the of vol. no. 34, molecular printed in u. s. a ... · brefeldin a causes disassembly of the golgi...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 263, No. 34, Issue of December 5, pp. 18545-18552,1988 Printed in U. S. A.
Brefeldin A Causes Disassembly of the Golgi Complex and Accumulation of Secretory Proteins in the Endoplasmic Reticulum*
(Received for publication, July 29, 1988)
Toshiyuki Fujiwara, Kimimitsu Oda, Sadaki YokotaS, Akira Takatsukig, and Yukio Ikeharan From the Department of Biochemistry, Fukuoka University School of Medicine, Jonan-ku, Fukuoka 814-01, the IDepartment of Anatomy, Yamanashi Medical School, Yamanashi 409-38, and the §Department of Agricultural Chemistry, Univeriity of Tokyo, Bunkyo-ku, Tokyo 113, Japan
The antiviral antibiotic brefeldin A (BFA) strongly inhibits the protein secretion in cultured rat hepato- cytes (Misumi, Y., Misumi, Y., Miki, K., Takatsuki, A., Tamura, G., and Ikehara, Y. (1986) J. Biol. Chern. 261, 11398-11403). We have further examined the inhibitory effect of the drug on intracellular transport of albumin by an immunocytochemical technique with peroxidase-conjugated Fab fragments of anti-rat al- bumin IgG. In hepatocytes treated with BFA (2.5 pg/ ml) for 1 h at 37 OC, no characteristic structures of the Golgi complex could be observed, and albumin was diffusely distributed in the endoplasmic reticulum (ER), nuclear envelope, and small vesicles around, in contrast to its condensed localization in the Golgi com- plex in the control cells. Such an unusual distribution of the secretory protein, however, was rearranged to the normal localization in the Golgi complex after 4 h even in the presence of the drug, possibly due to a metabolism of the drug to an inert form. Exposure of the cells to BFA with constant renewals (2.5 pg/ml at l -h intervals) or at a higher concentration (10 pg/ml) caused a prolonged accumulation of albumin in the ER, resulting in its dilation. These results indicate that BFA primarily blocks the protein transport from the ER to the Golgi complex, consistent with the biochem- ical data previously reported.
The general pathway for the transport of secretory proteins from their site of synthesis to the cell surface is now well established (Farquhar and Palade, 1981; Burgess and Kelly, 1987). Proteins destined to be secreted undergo three crucial sorting and translocation steps. The first segregation step into the lumen of the endoplasmic reticulum (ER)’ has been almost completely elucidated by discovery of the signal peptide/signal recognition particle/receptor system (Blobel and Dobberstein, 1975; Walter et al., 1984). The second step from the ER to the Golgi and the third step from the Golgi to the cell surface are also considered to be specific and highly regulated events. However, the complexity and specificity of these steps are still not totally understood. Recent studies indicate that the
* This work was supported in part by a Grant-in-Aid from the Ministry of Education, Science, and Culture of Japan. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduer- tisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
7 To whom correspondence and reprint requests should be ad- dressed Dept. of Biochemistry, Fukuoka University School of Medi- cine, Nanakuma, Jonan-ku, Fukuoka 814-01, Japan.
The abbreviations used are: ER, endoplasmic reticulum; BFA, brefeldin A; PBS, phosphate-buffered saline.
carboxyl-terminal sequence Lys-Asp-Glu-Leu appears essen- tial for retention of some proteins within the ER, suggesting a possible sorting signal for resident ER proteins discriminat- ing from other proteins including secretory proteins (Hender- shot et al., 1987; Munro and Pelham, 1987). Segregation of lysosomal proteins from secretory proteins occurs at the third step in the Golgi complex, where the mannose 6-phosphate- receptor system directs the targeting of lysosomal proteins to their destination in lysosomes (Kornfeld, 1987). Thus, the accumulating evidence suggests that transport of secretory proteins with no specific signals for retention in the ER and for targeting to lysosomes may occur by default in a rapid process of “bulk” flow (Burgess and Kelly, 1987; Rothman, 1987; Wieland et al., 1987).
Apart from such sorting mechanisms, the mechanisms un- derlying the transport system from the ER to the cell surface are also of importance to be elucidated. A number of electron microscopic observations suggest that transitional elements and small vesicles derived from them are thought to be in- volved in transport of proteins from the ER to the cis cisterna of the Golgi (Farquhar and Palade, 1981). A new type of coated vesicles that appear not to contain clathrin may play a role in protein transport within the Golgi stack (Orci et al., 1986). Coated vesicles containing clathrin in turn appear to mediate the transport to the cell surface via secretory vesicles (Griffiths et al., 1985; Orci et al., 1986). Individual steps in the transport machinery have been examined in detail by cell- free systems that reconstitute ER to Golgi transport (Hasel- beck and Scheckman, 1986; Balch et al., 1987) and cis to trans transport in the Golgi stack (Rothman et al., 1984), and also by in vivo systems which utilize different temperature de- pendencies at different transport steps (Matlin and Simons 1983; Saraste and Kuismanen, 1984; Fries and Lindstrom, 1986). Another invaluable approach in the analysis of these steps is the use of drugs affecting the secretory process at distinct sites in the cell, as elegantly demonstrated in the case of monensin (Tartakoff, 1983).
