX. Teichoic acid and phage
adsorption. Biochem 7:2385-2389.
23. Curtis, N. A. C, M. V. Hayes, A. W. Wyke, and J. B. Ward.
1980. A mutant of Staphylococcus aureus H lacking penicillin-binding protein
4 and transpeptidase activity. FEMS Microbiol Lett 9:263-266.
24. de Lencastre, H., B. L. M. de Jonge, P. R. Matthews, and A.
Tomasz. 1994. Molecular aspects of methicillin resistance in Staphylococcus
aureus. J Antimicrob Chemother 33:7-24.

Literaturverzeichnis 116
25. de Lencastre, H., and A. Tomasz. 1994. Reassessment of the
number of auxiliary genes essential for expression of high-level methicillin
resistance in Staphylococcus aureus. Antimicrob Agents Chemother 38:2590-
2598.
26. Duran, S. P., F. H. Kayser, and B. Berger-Bächi. 1996. Impact
of sar and agr on methicillin resistance in Staphylococcus aureus. FEMS
Microbiol Lett 141:255-260.
27. Ehlert, K., W. Schröder, and H. Labischinski. 1997. Specificities
of FemA and FemB for different glycine residues - FemB cannot substitute for
FemA in staphylococcal peptidoglycan pentaglycine side chain formation. J
Bacteriol 179:7573-7576.
28. Fahnestock, S. R., P. Alexander, J. Nagle, and D. Filpula.
1986. Gene for an immunoglobulin-binding protein from a group G
Streptococcus. J Bacteriol 167:870-880.
29. Figueiredo, A. M. S., E. Ha, B. N. Kreiswirth, H. de
Lencastre, G. J. Noel, L. Senterfit, and A. Tomasz. 1991. In vivo
stability of heterogeneous expression classes in clinical isolates of methicillin-
resistant staphylococci. J Infect Dis 164:883-887.
30. Fischer, W. 1990. Bacterial phosphoglycolipids and lipoteichoic acids.
M. Kates. Glycolipids, phosphoglycolipids, and sultoglycolipids. Plenum Press,
New York and London, pp 123-234.
31. Fleming, A. 1929. On the antibacterial action of cultures of a
pénicillium with special reference to their use in the isolation of B. influenzae.
Brit J Exp Pathol 10:226-236.

Literaturverzeichnis 117
32. Fujimura, T., and K. Murakami. 1997. Increase of methicillin
resistance in Staphylococcus aureus caused by deletion of a gene whose product
is homologous to lytic enzymes. J Bacteriol 179:6294-6301.
33. Funatsuki, K., R. Tanaka, S. Inagaki, H. Konno, K. Katoh,
and H. Nakamura. 1997. acrB mutation located at carboxyl-terminal region
of gyrase B subunit reduces DNA binding of DNA gyrase. J Biol Chem
272:13302-13308.
34. Gaisford, W. C, and P. E. Reynolds. 1989. Methicillin resistance
in Staphylococcus epidermidis. Relationship between the additional penicillin-
binding protein and an attachment transpeptidase. Eur J Biochem 185:211-
218.
35. Giesbrecht, P., J. Wecke, and B. Reinicke. 1976. On the
morphogenesis of the cell wall of Staphylococci. Int Rev Cytol 44:225-318.
36. Glanzmann, P. 1994. Genetische und physiologische Untersuchungen
anfemD Mutanten in Methicillin-resistenten Staphylococcus aureus.
Diplomarbeit. ETH Zürich.
37. Green, C. J., and B. S. Void. 1993. Staphylococcus aureus has
clustered tRNA genes. J Bacteriol 175:5091-5096.
38. Gustafson, J., A. Strässle, H. Hächler, F. H. Kayser, and B.
Berger-Bächi. 1994. The/<?mC locus of Staphylococcus aureus required for
methicillin resistance includes the glutamine synthetase Operon. J Bacteriol
176:1460-1467.
39. Haldenwang, W. G. 1995. The sigma factors of Bacillus subtilis.
Microbiol Rev 59:1-30.