Brefeldin A (BFA), an antiviral antibiotic, is a unique fungal metabolite of a 13-membered marocyclic lactone ring (Harri et al., 1963). Recently, BFA was found to cause intra- cellular accumulation of high mannose type G protein and inhibition of its expression on the cell surface in vesicular stomatitis virus-infected baby hamster kidney cells (Takat- suki and Tamura, 1985). Our previous study (Misumi et al., 1986) subsequently demonstrated that the drug blocks protein secretion in cultured rat hepatocytes as strongly as monensin. However, in contrast to the evidence that monensin causes a delayed secretion of unprocessed forms of plasma proteins, BFA allowed the completely processed forms to be secreted after a prolonged accumulation of the unprocessed forms. In addition, the drug caused no inhibitory effect on the endocytic
18545

18546 Blockade of ER to Golgi Transport by BFA
pathway as judged by cellular uptake and degradation of '251- asialofetuin. In the present study, we have further character- ized a mode of the BFA action on the secretory pathway in cultured rat hepatocytes by immunocytochemical techniques.
EXPERIMENTAL PROCEDURES
Materials and Preparative Procedures-~-[~~S]Methionine (1200 Ci/mmol) was purchased from Du Pont-New England Nuclear. Ea- gle's minimum essential medium was from Nissui Seiyaku (Tokyo, Japan). BFA was isolated in crystalline form from fungi, FL-24, as described previously (Takatsuki and Tamura, 1985; Misumi et al., 1986). Stock solutions of BFA (2.5 or 10 mg/ml) were prepared in methanol and stored at -20 "C until use. Rat serum albumin was prepared and used for immunization in rabbits (Ikehara and Pitot, 1973; Yokota and Fahimi, 1981). Monospecific anti-rat albumin IgG was obtained from rabbit sera by affinity chromatography through an albumin-coupled Sepharose 6B column and digested with pepsin (Ikehara and Pitot, 1973). The resulting F(ab')z was reduced to yield Fab', which was conjugated with horseradish peroxidase (type VI, Sigma) by the method of Ishikawa et al. (1986). The conjugates were subjected to chromatography through an Ultrogel AcA 34 column (LKB, Broma, Sweden), and fractions of the first protein peak obtained were used as the peroxidase-conjugated Fab' for immuno- cytochemical staining.
Cell Culture and BFA Treatment-Hepatocytes were isolated from adult male Wistar rats, weighing 200-250 g, by the collagenase perfusing method (Seglen, 1976). The isolated cells were inoculated into 35-mm (for immunocytochemical experiments) or 60-mm (for labeling experiments) Falcon dishes which had been coated with collagen and cultured in Eagle's minimum essential medium, supple- mented with 5% newborn calf serum, 0.1 p~ insulin, 1 p~ dexameth- asone, and 60 pg/ml kanamycin (Oda et al., 1983). After a 6-h incubation at 37 "C, free cells and medium were removed from the dishes, and the fresh medium together with the above supplements was added to the cell monolayers. All the following experiments were performed using the cells after a 24-h culture. Unless otherwise indicated, the cells were incubated at 37 "C with BFA at the indicated concentrations in the Eagle's medium.
Conventional Electron Microscopy-Hepatocytes incubated in the absence or presence of BFA were washed twice with PBS and fixed at room temperature with freshly prepared 2.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.3). After 30 min, the cells were washed with 0.1 M phosphate buffer (pH 7.3) containing 0.25 M sucrose and postfixed with 1% osmium tetroxide in 0.1 M phosphate buffer (pH 7.4) for 1 h at 4 "C. Finally, the cells were dehydrated through a graded ethanol series, cleared in 2-hydroxypropyl methacrylate, and embedded in situ in Epon according to Brinkley et al. (1967). Thin sections were cut with a diamond knife on a Porter-Blum MT-2B ultramicrotome, collected on copper grids, and stained with uranyl acetate and lead citrate. Specimens were examined in a Hitachi HU- 12A electron microscope operating at 75 kV.