Literaturverzeichnis 118
40. Hartman, B. J., and A. Tomasz. 1984. Low-affinity penicillin-
binding protein associated with beta-lactam resistance in Staphylococcus
aureus. J Bacteriol 158:513-516.
41. Henze, U., T. Sidow, J. Wecke, H. Labischinski, and B.
Berger-Bächi. 1993. Influence of femB on methicillin resistance and
peptidoglycan metabolism in Staphylococcus aureus, J Bacteriol 175:1612-
1620.
42. Hiramatsu, K., H. Hanaki, T. Ino, K. Yabuta, T. Oguri, and F.
C. Tenover. 1997. Methicillin-resistant Staphylococcus aureus clinical strain
with reduced vancomycin susceptibility. J Antimicrob Chemother 40:135-136.
43. Hollingshead, S. K., V. A. Fischetti, and J. R. Scott. 1986.
Complete nucleotide sequence of type 6 M protein of the group A
Streptococcus. Repetitive structure and membrane anchor. J Biol Chem
261:1677-1686.
44. Höltje, J. V. 1998. Growth of the stress-bearing and shape maintaining
murein sacculus of Escherichia coli. Microbiol Molec Biol Rev 62:181-203.
45. Inoue, H., H. Nojima, and H. Okayama. 1990. High efficiency
transformation of Escherichia coli with plasmids. Gene 96:23-8.
46. Jenni, R., and B. Berger-Bächi. 1998. Teichoic acid content in
different lineages of Staphylococcus aureus NCTC8325. Arch Microbiol
170:171-178.
47. Jevons, M. P. 1961. "Celbenin" resistant staphylococci. British Med J
1:124-125.

Literaturverzeichnis 119
48. Jolly, L., S. W. Wu, J. Van Heijenoort, H. De Lencastre, D.
Mengin Lecreulx, and A. Tomasz. 1997. The femR3\5 gene from
Staphylococcus aureus, the interruption of which results in reduced methicillin
resistance, encodes a phosphoglucosamine mutase. J Bacteriol 179:5321-5325.
49. Jones, K. F., S. K. Hollingshead, J. R. Scott, and V. A.
Fischetti. 1988. Spontaneous M6 protein size mutants of group A streptococci
display variation in antigenic and opsonogenic epitopes. Proc Natl Acad Sei U
S A 85:8271-8275.
50. Joris, B., K. Hardt, and J.-M. Ghuysen. 1994. Induction of ß-
lactamase and low affinity penicillin binding protein 2' synthesis in Gram-
positive bacteria, in. J.-M. Ghuysen and R. Hakenbeck. Bacterial cell wail.
Elsevier, Amsterdam. pp505-516.
51. Kayser, F. H., E. J. Benner, R. Troy, and P. D. Hoeprich.
1971. Mode of resistance against ß-lactam antibiotics in staphylococci. E. L.
Dulaney and A. I. Laskin. The problems of drug-resistant pathogenic bacteria.
The New York Academy of Science, New York, pp 106-117.
52. Kayser, F. H., K. A. Bienz, J. Eckert, and R. M. Zinkernagel.
1998. Medizinische Mikrobiologie, Georg Thieme Verlag, Stuttgart, New
York.
53. Kobayashi, N., K. Taniguchi, and S. Urasawa. 1998. Analysis of
diversity of mutations in the med gene and mecA promoter/operator region of
methicillin-resistant Staphylococcus aureus and Staphylococcus epidermidis.
Antimicrob Agents Chemother 42:717-720.