Irnmunoelectron Microscopy-Cells incubated under the indicated conditions were washed with PBS and fixed for 2 h at room temper- ature with the periodate-lysine-paraformaldehyde fixative containing 0.01 M periodate, 0.075 M lysine, and 2% paraformaldehyde in 37.5 mM phosphate buffer (pH 6.2) (McLean and Nakane, 1974). The cells were incubated for 30 min with 0.1 M glycine in PBS to quench aldehyde groups. After two washes with PBS, the cells were permea- bilized by treatment with 0.01% saponin in PBS for 20 min (Ohtsuki et al., 1978). Then the cells were incubated with peroxidase-conju- gated Fab' fragments of anti-albumin IgG (20 pg/ml) in PBS for 2 h at room temperature. The cells were washed with PBS and fixed with 2% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.4) for 20 min. After being washed with PBS, the cells were incubated for 15 min at room temperature with a substrate medium containing diaminoben- zidine (0.5 mg/ml) and 0.01% Hz02 in 0.05 M Tris-HC1 buffer (pH 7.0) (Graham and Karnovsky, 1966). The cells were washed with 0.1 M Tris-HC1 buffer (pH 7.0) and then osmicated with 1% osmium tetroxide in 0.1 M phosphate buffer (pH 7.4) at 4 "C for 1 h. The cells thus treated were processed for electron microscopy in the same way as described above. Thin sections were stained with lead citrate and examined in the Hitachi electron microscope.
Light Microscopy-For light microscopic observations, cells were cultured on 18-mm X 18-mm coverslips in 35-mm Falcon dishes. The cells on coverslips were directly fixed, permeabilized, and incubated with the immunoperoxidase followed by the enzyme reaction, as
described above. The coverslips attached with the cells were then mounted on glass slides with glycerol. Specimens were examined with a Nikon light microscope.
Labeling Experiments for Albumin Secretion-For pulse-chase ex- periments, hepatocytes (2 X lo6 cells/dish) were preincubated for 1 h at 37 "C in the absence or presence of BFA at the indicated concen- trations. The cells were pulse-labeled for 10 min with [35S]methionine (50 pCi/dish) and then chased. The drug, when indicated, was re- newed by medium exchanges at the pulse and chase. At the indicated times of chase, aliquots of the medium were taken and used for immunoprecipitation of albumin as described previously (Oda et al., 1983; Misumi et al., 1986).
Time-dependent effects of BFA on the secretion was examined as follows. Cells were preincubated at 37 "C in the presence of BFA (2.5 pglml). At the indicated times of preincubation, [35S]methionine (15 pCi/dish) was added into the medium followed by incubation of the cells without exchange of the medium. In one experiment, the medium which had been used for preincubation of cells for 4 h in the presence of BFA (2.5 pg/ml) was transferred to a fresh dish containing non- treated cells, which were incubated after the addition of [36S]methi- onine (15 pCi/dish) as above. Aliquots of the medium in each dish were taken at the indicated times and used for immunoprecipitation of albumin, followed by determination of the radioactivity. Relative secretion values are expressed as percentages of the radioactivity secreted in the presence of the drug compared with that secreted from the control culture at each time point.
RESULTS
Effect of BFA on Albumin Secretion-In a previous study (Misumi et al., 1986), we demonstrated that BFA has little effect on protein synthesis up to 1 pg/ml and at most 25% inhibition at 10 pg/ml in cultured rat hepatocytes. The drug, however, strongly inhibited the protein secretion in a dose- dependent manner. Fig. 1 shows the effect of BFA on the albumin secretion as a function of chase time after pulse label for 10 min with [35S]methionine. In the presence of BFA at 2.5 pg/ml, the secretion was completely blocked until 1 h; after that time, the newly synthesized albumin was rapidly secreted into the medium, reaching about 90% of the normally secreted level at 4 h of chase. At a higher concentration (10 pg/ml), the drug continuously blocked the secretion until 4 h of chase was examined. In the following experiments, we further examined ultrastructural changes of cells and intra- cellular localization of albumin under the two different con-
0 1 2 3 4 Chase Time ( h )
FIG. 1. Effect of BFA on albumin secretion. Rat hepatocytes (2 X IO6 cells/dish) were preincubated at 37 "C for 1 h in the absence (0) or presence of BFA at 2.5 pg/ml (0) and 10 pg/ml (A). The cells were pulse-labeled for 10 min with [35S]methionine (50 pCi/dish) and then chased. The drug, when indicated, was present in the medium throughout the pulse chase. At the indicated times of chase, aliquots of the medium were taken and used for immunoprecipitation of albumin. Values are expressed as percentages of the radioactivity secreted of the total radioactivity incorporated into the immunopre- cipitable albumin during a 10-min pulse.

Blockade of ER to Golgi Transport by BFA 18547
ditions with BFA, the reversible effect a t 2.5 pg/ml and the continuous effect a t 10 pg/ml.
Effect of BFA on Ultrastructure of Hepatocytes-Our pre- liminary observation (Misumi et al., 1986) showed contrasting findings depending upon drug doses used; no significant mor- phological changes were observed in the cells treated with 1 pg/ml for 2 h, whereas dilation of the ER was caused by treatment with 10 pg/ml for 4 h. In the present study, we have examined the drug effect on the ultrastructure in more detail. In contrast to the control cells where the Golgi complex was easily identified (Fig. 2 A ) , characteristic structures of the Golgi complex were hardly detectable in the cells upon incu- bation with BFA (2.5 pg/ml). Such a feature of the cells with no detectable Golgi structures was already observed as early as 15 min after exposure to the drug and maintained at least 1 h (Fig. 2B). A recognizable Golgi structure began to reappear around 2 h of the incubation, and the typical Golgi complex as that in the control cells was finally detectable at 3 to 4 h (Fig. 2C). During the incubation with BFA at 2.5 pg/ml, no significant changes were recognized in organelles other than the Golgi complex. When the cells were treated with a higher dose (10 pg/ml), no characteristic Golgi complex was observed in the cells throughout the incubation time up to 4 h (data not shown). In addition, a t later stages of incubation, the drug caused dilation of the ER (data not shown, see Fig. 5B), as previously reported (Misumi et al., 1986).
Effect of BFA on Intracellular Localization of Albumin-Fig. 3 shows light microscopic observations on albumin localiza- tion, for which cells cultured in monolayers were directly fixed, stained with immunoperoxidase, and observed without being sectioned. Under these conditions, peroxidase reaction products in the control cells were recognized as granules and tubular networks predominantly in perinuclear regions (Fig. 3A). The presence of the tubular network detected in the nonsectioned cells is in contrast to an immunostaining profile in sectioned cells, where albumin localization appears exclu- sively as granule-like (Yokota and Fahimi, 1981). Such a characteristic profile corresponding to the distribution of the Golgi complex, however, could not be observed in the cells treated with BFA (2.5 pg/ml) for 1 h (Fig. 3B). The reaction product was diffusely distributed throughout the cytoplasm of the treated cells, the photograph of which was slightly overexposed compared with that of control cells. When the cells were exposed to the drug for 2 h (Fig. 3C), the deposits were confined to granules, but did not yet form the typical tubular network. Essentially the same immunostaining profile with the network as that in the control cells was finally obtained in the cells which had been incubated for 4 h (Fig. 30).
The intracellular localization of albumin was further ex- amined in detail by immunoelectron microscopy (Fig. 4). In the control cells, the immunoperoxidase heavily labeled char- acteristic structures of the Golgi complex, a stack of cisternae and related vesicles (Fig. 4, A and 23). Our immunostaining procedure under the conditions used, however, did not reveal the presence of albumin in the ER (Fig. 4B), possibly due to a much lower concentration of the antigen therein. In con- trast, in the cells treated with BFA (2.5 pg/ml) for 1 h (Fig. 4, C and D), the ordinary structure of Golgi complex could not be identified anywhere, and the immunoreactive antigen was recognizable in the ER, nuclear envelope, and small vesicles scattered to the cytoplasm. A further incubation of the cells caused rearrangement in distribution of albumin. At 2 h of incubation, heavily stained structures, possibly imma- ture forms of Golgi complex, appeared, and concomitantly some areas of the ER showed a weak or negative reaction for
FIG. 2. Electron micrographs of rat hepatocytes. Cells were incubated a t 37 "C in the absence ( A ) or presence of BFA (2.5 pg/ml) for 1 h ( B ) and 4 h ( C ) . The cells were immediately fixed and processed for conventional electron microscopy. Thin sections were stained with uranyl acetate and lead citrate. Note that characteristic Golgi structures found in the control cells (A, inset) could not he observed in the cells treated with BFA for 1 h (R), but reappeared 4 h after the treatment ( C ) . GC, Golgi complex; M, mitochondria; N, nucleus. Bar, 0.5 pm.

18548 Blockade of ER to Golgi Transport by BFA
FIG. 3. Effect of BFA on intracellular localization of albumin observed by light microscopy. Cells were incubated a t 37 "C in the absence ( A ) or presence of BFA (2.5 pglml) for 1 h ( B ) , 2 h (C), and 4 h (D). The cells were immediately fixed, processed for immunostaining with anti-rat albumin Fab-horseradish peroxidase, and examined for light microscopy. The photograph of panel B was slightly over exposed compared with those of other panels. Bar, 10 pm.
the immunoperoxidase (Fig. 4E). In the cells treated for 4 h, Fig. 6, the most effective and almost complete blockade of the the immunoreaction was all negative in the ER and nuclear secretion was observed during the initial 1 h in the cells envelope, and the strongly stainable Golgi complex was finally identified (Fig. 40).