Literaturverzeichnis 120
54. Komatsuzawa, H., M. Sugai, K. Ohta, T. Fujiwara, S.
Nakashima, J. Suzuki, C. Y. Lee, and H. Suginaka. 1997. Cloning and
characterization of the fmt gene which affects the methicillin resistance level
and autolysis in the presence of Triton X-100 in methicillin-resistant
Staphylococcus aureus. Antimicrob Agents Chemother 41:2355-2361.
55. Komatsuzawa, H., M. Sugai, C. Shirai, J. Suzuki, K.
Hiramatsu, and H. Suginaka. 1995. Triton X-100 alters the resistance level
of methicillin-resistant Staphylococcus aureus to oxacillin. FEMS Microbiol
Lett 134:209-212.
56. Komatsuzawa, H., J. Suzuki, M. Sugai, Y. Miyake, and H.
Suginaka. 1994. The effect of Triton X-100 on the in-vitro susceptibility of
methicillin-resistant Staphylococcus aureus to oxacillin. J Antimicrob
Chemother 34:885-897.
57. Kopp, U., M. Roos, J. Wecke, and H. Labischinski. 1996.
Staphylococcal peptidoglycan interpeptide bridge biosynthesis: a novel
antistaphyiococcal target? Microb Drug Resist 2:29-41.
58. Kornblum, J., B. J. Hartman, R. P. Novick, and A. Tomasz.
1986. Conversion of a homogeneously methicillin-resistant strain of
Staphylococcus aureus to heterogeneous resistance by Tn55J -mediated
insertional inactivation. Eur J Clin Microbiol 5:714-718.
59. Kreiswirth, B. N., S. Löfdahl, M. J. Bentley, M. O'Reilly, P.
M. Schlievert, M. S. Bergdoll, and R. P. Novick. 1983. The toxic
shock syndrome exotoxin structural gene is not detectably transmitted by a
prophage. Nature 305:680-685.

Literaturverzeichnis 121
60. Kullik, I., and P. Giachino. 1997. The alternative sigma factor
sigma(b) in Staphylococcus aureus - regulation of the sigB Operon in response
to growth phase and heat shock. Arch Microbiol 167:151-159.
61. Kullik, I., P. Giachino, and T. Fuchs. 1998. Deletion of the
alternative sigma factor sigma(b) in Staphylococcus aureus reveals its function
as a global regulator of virulence genes. J Bacteriol 180:4814-4820.
62. Labischinski, H. 1992. Consequences of the interaction of beta-lactam
antibiotics with penicillin binding proteins from sensitive and resistant
Staphylococcus aureus strains. Med Microbiol Immunol 181:241-265.
63. Labischinski, H., H. Maidhof, M. Franz, D. Krüger, T. Sidow,
and P. Giesbrecht. 1988. Biochemical and biophysical investigations into the
cause of penicillin-induced lytic death of staphylococci: Checking predictions
of the murosome model, in. P. Actor, L. Daneo-Moore, M. L. Higgins, M. R.
J. Salton and G. D. Shockman. Antibiotic inhibition and function of bacterial
cell surface assembly and function. Society for Microbiology, Washington DC.
pp242-257.
64. Lacey, R. W., and J. Grinsted. 1973. Genetic analysis of
methicillin-resistant strains of Staphylococcus aureus; evidence for their
evolution from a single clone. J. Med. Microbiol. 6:511-526.
65. Ladefoged, S. A., S. Birkelund, S. Hauge, B. Brock, L. T.
Jensen, and G. Christiansen. 1995. A 135-kilodalton surface antigen of
Mycoplasma hominis PG21 contains multiple directly repeated sequences.
Infect Immun 63:212-223.

Literaturverzeichnis 122
66. Lee, C. Y., J. J. Schmidt, A. D. Johnson-Winegar, L. Spero,
and J. J. landolo. 1987. Sequence determination and comparison of the
exfoliative toxin A and toxin B genes from Staphylococcus aureus. J Bacteriol
169:3904-3909.
67. Ling, B. D., and B. Berger-Bächi. 1998. Increased overall
antibiotic susceptibility in Staphylococcus aureus femAB null mutants.
Antimicrob Agents Chemother 42:936-938.
68. Lorow, D., and J. Jessee. 1990. MAX efficiency DHlOßTM a new
host for cloning methylated DNA. Focus 12:19.
69. Lyon, B. R., and R. Skurray. 1987. Antimicrobial resistance of
Staphylococcus aureus: genetic basis. Microbiol Rev 51:88-134.
70. Maidhof, H., B. Reinicke, P. Bliimel, B. Berger-Bächi, and H.
Labischinski. 1991. femA, which encodes a factor essential for expression of
methicillin resistance, affects glycine content of peptidoglycan in methicillin-
resistant and methicillin-susceptible Staphylococcus aureus strains. J Bacteriol
173:3507-3513.
71. Maki, H., T. Yamaguchi, and K. Murakami. 1994. Cloning and
characterization of a gene affecting the methicillin resistance level and the
autolysis rate in Staphylococcus aureus. J Bacteriol 176:4993-5000.
72. Maniatis, T., E. F. Fritsch, and J. E. Sambrook. 1989.
Molecular cloning: a laboratory manual, Laboratory Manual, Cold Spring
Harbor Laboratories Press, New York.

Literaturverzeichnis
73. Matthews, P. R., and P. R. Stewart. 1984. Resistance heterogeneity
in methicillin-resistant Staphylococcus aureus. FEMS Microbiol Lett 22:161-
166.
74. Mengin-Lecreulx, D., and J. van Heijenoort. 1996.
Characterization of the essential gene glmM encoding phosphoglucosamine
mutase in Escherichia coli. J Biol Chem 271:32-39.
75. Münch, S. 1994. Nonradioactive Northern Blots in 1.5 days and with
substantially increased sensitivity through "alkaline blotting". Biochemica
3:30-32.
76. Murakami, K., K. Nomura, M. Doi, and T. Yoshida. 1987.
Production of low-affinity penicillin-binding protein by low- and high-
resistance groups of methicillin-resistant Staphylococcus aureus. Antimicrob
Agents Chemother 31:1307-1311.
77. Musser, J. M., and V. Kapur. 1992. Clonal analysis of methicillin-
resistant Staphylococcus aureus strains from intercontinental sources -
association of the mec gene with divergent phylogenetic lineages implies
dissemination by horizontal transfer and recombination. J Clin Microbiol
30:2058-2063.
78. Nakamura, H. 1968. Genetic determination of resistance to acriflavine,
phenethyl alcohol, and sodium dodecyl sulfate in Escherichia coli. J Bacteriol
96:987-996.
79. NCCLS, and N. C. f. C. L. Standards. 1997. Methods for dilution
antimicrobial susceptibility tests for bacteria that grow aerobically- forth
edition. Approved standard. NCCLS document M7-A4., National Committee
for Clinical Laboratory Standards document M7-A4., Villanova, Pa.

Literaturverzeichnis 124
80. Ng, E. Y., M. Trucksis, and D. C. Hooper. 1994. Quinolone
resistance mediated by norA: physiologic characterization and relationship to
flqB, a quinolone resistance locus on the Staphylococcus aureus chromosome.
Antimicrob Agents Chemother 38:1345-1355.
81. Niemeyer, D. M., M. J. Pucci, J. A. Thanassi, V. K. Sharma,
and G. L. Archer. 1996. Role of mecA transcriptional regulation in the
phenotypic expression of methicillin resistance in Staphylococcus aureus. J
Bacteriol 178:5464-5471.
82. Noble, W. C, Z. Virani, and R. G. A. Crée. 1992. Cotransfer of
vancomycin and other resistance genes from Enterococcusfaecalis NCTC
12201 to Staphylococcus aureus. FEMS Microbiol Lett 93:195-198.
83. North, E. A., and R. Christie. 1946. Acquired resistance of
staphylococci to the action of penicillin. Med J Australia 1:176-179.
84. Ornelas-Soares, A., H. de Lencastre, B. de Jonge, D. Gage,"
Y.-S. Chan, and A. Tomasz. 1993. The peptidoglycan composition of a
Staphylococcus aureus mutant selected for reduced methicillin resistance. J
Biol Chem 268:26268-26272.
85. Ornelas-Soares, A., H. de Lencastre, B. L. M. de Jonge, and
A. Tomasz. 1994. Reduced methicillin resistance in a new Staphylococcus
aureus transposon mutant that incorporates muramyl dipeptides into the cell
wall peptidoglycan. J Biol Chem 269:27246-27250.
86. Park, W., and M. Matsuhashi. 1984. Staphylococcus aureus and
Micrococcus luteus peptidoglycan transglycosylases that are not penicillin-
binding proteins. J. Bacteriol. 157:538-544.