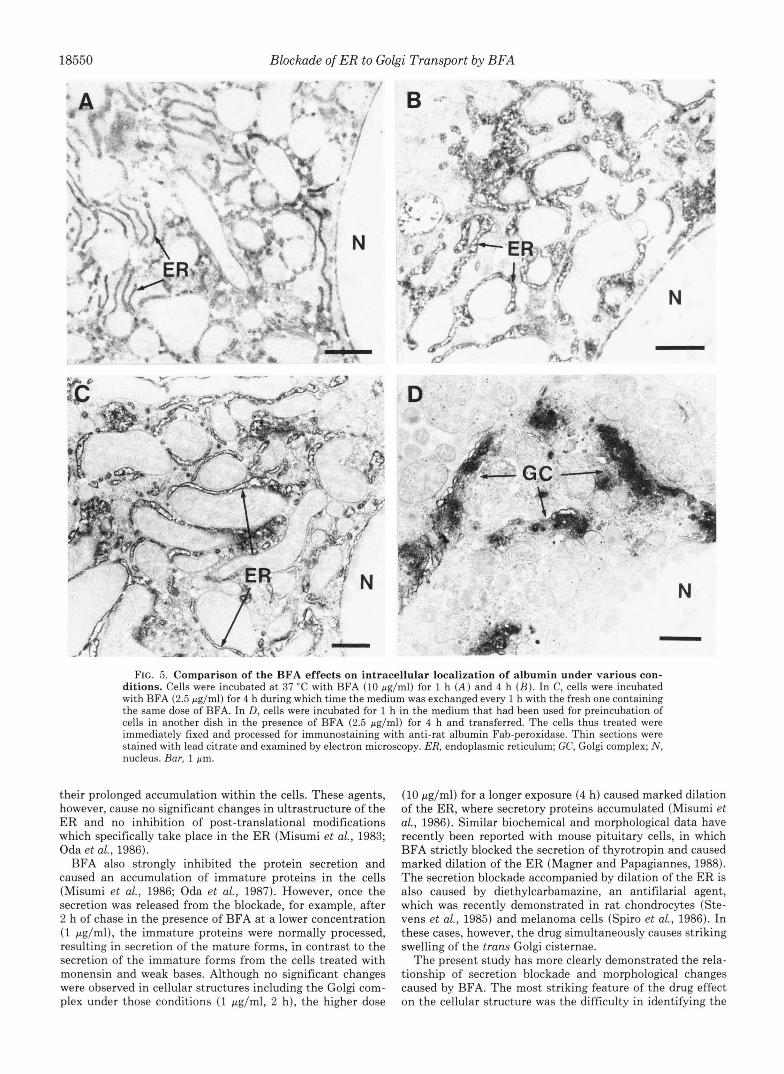
Exposure of cells to a higher dose (10 pg/ml) for 1 h caused a much clearer and intense immunostaining of the ER throughout the cells (Fig. 5A) compared with that in the cells treated with 2.5 pg/ml BFA for 1 h (Fig. 4C). A prolonged exposure (for 4 h) resulted in dilation of the ER, where albumin accumulated (Fig. 5B). No typical Golgi structures with or without the immunoreaction could be detected in the cells throughout the treatment period up to 4 h.
Time-dependent Loss of the BFA Effect-In contrast to the continuous effect by the high dose of BFA (Fig. 5, A and B ) , the effect caused by the lower dose (2.5 pg/ml) was found to be reversible (Fig. 4). However, when the medium containing 2.5 pg/ml BFA was renewed every 1 h, the drug caused disappearance of the Golgi complex and dilation of the ER even a t 4 h of the treatment (Fig. 5C), essentially the same profile as that in the cells treated with the higher dose (Fig. 5B). On the contrary, when the medium which had been used
exposed to the drug without preincubation, indicating that the drug exerts its effect immediately after its addition into the medium. When 1-h secretion values (open columns) were compared, the inhibitory effect of the drug was markedly decreased after 2 h of preincubation and only about 15% inhibition after 4 h. Such a time-dependent loss of the inhib- itory effect was disclosed earlier in 2-h secretion values (closed columns) as expected. The medium which had originally contained the same dose and been used for preincubation for 4 h in another dish (4(Re) in Fig. 6) had little effect on the albumin secretion, in agreement with the finding that the same medium caused no significant effect on the ultrastruc- ture of cells (Fig. 50). Taken together, these results suggest that the drug may be rapidly metabolized into an inert form during incubation with the cells.
DISCUSSION
The use of drugs affecting the secretory pathway at distinct sites in the cell may prove valuable for more detailed studies
for incubation of cells for 4 h in the presence of the drug (2.5 of specific steps in secretion and may lead to an understanding pg/ml) was re-used for incubation of fresh cells in another of the molecular basis of the mechanisms involved in intra- dish, no drug effect was observed in the cellular structure and cellular transport. One of the successful cases so far well albumin localization even a t 1 h of incubation (Fig. 50). characterized is the use of monensin, which is known to
The inhibitory effect of the drug on the albumin secretion perturb the function of the tram Golgi elements (Tartakoff was then examined as a function of preincubation time of and Vassalli, 1977; Tartakoff, 1983). The ionophore and other cells after the addition of the drug (2.5 pglml). As shown in acidotropic amines disrupt the maintenance of an acidic in-

Blockade of ER to Golgi Transport by BFA 18549
FIG. 4. Immunoelectron micrographs demonstrating changes in intracellular localization of albumin caused by BFA treatment. Cells were incubated in the absence ( A and B ) or presence of BFA (2 .5 pg/ml) for 1 h ( C and D), 2 h ( E ) , and 4 h ( F ) . The cells were immediately fixed and processed for immunostaining with anti- rat albumin Fab-peroxidase. Thin sections were stained with lead citrate. Panel D shows an enlargement of the squared area in panel C. ER, endoplasmic reticulum; GC, Golgi complex; M, mitochondria; NE, nuclear envelope; PM, plasma membrane. Bar, 1 pm.