Literaturverzeichnis 125
87. Patterson, T. A., and M. Dean. 1987. Preparation of high titer
lambda phage lysates. Nucleic Acids Res 15:6298.
88. Peng, H. L., R. P. Novick, B. Kreiswirth, J. Kornblum, and
P. Schlievert. 1988. Cloning, characterization, and sequencing of an
accessory gene regulator (agr) in Staphylococcus aureus. J Bacteriol
170:4365-4372.
89. Pinho, M. G., A. M. Ludovice, S. W. Wu, and H. De
Lencastre. 1997. Massive reduction in methicillin resistance by transposon
inactivation of the normal pbpl in a methicillin-resistant strain of
Staphylococcus aureus. Microb Drug Resist 3:409-413.
90. Plumbridge, J., and E. Vimr. 1999. Convergent pathways for
utilization of the amino sugars N-acetylglucosamine, N-acetylmannosamine,
and N-acetylneuraminic acid by Escherichia coli. J Bacteriol 181:47-54.
91. Projan, S. J., and R. P. Novick. 1997. The molecular basis of
pathogenicity, in:. K. B. Crossley and G. L. Archer. The staphylococci in
human disease. Churchill Livingstone, New York, pp 55-81.
92. Reynolds, P. E. 1988. The essential nature of staphylococcal
penicillin-binding proteins, in. P. Actor, L. Daneo-Moore, M. L. Higgins, M.
R. J. Salton and G. D. Shockman. Antibiotic inhibition of bacterial cell surface
assembly and function. ASM, Washington DC. pp 343-351.
93. Roos, M. H. H. 1995. Das Murein von Staphylococcus aureus.
Untersuchung des Einflusses von Antibiotikaresistenzdeterminanten auf die
Feinstruktur des Peptidoglykan von Staphylococcus aureus. Thesis, Freie
Universität Berlin.

Literaturverzeichnis 126
94. Ryffel, C, F. H. Kayser, and B. Berger-Bächi. 1992.
Correlation between regulation of mecA transcription and expression of
methicillin resistance in staphylococci. Antimicrob Agents Chemother 36:25-
31.
95. Saiki, R. K., D. H. Gelfand, S. Stoffel, S. J. Scharf, R.
Higuchi, G. T. Horn, K. B. Mullis, and H. A. Erlich. 1988. Primer-
directed enzymatic amplification of DNA with a thermostable DNA
polymerase. Science 239:487-491.
96. Schleifer, K. H., W. P. Hammes, and O. Kandier. 1976. Effect
of endogenous and exogenous factors on the primary structures of bacterial
peptidoglycan. Adv Microb Physiol 13:245-292.
97. Sharma, V. K., C. J. Hackbarth, T. M. Dickinson, and G. L.
Archer. 1998. Interaction of native and mutant Med repressors with
sequences that regulate mecA, the gene encoding penicillin-binding protein 2a
in methicillin-resistant staphylococci. J Bacteriol 180:2160-2166.
98. Shlaes, D. M., J. H. Shlaes, S. Vincent, L. Etter, P. D. Fey,
and R. V. Goering. 1993. Teicoplanin-resistant Staphylococcus aureus
expresses a novel membrane protein and increases expression of penicillin-
binding protein-2 complex. Antimicrob Agents Chemother 37:2432-2437.
99. Sidow, T., L. Johannsen, and H. Labischinski. 1990. Penicillin-
induced changes in the cell wall composition of Staphylococcus aureus before
the onset of bacteriolysis. Arch Microbiol 154:73-81.
100. Sieradzki, K., and A. Tomasz. 1996. A highly vanomycin-resistant
laboratory mutant of Staphylococcus aureus. FEMS Microbiol Lett 142:161-
166.

Literaturverzeichnis 127
101. Signas, C, G. Raucci, K. Jonsson, P. E. Lindgren, G. M.
Anantharamaiah, M. Hook, and M. Lindberg. 1989. Nucleotide
sequence of the gene for a fibronectin-binding protein from Staphylococcus
aureus: use of this peptide sequence in the synthesis of biologically active
peptides. Proc Natl Acad Sei U S A 86:699-703.
102. Skurray, R. A., D. A. Rouch, B. R. Lyon, M. T. Gillespie,
and J. M. Tennent. 1988. Multiresistant Staphylococcus aureus: genetics and
evolution of epidemic Australian strains. J Antimicrob Chemother 21:19-39.
103. Smith, H. E., F. H. Reek, U. Vecht, A. L. Gielkens, and M. A.
Smits. 1993. Repeats in an extracellular protein of weakly pathogenic strains
of Streptococcus suis type 2 are absent in pathogenic strains. Infect Immun
61:3318-3326.
104. Stewart, G. C, and D. E. Rosenblum. 1980. Genetic behavior of
the methicillin resistance determinant in Staphylococcus aureus. Journal of
Bacteriology 144:1200-1202.
105. Stranden, A. M., K. Ehlert, H. Labischinski, and B. Berger-
Bächi. 1997. Cell wall monoglycine cross-bridges and methicillin
hypersusceptibility in & femAB null mutant of methicillin-resistant
Staphylococcus aureus. J Bacteriol 179:9-16.
106. Stranden, A. M., M. Roos, and B. Berger-Bächi. 1996.
Glutamine synthetase and heteroresistance in methicillin-resistant
Staphylococcus aureus. Microb Drug Resist 2:201-207.

Literaturverzeichnis 128
107. Sugai, M., S. Yamada, S. Nakashima, H. Komatsuzawa, A.
Matsumoto, T. Oshida, and H. Suginaka. 1997. Localized perforation of
the cell wall by a major autolysin - atl gene products and the onset of
penicillin-induced lysis of Staphylococcus aureus. J Bacteriol 179:2958-2962.
108. Suzuki, E., K. Kuwaharaarai, R. J. F., and K. Hiramatsu.
1993. Distribution of mec regulator genes in methicillin-resistant
staphylococcus clinical strains. Antimicrob Agents Chemother 37:1219-1226.
109. Tomasz, A. 1990. Auxiliary genes assisting in the expression of
methicillin resistance in Staphylococcus aureus, in. R. P. Novick. Molecular
biology of the staphylococci. VCH Publishers Inc., New York, pp 565-584.
110. Tschierske, M., K. Ehlert, A. M. Stranden, and B. Berger-
Bächi. 1997. Lif, the lysostaphin immunity factor, complements FemB in
staphylococcal peptidoglycan interpeptide bridge formation. FEMS Microbiol
Lett 153:261-264.
111. Vasilieva, E., H. Labischinski, and B. Berger-Bächi. 1997.
Identification and mapping of new chromosomal sites affecting response to ß-
lactams in Staphylococcus aureus. Int J Antimicrob Agents 8:13-21.
112. Wada, A., and H. Watanabe. 1998. Penicillin-binding protein 1 of
Staphylococcus aureus is essential for growth. J Bacteriol 180:2759-2765.
113. Wecke, J., M. Lahav, I. Ginsburg, W. Kwa, and P.
Giesbrecht. 1986. Inhibition of wall autolysis of staphylococci by sodium
polyanetholsulfonate "liquoid". Arch Microbiol 144:110-115.
114. Welch, R. A. 1991. Pore-forming cytolysins of gram-negative
bacteria. Mol Microbiol 5:521-528.