ternal milieu which is functionally important in the trans (Oda and Ikehara, 1982; Oda et al., 1983) and weakly basic Golgi (Tartakoff, 1983; Anderson and Pathak, 1985; Schwartz amines (Oda and Ikehara, 1985; Oda et al., 1986) strongly et al., 1985), resulting in marked swelling and vacuolization inhibit the proteolytic and oligosaccharide processing of se- of the related elements and in blockade of some protein cretory proteins in cultured rat hepatocytes, and unprocessed modifications specifically occurring therein. In fact, monensin immature forms are finally released into the medium after
18550 Blockade of ER to Golgi Transport by BFA -
f
FIG. 5. Comparison of the BFA effects on intracellular localization of albumin under various con- ditions. Cells were incubated at 37 "C with BFA (10 pg/ml) for 1 h ( A ) and 4 h ( B ) . In C, cells were incubated with BFA (2.5 pg/ml) for 4 h during which time the medium was exchanged every 1 h with the fresh one containing the same dose of BFA. In D, cells were incubated for 1 h in the medium that had been used for preincubation of cells in another dish in the presence of BFA (2.5 pg/ml) for 4 h and transferred. The cells thus treated were immediately fixed and processed for immunostaining with anti-rat albumin Fab-peroxidase. Thin sections were stained with lead citrate and examined by electron microscopy. ER, endoplasmic reticulum; GC, Golgi complex; N , nucleus. Bur, 1 pm.
their prolonged accumulation within the cells. These agents, however, cause no significant changes in ultrastructure of the ER and no inhibition of post-translational modifications which specifically take place in the ER (Misumi et al., 1983; Oda et al., 1986).
BFA also strongly inhibited the protein secretion and caused an accumulation of immature proteins in the cells (Misumi et al., 1986; Oda et al., 1987). However, once the secretion was released from the blockade, for example, after 2 h of chase in the presence of BFA at a lower concentration (1 pg/ml), the immature proteins were normally processed, resulting in secretion of the mature forms, in contrast to the secretion of the immature forms from the cells treated with monensin and weak bases. Although no significant changes were observed in cellular structures including the Golgi com- plex under those conditions (1 pglml, 2 h), the higher dose
(10 pg/ml) for a longer exposure (4 h) caused marked dilation of the ER, where secretory proteins accumulated (Misumi et al., 1986). Similar biochemical and morphological data have recently been reported with mouse pituitary cells, in which BFA strictly blocked the secretion of thyrotropin and caused marked dilation of the ER (Magner and Papagiannes, 1988). The secretion blockade accompanied by dilation of the ER is also caused by diethylcarbamazine, an antifilarial agent, which was recently demonstrated in rat chondrocytes (Ste- vens et al., 1985) and melanoma cells (Spiro et al., 1986). In these cases, however, the drug simultaneously causes striking swelling of the tram Golgi cisternae.
The present study has more clearly demonstrated the rela- tionship of secretion blockade and morphological changes caused by BFA. The most striking feature of the drug effect on the cellular structure was the difficulty in identifying the

Blockade of ER to Golgi Transport by BFA 18551
0 1 ~ 2 4 4
Preincubation Time ( h )
FIG. 6. Time-dependent loss of the BFA effect on albumin secretion. Cells were preincubated at 37 “C in the absence or pres- ence of BFA (2.5 wg/ml). At the indicated times (0, 1, 2, and 4 h), [35S]methionine (15 pCi/dish) was directly added into the medium followed by incubation of the cells without medium exchange. In one experiment (indicated by 4(Re)), the medium which had been used for preincubation of cells with the same dose of BFA for 4 h was reused for another dish containing nontreated cells, which then received [35S]methionine and were incubated as above. Aliquots of the medium were taken from each dish at 1 h (open column) and 2 h (closed column) after the addition of [35S]methionine and used for immunoprecipitation of albumin, followed by determination of the radioactivity. Values are expressed as percentages of the radioactivity secreted from the treated cells as compared with that secreted from the control culture at each time point.
Golgi complex. The immunocytochemical examination of al- bumin localization easily identified the characteristic Golgi structure in the normal cells: granule-tubular network under light microscopy and stacks of cisternae and related vesicles under electron microscopy. Such characteristic profiles of the Golgi complex could not be observed in the cells treated with BFA. Its disappearance was found to be a very rapid event recognizable as early as 15 min after the exposure to the drug. Concomitantly, albumin began to accumulate in the ER, nuclear envelope, and small vesicles scattered in the cyto- plasm, in all of which the antigen was undetectable in the normal cells under the conditions used here. At a low concen- tration (2.5 pg/ml), such an unusual distribution of albumin was recovered to the normal localization in the Golgi complex at later stages of incubation, accounting for the delayed se- cretion of the newly synthesized albumin and the previous morphological observation obtained a t 1 pg/ml for 2 h (Mis- umi et al., 1986). At the higher dose (10 pg/ml) causing the continuous blockade of secretion, however, the altered distri- bution of albumin was maintained throughout the observation period up to 4 h, resulting in dilation of the ER. Apparent lack of the effect at later stages of incubation with the lower dose may be attributed to a metabolism of the drug to an inert form within the cells. Constant renewals of the drug with the same dose caused the same effects on the secretion and cellular structures as those obtained with the high dose (10 pg/ml), whereas the drug contained in the preincubated me- dium no longer exerted the effect. Such a drug-metabolizing activity may be variable depending upon one cell type to another; in baby hamster kidney cells, the inhibitory effect was caused by 0.5 pg/ml BFA and maintained as long as 4 h (Takatsuki and Tamura, 1985).
Taken together, these results suggest that BFA rapidly causes disassembly of the stack of Golgi cisternae and blocks the formation of vesicles involved in protein transport from the ER to the cis Golgi cisterna. The vesicular transport
system between compartments primarily consists of two major events, vesicle formation (budding) from one compartment and membrane fusion with another compartment. Recent detailed studies with cell-free systems have revealed various requirements for the vesicular transport from the ER to the cis Golgi cisterna (Balch et al., 1986, 1987) and from the cis to trans Golgi cisternae (Rothman et al., 1984; Balch et al., 1984; Wattenberg et al., 1986). ATP, cytosolic components, and N-ethylmaleimide-sensitive component(s) are found to be essential factors, especially for the first vesicle formation step in both the systems (ER to Golgi and cis to trans Golgi), although it remains to be determined whether there is a fundamental difference in this step between the two systems. Of these factors, at least the intracellular ATP level was not influenced by BFA under various conditions examined (1 to 10 pg/ml for 1 to 3 h) (Misumi et al., 1986). This may also be supported by the fact that the drug has no significant effect on the protein synthesis as well as on the protein degradation coupled with the receptor-mediated endocytosis. Although involvement of BFA in dysfunction of the other factors men- tioned above remains to be determined, the lipophilic nature of the drug (Harri et al., 1963) suggests its direct interaction with a membrane component playing a crucial role in the vesicular transport of the secretory pathway.
Little is known about the mechanisms underlying the or- ganization and maintenance of the Golgi stack with ordered arrangement from the cis to trans cisternae. Thus, there is no reasonable explanations available a t present for the mecha- nism accounting for the rapid disassembly or disappearance of the characteristic Golgi structures caused by BFA. It is, however, noteworthy that marked alterations of the protein transport and Golgi structures also occur in cells during mitosis. In mitotic cells after infection with vesicular stoma- titis viruses, the intracellular transport of G protein was found to be arrested, resulting in its accumulation in the ER (Feath- erstone et al., 1985), where the typical Golgi stack disappeared and instead there were observed multivesicular structures termed “Golgi clusters” carrying the marker enzymes galac- tosyltransferase and thiamine pyrophosphatase (Lucocq et al., 1987). Although these findings are very similar with those obtained with BFA, the lack of effect of the drug on endocy- tosis (Misumi et al., 1986) is in contrast to the apparent block of endocytosis observed during mitosis. A further exploration on the mechanism of the drug action should provide insight into the molecular basis of organization of the secretory pathway.
Acknowledgments-We thank Drs. Y. Misumi and N. Takami for discussion and help in some part of the experiments, and Drs. T. Katsuki and M. Yamaguchi (Dept. of Chemistry, Kyushu University School of Science) for providing synthetic compounds related to BFA.
REFERENCES
Anderson, R. G. W., and Pathak, R. K. (1985) Cell 40, 635-643 Balch, W. E., Blick, B. S., and Rothman, J. E. (1984) Cell 39, 525-
Balch, W. E., Elliott, M. M., and Keller, D. S. (1986) J. Biol. Chem.
Balch, W. E., Wagner, K. R., and Keller, D. S. (1987) J. Cell Biol.
Blobel, G., and Dobberstein, B. (1975) J. Cell Biol. 67, 835-851 Brinkley, B. R., Murphy, P., and Richardson, L. C. (1967) J . Cell
Burgess, T. L., and Kelly, R. B. (1987) Annu. Reu. Cell Biol. 3, 243-
Farquhar, M. G., and Palade, G. E. (1981) J. Cell Biol. 91, 77s-103s Featherstone, C., Griffiths, G., and Warren, G. (1985) J. Cell Biol.