Literaturverzeichnis
115. Wren, B. W. 1991. A family of clostridial and streptococcal ligand-
binding proteins with conserved C-terminal repeat sequences. Mol Microbiol
5:797-803.
116. Wu, S. W., H. de Lencastre, A. Sali, and A. Tomasz. 1996. A
phosphoglucomutase-like gene essential for the optimal expression of
methicillin resistance in Staphylococcus aureus: molecular cloning and DNA
sequencing. Microb Drag Resist 2:277-286.
117. Wu, S. W., H. de Lencastre, and A. Tomasz. 1996. Sigma-B, a
putative Operon encoding alternate sigma factor of Staphylococcus aureus RNA
polymerase: molecular cloning and DNA sequencing. J Bacteriol 178:6036-
6042.
118. Xiong, Z., and F. A. Kapral. 1992. Carotenoid pigment levels in
Staphylococcus aureus and sensitivity to oleic acid. J Med Microbiol 37:192-
194.

Anhang
6. Anhang
Lebenslauf
Name: Philipp Johann Glanzmann
Geburtsdatum: 6. Mai 1969
Zivilstand: ledig
Heimatort: Luzern
Staatsangehörigkeit: Schweiz
Ausbildung: Seit Juni 1995: Dissertation am Institut für
Medizinische Mikrobiologie der Universität Zürich
bei Frau Prof. B. Berger-Bächi.
Oktober 1990 - Mai 1995: Biologiestudium an der
ETH Zürich. Abschluss als diplomierter
Naturwissenschafter ETH in den Fächern: Gen- und
Enzymtechnologie/Immunologie, Genetik,
Immunbiologie, Mikrobiologie,
Molekularbiologie/Biophysik.
ETH-externe Diplomarbeit am Institut für
Medizinische Mikrobiologie der Universität Zürich.
August 1984 - Juni 1989: Kantonsschule Alpenquai,
Luzern; Literaturgymnasium, Matura Typus B.
August 1982 - Juli 1984: Sekundärschule Utenberg,
Luzern.
August 1976 - Juli 1982: Primarschule Maihof, Luzern.

Anhang 131
Publikationen
Ph. Glanzmann, J. Gustafson, H. Komatsuzawa, K. Ohta and B.
Berger-Bächi. 1999. glmM Operon and Methicillin-Resistant glmM
Suppressor Mutatnts in Staphylococcus aureus.
Antimicrob Agents Chemother 43: 240-245.
H. Komatsuzawa, K. Ohta, M. Sugai, T. Fujiwara, Ph. Glanzmann,
B. Berger-Bächi and H. Suginaka. Tn55 / Mediated Insertional
Inactivation of the fmtB Gene Encoding a Cell Wall Associated Protein
Abolishes Methicillin-Resistance in Staphylococcus aureus.
Manuskript eingereicht.
Konferenzen und Präsentationen:
20. - 21. Februar 1997 SGM St. Gallen; Poster:
femD is a Chromosomally Encoded Factor that affects
Methicillin-Resistance Expression in Staphylococcus
aureus.
25. - 28. Mai 1997 8th European Congress of Clinical Microbiology and
Infectious Diseases, Lausanne; Poster:
femD is a Chromosomally Encoded Factor that affects
Methicillin-Resistance Expression in Staphylococcus
aureus.
15. - 16. Oktober 1998 Scientific Meeting: Evolution of Bacterial Virulence
and Antibiotic Resistance, Lugano; Vortrag:
glmM Operon and Methicillin-Resistant glmM
Suppressor Mutants in Staphylococcus aureus.

Anhang 132
Danksagung
Frau Prof. Dr. Brigitte Berger-Bächi möchte ich besonders herzlich
dafür danken, dass ich in ihrer Arbeitsgruppe meine Dissertation durchführen
konnte und dass sie mich zu jeder Zeit und in jeder Hinsicht unterstützte.
Herrn Prof. Dr. Michael Teuber bin ich sehr dankbar dafür, dass er sich
bereit erklärt mir die Möglichkeit zu gegeben, meine Dissertation extern
auszuführen.
Ein besonderer Dank geht selbstverständlich an alle, die ich während den
Jahren am Institut kennen- und schätzen gelernt habe. Auch auf ihre Hilfe
konnte ich immer zurückgreifen.
Mein abschliessender Dank gebührt meiner Familie, die mir stets
hilfreich zur Seite stand, mich bei all meinen Vorhaben moralisch unterstützte
und mich darin bekräftigt hat nie aufzugeben.