Fries, E., and Lindstrom, I. (1986) Biochem. J. 237, 33-39
536
261, 14681-14689
104,749-760
Biol. 35, 279-283
293
101, 2036-2046

18552 Blockade of ER to Golgi Transport by BFA Graham, R. C., and Karnovsky, M. J. (1966) J. Histochem. Cytochem.
Griffiths, G., Pfeiffer, S., Simons, K., and Matlin, K. (1985) J. Cell
Harri, E., Loeffler, W., Sing, H. P., Stahelin, H., and Tamm, C. (1963)
Haselbeck, A., and Scheckman, R. (1986) Proc. Natl. Acad. Sci.
Hendershot, L., Bole, D., Kohler, G., and Kearney, J. F. (1987) J.
Ikehara, Y., and Pitot, H. C. (1973) J. Cell Bwl. 5 9 , 28-44 Ishikawa, E., Hashida, S., Kohno, T., Kotani, T., and Ohtaki, S.
(1986) in Monoclonal Antibodies: Hybridoma Techniques (Schook, L. B., ed) pp. 113-137, Marcel Dekker, Inc., New York
14,291-302
Biol. 101, 949-964
Helv. Chim. Acta 46, 1235-1243
U. S. A. 8 3 , 2017-2021
Cell Biol. 104 , 761-767
Kornfeld, S. (1987) FASEB J. 1, 462-468 Lucocq, J . M., Pryde, J. G., Berger, E. G., and Warren, G. (1987) J.
Magner, J . A., and Papagiannes, E. (1988) Endocrinology 122 , 912-
Matlin, K., and Simons, K. (1983) Cell 34 , 233-243 McLean, I. W., and Nakane, P. K. (1974) J. Histochem. Cytochem.
Misumi, Y., Tanaka, Y., and Ikehara, Y. (1983) Biochem. Biophys. Res. Commun. 114,729-736
Misumi, Y., Misumi, Y., Miki, K., Takatsuki, A., Tamura, G., and Ikehara, Y. (1986) J. Biol. Chem. 261 , 11398-11403
Munro, S., and Pelham, H. R. B. (1987) Cell 48,899-907 Oda, K., and Ikehara, Y. (1982) Biochem. Biophys. Res. Commun.
Oda, K., and Ikehara, Y. (1985) Eur. J. Biochem. 152,605-609 Oda, K., Misumi, Y., and Ikehara, Y. (1983) Eur. J. Biochem. 135,
Cell Biol. 104, 865-874
920
22,1077-1083
105,766-772
209-216
Oda, K., Koriyama, Y., Yamada, E., and Ikehara, Y. (1986) Biochem.
Oda, K., Hirose, S., Takami, N., Misumi, Y., Takatsuki, A., and
Ohtsuki, I., Manzi, R. M., Palade, G. E., and Jamieson, J . D. (1978)
Orci, L., Glick, B. S., and Rothman, J. E. (1986) Cell 4 6 , 171-184 Rothman, J. E. (1987) Cell 50,521-522 Rothman, J. E., Miller, R. L., and Urbani, L. J. (1984) J. Cell Biol.
Saraste, J., and Kuismanen, E. (1984) Cell 38,535-549 Schwartz, A. L., Strous, G. J. A. M., Slot, J. W., and Geuze, H. J.
Seglen, P. (1976) in Methods in Cell Biology (Prescott, D. M., ed) Vol. 13, pp. 29-83, Academic Press, Orlando, FL
Spiro, R. C., Parsons, W. G., Perry, S. K., Caulfield, J. P., Hein, A., Reisfeld, R. A., Harper, J. R., Austen, K. F., and Stevens, R. L.
Stevens, R. L., Parsons, W. G., Austen, K. F., Hein, A., and Caulfield,
Takatsuki, A., and Tamura, G. (1985) Agric. Biol. Chem. 4 9 , 899-
Tartakoff, A. M. (1983) Cell 32, 1026-1028 Tartakoff, A. M., and Vassalli, P. (1977) J. Exp. Med. 146 , 1332-
Walter, P., Gilmore, R., and Blobel, G. (1984) Cell 38 , 5-8 Wattenberg, B. W., Balch, W. E., and Rothman, J. E. (1986) J. Biol.
Wieland, F. T., Gleason, M. L., Serafini, T. A., and Rothman, J . E.
Yokota, S., and Fahimi, H. D. (1981) Proc. Natl. Acad. Sci. U. S. A.
J. 240 , 739-745
Ikehara, Y. (1987) FEBS Lett. 2 1 4 , 135-138
Biol. Cell 3 1 , 119-126
99,260-271
(1985) EMBO J. 4,899-904
(1986) J. BWl. Chem. 261,5121-5129
J. P. (1985) J. Biol. Chem. 260,5777-5786
902
1345
Chem. 261,2202-2207
(1987) Cell 50,289-300
78,4970-4974